土壤水分對烏桕秋葉生理指標及觀賞效果的影響1)
2021-05-27 08:43:52吳飛洋柳新紅王成龍呂江波張麗芳李因剛
東北林業大學學報 2021年5期
吳飛洋 柳新紅 王成龍 呂江波 張麗芳 李因剛
(浙江省林業科學研究院,杭州,310023)(浙江省建德市林業局)(浙江省林業科學研究院)
烏桕(Triadicasebifera(Linnaeus) Small.)為大戟科(Euphorbiaceae)烏桕屬(Triadica)高大落葉喬木,樹高可達15 m,樹干挺拔,葉形獨特,在主產區常被稱為木子樹、桕子樹[1]。烏桕天然分布于中國、日本及越南,在我國主要分布在黃河以南各省區,北達陜西、甘肅[2]。烏桕資源豐富,喜光喜水,多生長于塘邊、溪旁、曠野和向陽的山坡等地[3]。近年來,科研人員主要在烏桕抗逆、遺傳學等方面進行了大量研究[4-9],其葉片生理活性和藥理作用研究也有報道[10-11]。同時,烏桕作為工業原料樹種可應用于油漆和制皂等生產中,還可用來制備類可可脂和生物柴油[11-14]。
烏桕也是重要的秋色葉樹種,葉色從亮黃到紅紫變化豐富,觀賞價值高,在彩色森林構建和園林景觀造景中的應用也越來越多。彩葉植物豐富的葉色變化除受自身遺傳基因決定外,外部生態環境因子也對其產生重要影響。土壤水分是植物生長的重要條件,有研究發現干旱或淹水脅迫對烏桕幼苗生長造成影響,但其能通過自身調節來緩解脅迫傷害,且不同種源間存在差異[15-16]。研究水分脅迫對葉色的影響能為彩葉植物栽培應用提供實踐理論依據,對于提高其觀賞價值也具有重要意義。干旱或淹水脅迫會使植物葉片內的葉綠素和花色素苷質量分數發生變化,紅葉石楠(Photiniafraseri)在長期干旱環境下不利于花色素苷合成[17],而美國紅楓(Acerrubrum)和紅葉桃(Prunuspersicaf.alropurpurea)則在中度干旱下最利于葉片中花色素苷的積累[18-19]。本文以烏桕屬植物新品種‘浦大紫’無性系盆栽苗為材料,設置不同的水分梯度以研究葉片各相關生理指標的變化規律,并探討水分對烏桕秋季葉色觀賞效果的影響,以期為彩葉觀賞烏桕的合理高效栽培利用提供依據。
1 試驗地概況
試驗地位于浙江省杭州市西湖區的浙江省林業科學研究院試驗苗圃,地理位置為30°13′9″N,120°1′44″E,海拔30 m。該地屬亞熱帶季風氣候區,陽光充足、冬冷夏熱,歷年降水量為1 100~1 655 mm[20]。根據浙江杭州城市森林生態系統國家定位觀測研究站數據,試驗期間的日平均溫度由24 ℃逐漸降低至11 ℃,日平均濕度為67%~86%(表1)。
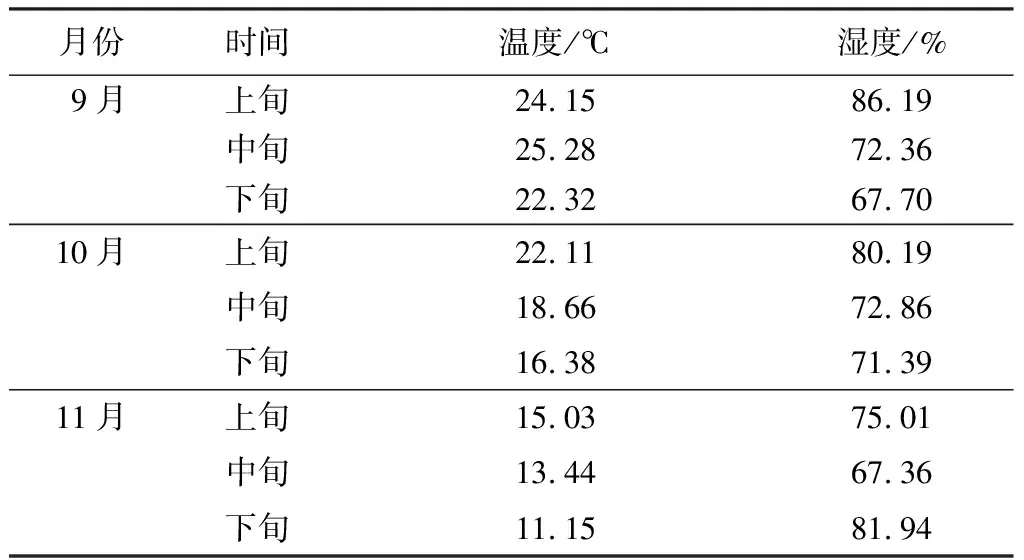
表1 試驗期間的溫濕度變化情況
2 材料與方法
2.1 試驗材料與設計
試驗以國家林業和草原局授權的烏桕屬烏桕植物新品種‘浦大紫’(品種權號20180397)的當年生嫁接苗為試驗材料。砧木為1年生烏桕實生苗,地徑0.7~1.0 cm,于2019年4月11日完成嫁接,接穗為‘浦大紫’。試驗期間,苗木定植于容量為2加侖(7.57L)的塑料花盆內,栽培基質為山地紅壤。試驗材料生長到9月時,株高為80~130 cm,冠幅為40~70 cm。
