水竹菱斑病發病特點、病原生物學特性及室內藥劑篩選
2021-05-27 08:43:38齊若涵楊春琳李琳劉暢許秀蘭劉裕峰曾倩劉利娟劉應高
東北林業大學學報 2021年5期
關鍵詞:生長
齊若涵 楊春琳 李琳 劉暢 許秀蘭 劉裕峰 曾倩 劉利娟 劉應高
(長江上游森林資源保育與生態安全國家林業和草原局重點實驗室(四川農業大學),成都,611130))
水竹(Phyllostachysheteroclada),又名黑水竹、奉化水竹、黎子竹等,隸屬禾本科(Poaceae)、竹亞科(Bambusoideae)、簕竹族(Bambuseae)、剛竹屬(Phyllostachys),屬于中小型散生竹種,主要分布于四川、甘肅、陜西、河南,長江流域及其以南地區。在四川省的雅安、宜賓、樂山等地存在大面積純林,全省分布面積約2萬hm2,在水土保持和生物多樣性保育[1-2]、竹筍制品和生物制劑[3-4]、竹工藝品和竹漿造紙[5-6]、庭院裝飾和園林布景[7]等方面發揮著重要作用。目前,關于水竹病害的研究報道較少,且早期文獻僅有簡單的記述,如枯萎病(Fusariumspp.)、稈枯病(Ellisembiapseudoseptata)、稈銹病(Stereostratumcorticioides)、竹團子病(Shiraiabambusicola)、竹肉球病(Engleromycesgoetzei)、叢枝病(Aciculosporiumtake)、黑粉病(Bambusiomycesshiraianus)和疹斑病(Phyllachoraspp.)等[8]。
2015年7月,課題組在雅安市雨城區開展竹類病害調查時首次發現該病害。水竹菱斑病主要危害當年生及以上成竹的竹稈、枝條和竹鞭(裸露),感病部位可形成典型的菱形病斑,在林間最終形成枯株型、枝枯型和梢枯型病竹。形態學特征描述結合多基因系統演化分析(LSU、SSU、ITS和TEF1-α),確定水竹菱斑病病原菌(Neostagonosporellasichuanensis)為一新屬新種[9]。2016—2019年野外調查發現,該病害在雅安市雨城區、天全縣和蘆山縣,以及廣安華鎣市均有不同程度的發生,部分地區危害嚴重,發病率達100%。本研究針對水竹菱斑病的發生展開定點定期調查,對病原菌進行生物學特性觀測和室內藥劑毒力試驗,旨在掌握病害的發生特點和病原的培養特性,為藥劑防治提供科學依據。