本試驗設置4種水分梯度處理,其土壤含水量分別為田間最大持水量的80%(對照組)、65%(輕度脅迫)、50%(中度脅迫)和35%(重度脅迫),分別用CK、LS、MS和SS表示。試驗采用隨機區組設計,每個處理重復3次,每個重復10株。按照試驗設計,參試苗木隨機擺放于開闊、平整的圃地上,彼此間保持0.6 m的間距。8月28日,在試驗苗木上方搭建2.0 m高的塑料遮雨棚,材質為白色塑料大棚膜,防止自然降雨對水分控制造成影響。9月1日開始進行水分脅迫處理,采用盆栽控水法于每天下午17:00時左右稱質量補水,使各處理維持在脅迫范圍內。
2.2 樣品采集與指標測定
于開始脅迫處理的第23、30、37、46、57 d進行取樣,每次取樣時間均為早上09:00—10:00,其中46 d為MS處理進入變色始期,57 d為SS處理進入落葉始期。每個脅迫處理的所有植株均進行采樣,在每個植株的側枝上采集從頂端往下第3至4輪葉片2~3片,混合后放入已經提前做好標記的塑料封口袋,然后立即將樣品帶回實驗室進行各指標的測定工作。
葉綠素和類胡蘿卜素質量分數測定采用丙酮浸提法[21],花色素苷質量分數測定采用鹽酸浸提法[22],可溶性糖質量分數測定采用蒽酮比色法[23]。葉片色度測定采用柯尼卡美能達小型色差計CR-10 Plus,并記錄L*、a*和b*值,其中L*值代表明亮程度,a*值代表紅綠色,b*值代表黃藍色。植物地徑的測定采用游標卡尺測離地10 cm處的數值,精度為0.01 cm。從植株主干上分化出來的枝條確定為一級枝條。
2.3 葉色觀賞期
葉變色期為苗木植株上50%葉片出現變色的時間,落葉期為植株上50%葉片落葉的時間,變色期至落葉期之間的天數為觀賞期。
2.4 數據處理
所得數據均用Excel 2010進行數據處理,SPSS 22.0進行方差分析,置信區間為95%,并用Origin2017進行制圖,文中圖表數據為平均值±標準差。
3 結果與分析
3.1 不同水分處理對葉綠素質量分數及其組成的影響
由表2可知,隨著脅迫持續時間的延長,不同水分處理對葉綠素a、葉綠素b和葉綠素(a+b)的質量分數均表現出不同程度的下降趨勢。其中,MS處理的葉綠素a、葉綠素b與葉綠素(a+b)質量分數在本試驗觀測期間內下降量最多,其次是SS處理;到第57 d,葉綠素(a+b)質量分數下降幅度大小的順序依次為MS、SS、LS、CK,分別比初期下降了81.0%、77.6%、53.5%和50.0%。
在試驗的各個階段,葉綠素質量分數在脅迫處理間均存在一定差異。在23 d時,3個脅迫處理與CK間的差異均達到了顯著性水平(P<0.05);到57 d時,3個脅迫處理仍與CK差異顯著(P<0.05),且MS、SS處理的質量分數顯著低于CK和LS。
w(葉綠素a)∶w(葉綠素b)值在前30 d穩定在3.0左右,之后開始逐漸增大至3.5左右;除了46 d時SS處理與其他處理間存在顯著性差異(P<0.05),在其余階段各個處理間均未表現出顯著性差異。
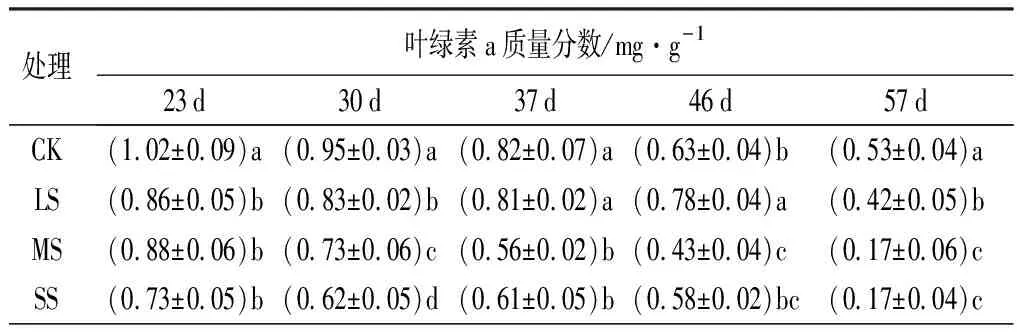
表2 葉片中各生理指標在不同水分處理下的變化情況

處理葉綠素b質量分數/mg·g-123d30d37d46d57dCK(0.34±0.03)a(0.32±0.03)a(0.26±0.02)a(0.19±0.02)b(0.15±0.02)aLS(0.30±0.01)b(0.28±0.02)b(0.27±0.01)a(0.23±0.01)a(0.12±0.01)bMS(0.29±0.01)b(0.24±0.01)bc(0.19±0.02)b(0.13±0)c(0.05±0.01)cSS(0.25±0.02)c(0.20±0)c(0.18±0.01)b(0.16±0.01)bc(0.05±0.01)c

處理葉綠素(a+b)質量分數/mg·g-123d30d37d46d57dCK(1.36±0.11)a(1.27±0.06)a(1.08±0.08)a(0.81±0.06)b(0.68±0.05)aLS(1.16±0.06)b(1.10±0.01)b(1.08±0.03)a(1.01±0.06)a(0.54±0.05)bMS(1.16±0.06)b(0.97±0.07)c(0.74±0.04)b(0.60±0.01)c(0.22±0.08)cSS(0.98±0.08)c(0.82±0.06)d(0.79±0.06)b(0.73±0.03)bc(0.22±0.05)c
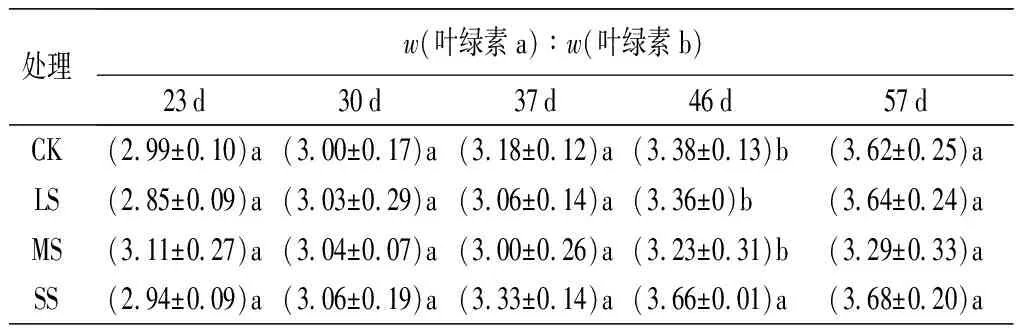
處理w(葉綠素a)∶w(葉綠素b)23d30d37d46d57dCK(2.99±0.10)a(3.00±0.17)a(3.18±0.12)a(3.38±0.13)b(3.62±0.25)aLS(2.85±0.09)a(3.03±0.29)a(3.06±0.14)a(3.36±0)b(3.64±0.24)aMS(3.11±0.27)a(3.04±0.07)a(3.00±0.26)a(3.23±0.31)b(3.29±0.33)aSS(2.94±0.09)a(3.06±0.19)a(3.33±0.14)a(3.66±0.01)a(3.68±0.20)a

處理類胡蘿卜素質量分數/mg·g-123d30d37d46d57dCK(0.40±0.03)a(0.56±0.05)a(0.38±0.03)b(0.36±0.05)ab(0.34±0.03)aLS(0.30±0.02)b(0.54±0.02)bc(0.45±0)a(0.43±0)a(0.32±0.02)aMS(0.29±0.03)b(0.51±0.02)bc(0.33±0.03)c(0.31±0)b(0.27±0.03)bSS(0.29±0.02)b(0.49±0.01)c(0.35±0.01)bc(0.34±0.01)b(0.33±0.02)a