1 材料與方法
1.1 野外調查
2015年7月至2016年4月,在雅安市雨城區、天全縣和蘆山縣32個樣地開展野外調查,依據“病害發生普遍,水竹純林地,人為干擾小,便于設點調查”的4項原則,從以上32個樣地中選取3個典型樣地進行定點定期調查(見表1)。采用踏查記錄法(S曲線),通過目測(近地面2 m以下清點病斑數,2 m以上估測病斑數),對樣地各竹株(成竹)發病情況進行統計和記錄,依此對各樣地的病害發生情況進行評估。定點調查時間段為2016年4月至2017年4月,每2月進行1次調查和記錄。依據水竹菱斑病病情指數分級標準(見表2),計算病情指數,同時觀察新老水竹生長狀況和出筍等情況。此外,定點觀測病原菌的生態習性,主要包括病菌生活世代、越冬形態、初侵染來源、侵入途徑等。病情指數計算公式如下:


表1 水竹菱斑病病情試驗觀測樣點簡況
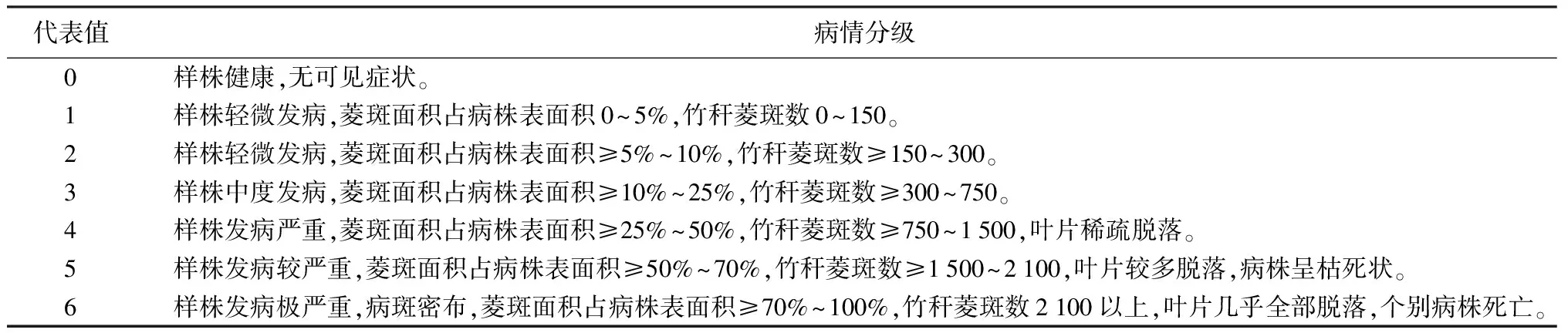
表2 水竹菱斑病病情指數分級標準
1.2 供試菌株
試驗菌株四川新小滴孢腔菌(Neostagonosporellasichuanensis)來源于四川農業大學微生物標本保藏室(SICAUCC),菌株編號SICAUCC 16-0001(模式菌株),分離自雅安市雨城區孔坪鄉九層巖水竹罹病枝條。
1.3 生物學特性試驗
不同培養基處理:無菌條件下,用直徑6 mm打孔器取病原菌菌餅分別接種于馬鈴薯葡萄糖瓊脂(PDA)、玉米粉瓊脂(CMA)、燕麥粉瓊脂(OMA)、馬鈴薯蔗糖瓊脂(PSA)、查氏瓊脂(CDA1)、胡蘿卜瓊脂(CDA2)和水瓊脂(WA)培養基中。培養基配方參見文獻[10]。每種培養基設置4個重復,置于25 ℃、12 h光照和12 h黑暗交替條件下培養。每5 d采用十字交叉法測量菌落直徑,共計測量8次。同時,觀察不同培養基上的菌落形態,40 d后,采用徒手制片法,借助顯微鏡觀察產孢情況等[11]。