處理花色素苷質量分數/U·g-123d30d37d46d57dCK(0.98±0.04)ab(1.64±0.01)a(4.84±0.43)b(8.93±0.57)b(11.06±1.08)cLS(1.03±0.06)a(1.34±0.05)b(2.03±0.13)c(5.26±0.19)c(11.85±0.65)cMS(0.92±0.05)bc(1.66±0.11)a(6.82±0.45)a(13.26±0.83)a(31.08±2.53)bSS(0.88±0.04)c(1.38±0.04)b(5.00±0.43)b(5.60±0.21)c(36.88±3.42)a
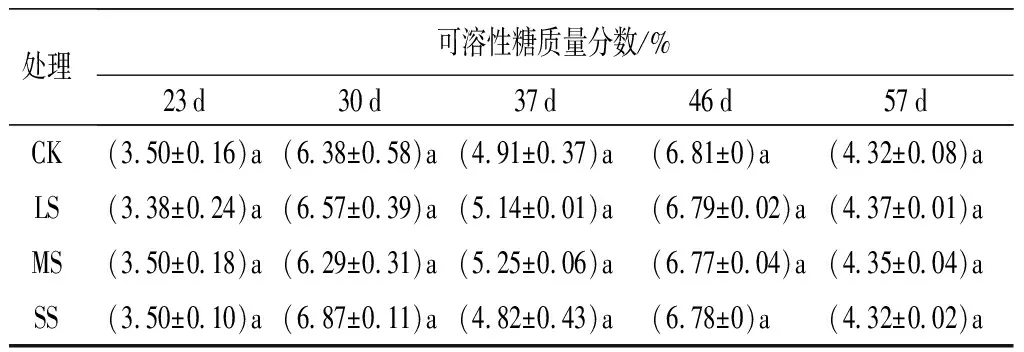
處理可溶性糖質量分數/%23d30d37d46d57dCK(3.50±0.16)a(6.38±0.58)a(4.91±0.37)a(6.81±0)a(4.32±0.08)aLS(3.38±0.24)a(6.57±0.39)a(5.14±0.01)a(6.79±0.02)a(4.37±0.01)aMS(3.50±0.18)a(6.29±0.31)a(5.25±0.06)a(6.77±0.04)a(4.35±0.04)aSS(3.50±0.10)a(6.87±0.11)a(4.82±0.43)a(6.78±0)a(4.32±0.02)a
3.2 不同水分處理對類胡蘿卜素和花色素苷質量分數的影響
由表2可知,隨著脅迫試驗的進行,類胡蘿卜素質量分數在各處理的變化趨勢一致,即先快速上升后緩慢下降,且均在30 d出現峰值。在試驗前30 d各脅迫處理均與CK有顯著性差異(P<0.05);到57 d時MS顯著低于其他3個處理(P<0.05),其他處理差異不顯著。
隨著烏桕秋季葉色逐漸變紅,花色素苷質量分數也逐漸升高。如表2所示,在脅迫初期葉未變色前,各處理的花色素苷質量分數均處于較低水平,僅0.95 U·g-1左右,之后開始逐步升高。在57 d之前,MS處理的花色素苷積累更多;但到57 d時,質量分數大小變為SS、MS、LS、CK,其中SS、MS均與CK有顯著差異(P<0.05),其質量分數分別是CK的3.33和2.81倍。
3.3 不同水分處理對可溶性糖質量分數的影響
可溶性糖是花色素苷合成的前體物質,也是影響葉色變化的一個重要指標。從表2可知,在試驗期間4個處理的可溶性糖質量分數呈現出波動起伏的變化,以先升后降的趨勢循環出現。雖然可溶性糖質量分數在試驗前后的波動變化較大,但是在試驗期間,3個脅迫處理及CK基本處于同一水平,彼此間并未表現出顯著差異。
3.4 不同水分處理對葉片色度值的影響
表3顯示,所有4個處理的色度值L*在前30 d均有不同程度的升高,從37 d開始逐漸下降;并且在前30 d各脅迫處理均與CK有顯著差異(P<0.05),從37 d開始到試驗結束只有SS與CK一直維持著顯著性差異(P<0.05)。57 d時,SS與MS的L*值分別比CK高了10.18%、13%,而LS處理與CK無顯著差異。
從表3能直觀發現,隨著試驗進行各處理的a*值均由負值逐漸接近正值,并明顯增大,說明烏桕葉色逐漸由綠變紅;在57 d各處理均轉為正值,葉色變化最為明顯。在試驗的第23天,各處理的a*值與CK都無顯著差別;但隨著葉色發生轉變,從37 d開始,各脅迫開始與CK表現出顯著性差異(P<0.05);到試驗的57 d,a*值大小依次為MS、SS、LS、CK。其中,MS和SS處理的a*值分別是CK的6.21、5.58倍。
在前30 d,各處理的b*值均表現為升高,在37 d略有回調后再繼續升高,到試驗結束時最終下降(表3)。在同一時間下,b*值均隨著脅迫程度的加劇而增大,即表現為SS、LS、MS、CK。在試驗過程中SS處理一直與CK保持著顯著性差異(P<0.05),MS處理除了46 d其余時間也都與CK具有顯著差異(P<0.05),而LS一直與CK無顯著差異。

表3 色度值在不同水分處理下的變化情況

處理色度值a?23d30d37d46d57dCK(-7.98±0.08)a(-7.64±0.11)a(-5.58±0.25)b(-2.77±0.26)b(4.84±0.83)bLS(-8.09±0.11)a(-8.18±0.02)b(-6.81±0.12)c(-4.05±0.34)c(5.86±0.16)bMS(-8.02±0.12)a(-7.70±0.09)a(-2.14±0.19)a(-0.99±0.07)a(30.06±1.94)aSS(-8.04±0.10)a(-8.01±0.11)b(-5.29±0.42)b(-4.82±0.37)c(27.02±2.60)a
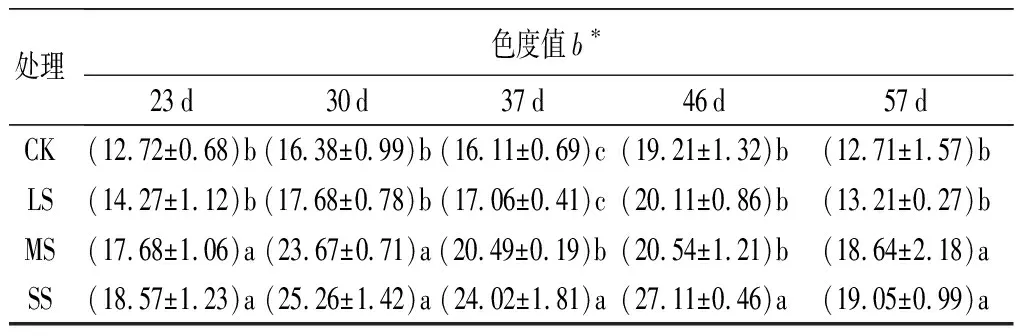
處理色度值b?23d30d37d46d57dCK(12.72±0.68)b(16.38±0.99)b(16.11±0.69)c(19.21±1.32)b(12.71±1.57)bLS(14.27±1.12)b(17.68±0.78)b(17.06±0.41)c(20.11±0.86)b(13.21±0.27)bMS(17.68±1.06)a(23.67±0.71)a(20.49±0.19)b(20.54±1.21)b(18.64±2.18)aSS(18.57±1.23)a(25.26±1.42)a(24.02±1.81)a(27.11±0.46)a(19.05±0.99)a
3.5 各指標間的相關分析
由表4知,在試驗進行過程中烏桕葉片的葉綠素(a+b)僅與類胡蘿卜素為極顯著正相關(P<0.01),而與花色素苷、色度值a*均呈極顯著負相關(P<0.01)。其中,類胡蘿卜素與花色素苷、色度值a*的關系分別為顯著負相關(P<0.05)和極顯著負相關(P<0.01),與可溶性糖為極顯著正相關(P<0.01)。花色素苷與色度值a*為極顯著正相關(P<0.01),可溶性糖、L*與b*值三者之間互為極顯著正相關(P<0.01)。