不同溫度處理:無菌條件下,取直徑6 mm的菌餅接種于PDA培養基,分別置于恒溫培養箱中,5、15、25、35 ℃,12 h光照和12 h黑暗交替。每個處理設置4個重復,菌落直徑測量和產孢情況觀測方法同不同培養基處理。
不同pH處理:無菌條件下,分別使用1 mol·L-1NaOH和1 mol·L-1HCl調節PDA培養基pH為4.0、5.0、6.0、7.0、8.0、9.0和10.0。接種6 mm的菌餅置于25 ℃、12 h光照和12 h黑暗交替條件下培養。每個處理設置4個重復,菌落直徑測量和產孢情況觀測方法同不同培養基處理。
不同光照處理:無菌條件下,取直徑6 mm菌餅接種于PDA培養基,分別置于完全黑暗、12 h光照和12 h黑暗交替、完全光照的條件下,25 ℃恒溫培養。4個重復,菌落直徑測量和產孢情況觀測方法同不同培養基處理。
不同碳、氮源處理:以真菌生理培養基[12]為基礎培養基,配比為氮源1.0 g、磷酸氫二鉀(K2HPO4)1.0 g、硫酸鎂(MgSO4·7H2O)0.5 g、碳源5.0 g、瓊脂20.0 g、蒸餾水1 L。以KNO3為氮源,分別用5種碳源(葡萄糖、蔗糖、乳糖、麥芽糖、可溶性淀粉)配成含有不同碳源的固體培養基,以不加碳源的基礎培養基作為對照。置于25 ℃、12 h光照和12 h黑暗交替條件下培養,每個處理設置4個重復,菌落直徑測量和產孢情況觀測方法同不同培養基處理。以真菌生理培養基為基礎,以葡萄糖為碳源,分別用5種氮源(硝酸鉀、硝酸銨、牛肉膏、蛋白胨、酵母粉)配成含有不同氮源的固體培養基,以不加氮源的基礎培養基作為對照。置于25 ℃、12 h光照和12 h黑暗交替條件下培養,每個處理設置4個重復,菌落直徑測量和產孢情況觀測方法同不同培養基處理。
1.4 室內藥劑篩選方法