3.6 不同水分處理對苗木生長和觀賞期的影響
從表5可看出,試驗結束時參試苗木的地徑和一級枝條數在不同脅迫處理間表現出顯著差異(P<0.05)。其中地徑隨著脅迫程度的加劇而減小,4個處理彼此間的差異均達到顯著水平(P<0.05),SS、MS、LS分別比CK減小了2.31、1.54和0.69 mm。SS處理的一級枝條數均與SS、LS、CK有顯著差異(P<0.05),而SS、LS、CK彼此間均無顯著差異。總體而言,水分脅迫對植株生長造成一定程度的影響,且重度脅迫下的影響最大,生長狀況最差。
通過觀測發現,隨著溫度的逐步下降(表1),MS處理從第49天(10月19日)開始進入觀賞期,SS則在處理第51天(10月21日)進入。到第60天(10月30日)時,SS處理的烏桕已經進入落葉期,觀賞期僅9 d;而此時MS處理的烏桕葉已完全變色,處于觀賞盛期;LS和CK處理也開始變色進入觀賞期。第68天(11月7日)時,SS處理下的烏桕已基本完全無葉;MS處理開始進入落葉期,從變色到落葉19 d;LS和CK處理已完全變色,均進入最佳觀賞期。之后,對CK和LS處理的葉色和落葉進行持續觀測,發現二者的最終觀賞期分別為23、21 d。
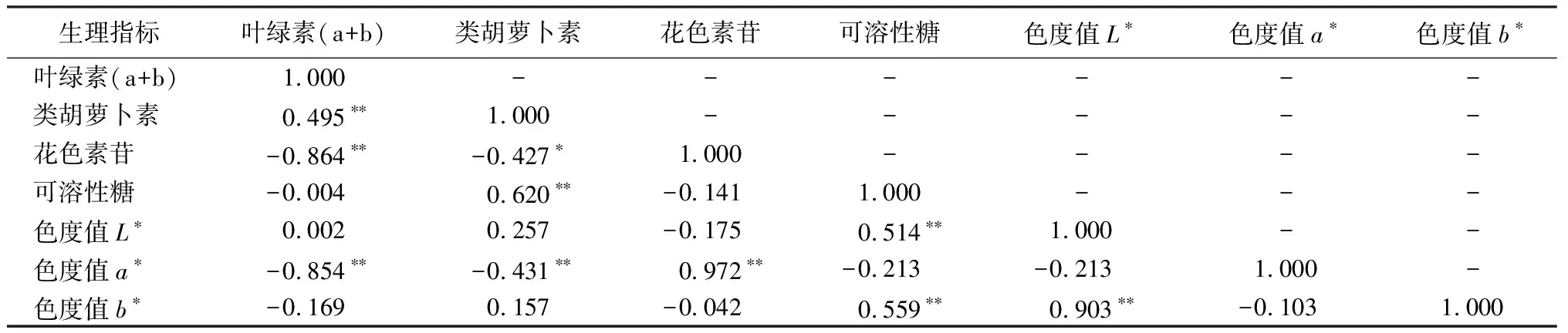
表4 烏桕生理指標間的相關性分析

表5 不同處理的苗木生長和觀賞期
4 結論與討論
彩葉植物的葉色表達是葉綠素、類胡蘿卜素和花色素苷等多種色素綜合作用的結果,如三葉海棠(Malussieboldii)秋季葉色發生變化正是由于色素質量分數及其比例的改變[24]。研究表明,適度的干旱條件能誘導花色素苷合成[25]。本試驗發現,隨著試驗進行所有處理的花色素苷質量分數大幅升高,同時葉綠素質量分數(包括葉綠素a、葉綠素b)均逐漸減少,而類胡蘿卜素質量分數雖然在初期呈現為升高但最終緩慢下降,這與前人的研究結果一致[16]。不同水分條件對葉色呈現是有差異的,從各色素的變化情況來看,MS和SS處理下的葉色變化較快,具體表現為到57d時花色素苷積累量最多(由大到小分別是SS、MS、LS、CK),這與美國紅楓的研究結果相符[18];葉綠素降解量也較多(由大到小分別是MS、SS、LS、CK),且MS和SS處理均與CK、LS存在顯著差異(P<0.05);但在前期MS的花色素苷質量分數積累更早,而LS處理到57d時其葉綠素質量分數和花色苷合成量雖然分別低于和高于對照組CK,但均未達到顯著差異水平。這表明水分脅迫能促進花色素苷的合成和葉綠素的降解,但在MS(中度)和SS(重度)脅迫下葉色變化更快更明顯。此外,在本試驗中葉綠素a與葉綠素b比值在前30 d穩定在3.0左右,但之后出現增大趨勢,隨著脅迫時間延長葉綠素b的降解速度大于葉綠素a。
可溶性糖是合成花色素苷的前體物質和能量提供者,同時也是重要的信號物質,起到多方面的作用[26]。在本次試驗中可溶性糖質量分數變化波動較大,逐步升高后又下降,而且各脅迫處理與CK間并未有顯著差異。其原因可能是可溶性糖不僅為花色素苷合成提供能量,也為其他的生理活動提供能量,因此其質量分數會受到多方面的影響。
色度值L*、a*和b*能通過數值變化精確反應出葉色變化情況。在試驗過程中,所有處理的亮度值L*均大致表現為先上升后下降的趨勢;試驗到57 d時,MS、SS處理與CK有顯著差異(P<0.05),并且MS處理的亮度值更高,說明葉色已經由綠色轉為更明亮的顏色色,視覺效果更好。紅綠值a*在前30 d變化波動不大,從37 d開始各處理的a*值均快速升高,表明葉色逐漸變紅,這與花色素苷的變化規律基本一致;到57 d時已從負值轉正,葉色已全部為紅色,其中升高幅度最大的是中度脅迫MS,與前人的試驗結果相符[27],其次才是SS、LS和CK,說明在本試驗下MS處理的葉色更紅艷。相關性分析結果中,a*與所有色素均極顯著相關(P<0.01),其中與花色素苷為極顯著正相關(P<0.01)。這是由于a*的大小是葉色紅度的表現,而花色素苷的升高是葉色顯紅的內在原因。隨著試驗進行,所有處理的黃藍值b*總體呈現先上升后下降的趨勢,并且在同一時間b*均隨著脅迫程度的加劇而加大,由大到小順序為SS、MS、LS、CK,這可能是由于水分脅迫影響到了葉片內的含水量,并且SS在試驗期間一直都與CK處理具有顯著差異(P<0.05),在重度脅迫下葉色更易變淡發黃,視覺觀賞效果差。
水分脅迫對植物生長產生了不利影響,通過測得的地徑和一級枝條數發現,隨著脅迫程度的加劇地徑越來越小,且4個處理彼此間的差異均達顯著水平(P<0.05);而一級枝條數僅SS與其他處理間有顯著差異(P<0.05)。總的來說SS處理下的地徑最細、一級枝條數最少,進而致使整個植株的葉片稀少[28],觀賞效果變差。從觀賞期來看,SS處理的烏桕落葉早,觀賞期僅有9 d;MS處理最早開始變色,觀賞期為19 d且葉片數量影響較小;而LS和CK處理的觀賞雖然相對較長,但其變色較晚,推遲變色將更容易遭受到氣候條件的劇烈變化,因氣溫驟降或大風導致提前落葉而縮短觀賞期。
因此從本試驗來看,在烏桕秋季葉色變化過程中,花色素苷和葉綠素的質量分數變化對葉色的影響較大,并且中度和重度水分脅迫能在一定程度上促進各色素變化,加快葉色呈現;但相較之下,中度脅迫下的葉色變化會更早,呈色也會更明亮紅艷,視覺觀賞效果好,并且觀賞周期長;而重度脅迫下不僅觀賞期短,生長情況也更差。本研究結果可為烏桕彩葉品種的科學合理栽培、管理撫育和觀賞應用等方面提供參考價值。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國生殖健康(2019年2期)2019-08-23 08:12:08
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
產品可靠性報告(2017年7期)2017-09-05 09:49:12
汽車觀察(2016年3期)2016-02-28 13:16:26
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