6種供試藥劑信息詳見表3。采用菌絲生長速率法測定供試藥劑的EC50[13]。將供試藥劑用紫外燈照射2~3 h除菌后,用無菌水分別稀釋為0.1、1.0、10.0、100.0 mg·L-1,按V(藥液)∶V(培養基)=1∶9的比例,將冷卻至50 ℃左右的PDA培養基混合均勻,制成不同質量濃度的含藥平板[14]。每皿接種直徑為6 mm的菌餅,4個重復,以不加藥劑只加同量無菌水的處理為空白對照,置于恒溫培養箱中25 ℃倒置培養。每5 d用十字交叉法測量并記錄菌落直徑,按如下公式計算抑菌率,利用概率值法[13]求出各藥劑的毒力回歸曲線,并計算EC50,同時觀察與記錄菌落形態等。


表3 供試藥劑的名稱及來源
1.5 數據處理
試驗數據用Microsoft Excel 2016整理,獲得毒力回歸方程并計算EC50,使用SPSS(24.0)軟件進行差異顯著性分析,采用Origin 8.5軟件繪制圖形,Photoshop 2017軟件處理圖片。
2 結果與分析
2.1 發病特點
經過1年的調查,結果顯示,3個樣地病情指數變化趨勢基本一致(見圖1),整體呈現上升趨勢。總體而言,孔坪鄉樣地水竹林病情指數高于其他2個樣地,上里鎮樣地水竹林病情指數相對較低。各樣地水竹林病情指數變化可細化為3個階段:2016年4月至8月,病情指數逐漸增加;2016年8月至12月,病情指數呈下降趨勢后出現小幅回升,當年生新竹的補充是病情指數下降的主要原因(每年3—4月份,水竹林開始發筍,經3~4個月的高生長期,即生長為成竹);2016年12月至翌年4月,隨著當年生新竹感病以及感病竹株進一步加重,病情指數再次增加。
11月至翌年4月,在水竹竹稈和枝條的迎風面,形成褐色至暗褐色小斑,小斑逐漸向四周擴展,形成菱形、近菱形、橢圓形或不規則形病斑,病斑常連成一片,有時散生,隨后病斑中央隆起,形成半球形至近球形子實體,即四川新小滴孢腔菌的子座。子囊果成熟后,子囊孢子從孔口溢出,借助風雨等途徑進行傳播與擴散。一般在2—4月期間,是子囊果形成和成熟以及子囊孢子擴散的集中時期;5—10月,以病原菌在病斑周圍危害為主,隨著持續侵染,病斑逐漸擴大并最終形成枯斑,期間不產生子座;11月,即進入下一輪侵染循環。分生孢子器在全年均有產生,常形成長梭形、長橢圓形或不規則形狀,與子座混生,或散生。子囊孢子和分生孢子是當年主要的初侵染源。一般連續侵染1~3 a,可致水竹整株布滿病斑,并最終枯萎死亡。
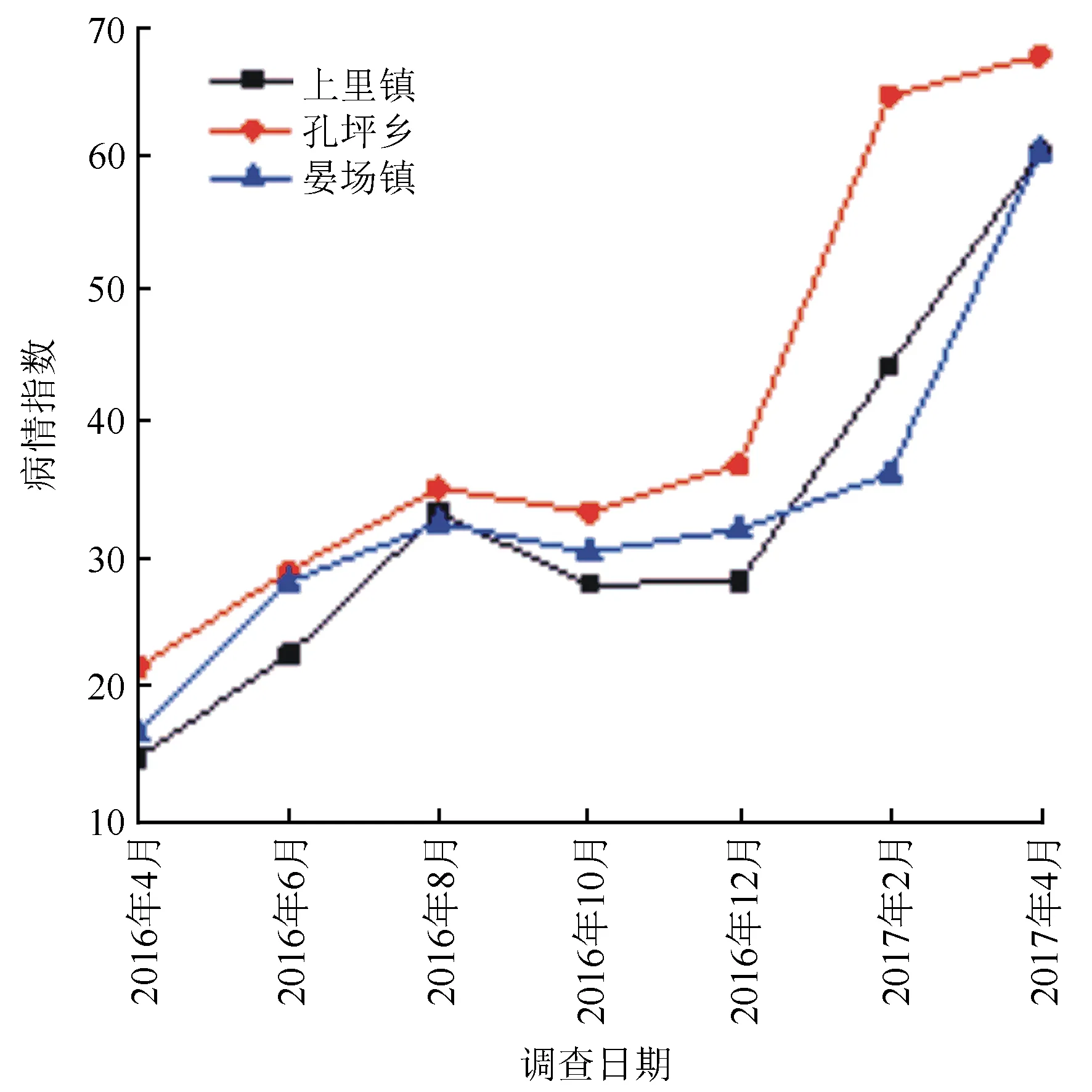
圖1 水竹菱斑病病情指數
2.2 生物學特性
2.2.1培養基對菌株SICAUCC 16-0001生長的影響
菌株在6種培養基上均能生長,且均未發現產孢現象,但菌落形態(見圖2)和生長速率存在差異。在PDA、OMA、CDA2培養基上菌落為圓形,邊緣整齊,表面有顏色不同的同心輪紋,正面為灰白色、黑灰色或褐色,背面多為灰褐色,質地干燥。顯微鏡觀察可見發達的氣生菌絲,有菌環、菌索等特殊結構,未見產孢結構。在PSA培養基上有水滴狀分泌物。在CDA1和WA培養基上菌落為黑褐色,菌絲稀疏。
在不同培養基中,菌落生長直徑差異顯著。CDA2培養基最適合菌株生長,培養40 d菌落直徑達到(6.23±0.29)cm,與其他處理之間差異極顯著(P<0.01);CMA和OMA培養基,培養40 d菌落直徑分別為(5.28±0.36)、(4.80±0.37)cm;PDA培養基,菌絲生長較慢,培養40 d菌落直徑達(4.00±0.37)cm;PSA、CDA1和WA培養基,菌絲生長緩慢,培養40 d菌落直徑分別為(2.90±0.22)、(2.77±0.25)和(3.03±0.21)cm,與其他處理之間差異顯著(P<0.01),但三者之間無顯著性差異。
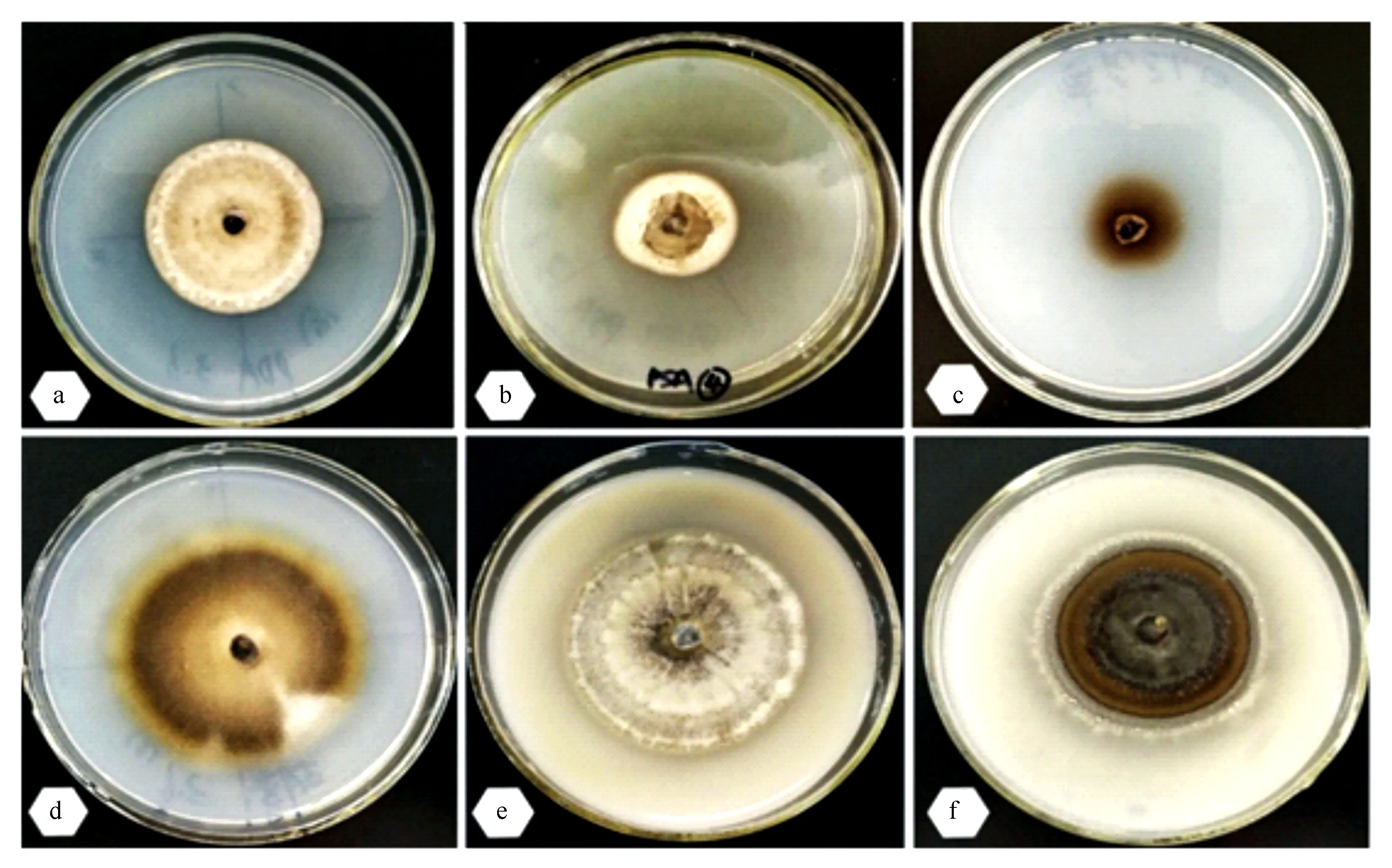
a.PDA;b.PSA;c.CDA1;d.CDA2;e.OMA;f.CMA。
2.2.2溫度對菌株SICAUCC 16-0001生長的影響
菌株在5~30 ℃均能生長,且均未觀測到產孢現象。試驗結果顯示,35 ℃下菌絲停止生長,且不同溫度下菌絲生長速度呈顯著差異(見表4)。在5、35 ℃條件下生長緩慢,15 ℃時生長速度加快,25 ℃時菌絲生長最快,培養40 d菌落直徑可達(3.90±0.26)cm,與5、35 ℃處理之間差異極顯著(P<0.01)。因此,25 ℃為該菌株最適生長溫度,較低和較高溫度下均不適宜該菌株生長。
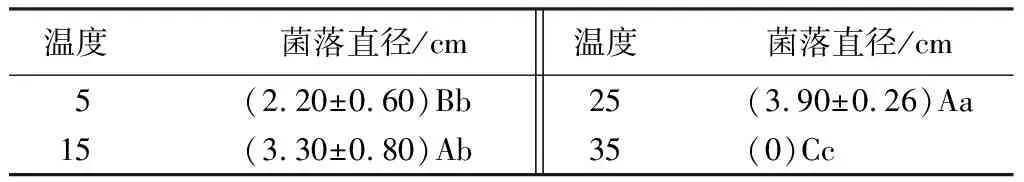
表4 不同溫度對菌絲生長的影響
2.2.3pH對菌株SICAUCC 16-0001生長的影響
pH=4~10菌株均能生長,且均未觀測到產孢現象。但不同pH對菌絲生長影響不同(見表5)。pH為6或7時最適合菌絲生長,培養40 d菌落直徑分別為(4.53±0.54)、(4.48±0.73)cm,與pH=5、pH=8、pH=9、pH=10處理差異顯著,與pH=4處理差異極顯著(P<0.01)。
2.2.4光照對菌株SICAUCC 16-0001生長的影響
菌株在全光照、12 h光照和12 h黑暗交替、全黑暗條件下均能生長,培養40 d菌落直徑分別為(3.58±0.50)、(5.03±0.86)、(3.65±0.34)cm,其中在交替光下菌絲生長最快,與其他處理之間差異顯著(P<0.05)。3種處理下菌落形態上有所不同,在黑暗條件下菌落中央有水滴狀分泌物產生,但均未觀測到產孢現象。
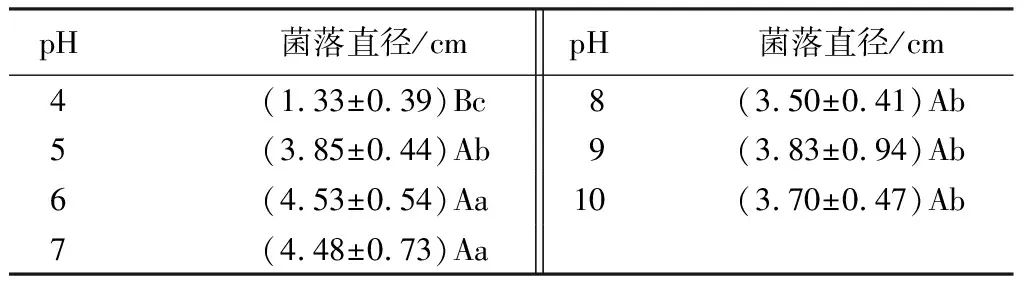
表5 不同pH對菌絲生長的影響
2.2.5碳、氮源對菌株SICAUCC 16-0001生長的影響
菌株在5種不同碳源條件下均能生長且均未發現產孢現象,但菌絲生長的速度不同(見表6)。其中以乳糖為碳源的培養基上菌絲生長最好,培養40 d菌落直徑可達(3.20±0.49)cm,與其他處理之間差異極顯著(P<0.01),為該菌株的最適碳源。其次為可溶性淀粉、蔗糖和麥芽糖,3種處理培養40 d菌落直徑最大可達(2.63±0.38)cm。而葡萄糖培養40 d菌落直徑僅達乳糖處理的78%。
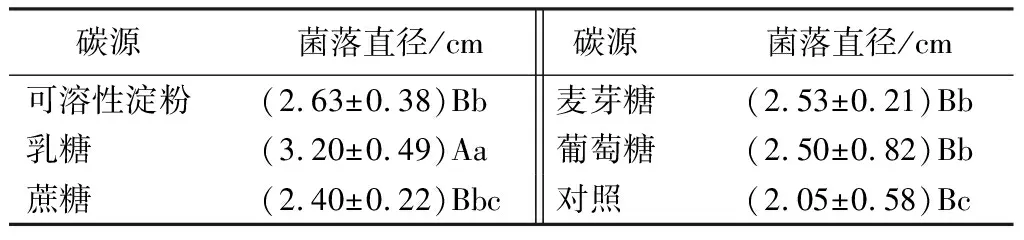
表6 不同碳源對菌絲生長的影響
菌株在5種供試氮源中均能生長且均未發現產孢現象,但是生長情況存在差異(見表7),以牛肉膏為氮源的培養基上菌絲生長最快,培養40 d菌落直徑可達(4.23±0.39)cm,與其他處理之間差異極顯著(P<0.01),為該菌株的最適氮源。其次是酵母粉和蛋白胨,培養40 d菌落直徑分別達到(3.50±0.75)、(3.40±0.42)cm。以硝酸銨為氮源時,菌絲生長最慢,顯著低于對照(P<0.05)。
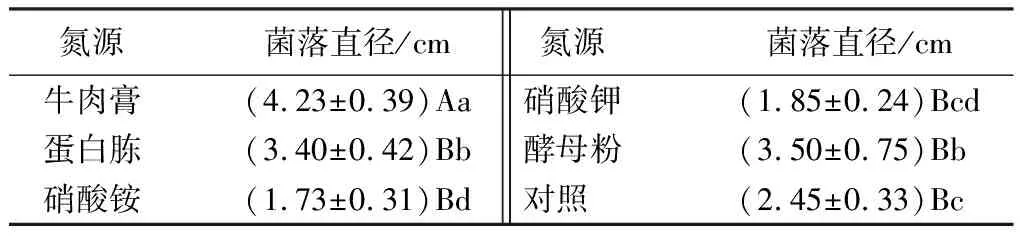
表7 不同氮源對菌絲生長的影響
2.3 不同室內藥劑抑制作用差異
6種供試殺菌劑對菌株生長均表現出不同程度的抑制作用。在菌落形態上,與空白對照相比,藥劑處理后的菌落邊緣不規則,有明顯的褶皺,整體顏色變淺。通過數據統計和擬合毒力回歸方程(見表8)可見:烯肟·戊唑醇對菌株的抑菌效果遠優于其他殺菌劑,EC50為5.64 mg·L-1,而其余5種藥劑的EC50為47.74~76.46 mg·L-1。6種殺菌劑的抑菌效果排序由高到低為:烯肟·戊唑醇、噁霜·錳鋅、霜脲·錳鋅、百菌清、甲基硫菌靈、精甲霜·錳鋅。
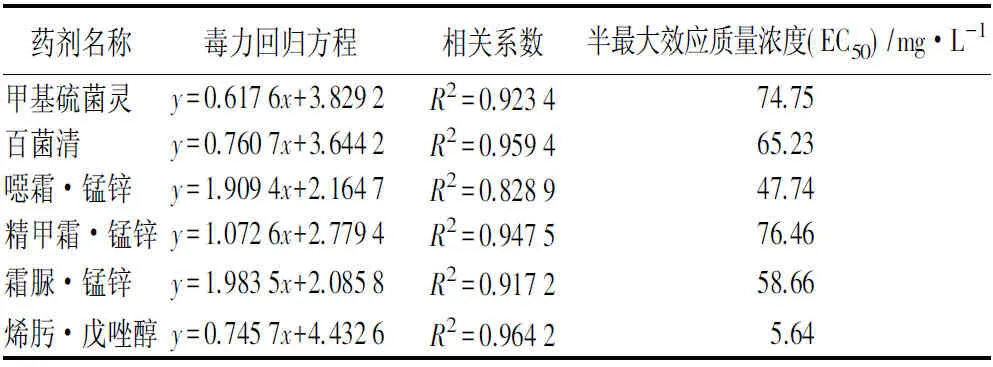
表8 6種殺菌劑對水竹菱斑病的毒力效應
3 討論
水竹菱斑病為竹類新病害,屬國內外首次報道,其主要致病菌為四川新小滴孢腔菌。水竹菱斑病發病的典型癥狀與竹類枯梢病或梢枯病截然不同[15-16],其具體表現為:致病菌不同,枯梢病病原為竹喙球菌(Ceratosphaeriaphyllostachydis),隸屬喙球殼科(Ceratosphaeriaceae),梢枯病病原為暗孢節菱孢(Arthriniumphaeospermum),隸屬梨孢假殼科(Apiosporaceae);危害部位不同,水竹菱斑病可危害竹稈、枝條和竹鞭(裸露),而枯梢病或梢枯病危害竹稈籜環處或側枝分枝處;病斑類型有別,水竹菱斑病病斑較小,呈菱形、近菱形、橢圓形或不規則形,常聚集成片,而枯梢病或梢枯病常在籜環處或分枝處向上向下擴展,形成較大的近菱形病斑。就致病力而言,類似毛竹枯梢病,水竹菱斑病有較強的致病性,是一類危害性極大的竹類枝稈病害[17],而雜交竹梢枯病可能為繼發性病害,凍霜害是主要誘因[18-19]。
病原菌的生物學特性研究結果表明,菌絲最適生長溫度為25 ℃,超過30 ℃時菌絲生長減緩,說明該菌對高溫敏感,這與該病害在田間表現出來的發病規律吻合。菌株最適生長碳源為乳糖,氮源為牛肉膏,這與曾慧蘭等[20]研究的結果相似,說明一般情況下乳糖和牛肉膏可以促進菌絲生長。在供試的6種培養基中,四川新小滴孢腔菌在CDA2、CMA、OMA培養基上生長較好,這與蔡吉苗等[21]得到的研究結果相似。菌絲在pH=4~10均能生長,但是pH=6~7最適合生長,這與王中武等[22]研究結果相近。在培養特性試驗中,均未發現該病原菌形成產孢結構,說明該病原菌的室內產孢條件可能較為特殊[23]。
從6種不同殺菌劑對四川新小滴孢腔菌的抑制效果來看,烯肟·戊唑醇對四川新小滴孢腔菌的抑制效果遠高于其他殺菌劑,其EC50值為5.64 mg·L-1。烯肟·戊唑醇是三唑類殺菌劑戊唑醇與甲氧基丙烯酸酯類殺菌劑烯肟菌胺的復配劑,研究表明,烯肟·戊唑醇對多種植物真菌病害具有良好的防治效果,如蘋果斑點落葉病(Alternariaalternata)[24]、豇豆根腐病(Fusariumcommune)[25]和桃樹褐斑穿孔病(Pruniphilomycescircumscissus)[26]等。同時結合野外調查發現,11月至翌年4月屬于病害侵染前期,也是病害防控的關鍵時期,可采取清除病枝枯竹或罹病組織等營林措施,有效降低初侵染源數量,同時輔以化學藥劑抑制病原菌的生長,防止病害的蔓延。此外,野外防控受制于多種因素,包括立地條件、氣候因子、人為因素等,具體的綜合治理技術還有待于進一步研究和評價。
結合野外調查與室內分離鑒定,在水竹菱斑病病斑周圍常伴隨其他2種致病菌的侵染,即存在復合侵染[27]的現象。水竹生擬卡氏革菌(Parakarsteniaphyllostachydis)和四川柄赤叢殼(Podonectriasichuanensis)[28-29],二者常在四川新小滴孢腔菌所致病斑的病健交界處生長,甚至覆蓋在菱形病斑表面,也可致使病斑向外圍進一步擴大。野外調查結合室內顯微觀察發現,在寄主植物上,2種病原菌可形成明顯的子座,生長發育受季節變化影響不大,1年可發生多代,無疑加重水竹病害的發生,加劇水竹林的衰退。3種病原菌的復合侵染機制還有待于深入研究,如病原間互作形式和驅動機制等。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
