杜鵑花品種間雜交F1代主要形態性狀變異1)
2021-05-26 07:56:50蘇鳴夏溪張春英奉樹成倪穗
東北林業大學學報 2021年5期
蘇鳴 夏溪 張春英 奉樹成 倪穗
(寧波大學,寧波,315800)(上海植物園)(寧波大學)
杜鵑花是杜鵑花科(Ericaceae)杜鵑花屬(Rhododendron)植物的總稱,是著名觀賞植物類群、我國十大名花之一[1]。杜鵑花資源豐富,觀賞價值高,是城市綠化、庭院美化的重要植物類群。據統計,全世界杜鵑花屬植物約967種,其中562種原產中國[2],我國西南山區是杜鵑花資源最重要的發源地和集聚中心[3]。根據英國皇家園藝學會(RHS)統計,全世界登錄的杜鵑花品種已經超過3萬種,但是絕大多數杜鵑花品種卻源自歐美和日本[4-6]。雖然近年來我國陸續有杜鵑花新品種推出[7-9],但是截至2020年國家林業和草原局植物新品種保護辦公室審定的杜鵑花品種數量剛剛突破百種。
隨著我國經濟水平的提高、生態文明和美麗中國建設的推進,人們對觀賞植物品種多樣性的需求不斷增加。我國長江流域及以南地區園林綠地中應用最多的杜鵑花是錦繡杜鵑(Rhododendronpulchrum)類品種[10],抗性強,栽培容易,在綠化市場備受青睞。由于國內露地栽培杜鵑花抗逆好的品種少,可用于綠地栽培的杜鵑花品種十分單調,無法滿足綠地品種多樣性的需求,因此如何快速培育特色各異的抗逆品種成為杜鵑花產業發展亟待解決的問題。目前,雜交育種是國內外杜鵑花新品種培育中最有效、最常用的方法之一[10]。雜交后代主要性狀變異規律的預測是親本選配和提高育種效率的關鍵。常見觀賞植物如菊花(Chrysanthemumspp.)[12]、紫薇(Lagerstroemiaspp.)[13]、月季(Rosaspp.)[14]、百合(Liliumspp.)[15]等雜交子代性狀變異規律均有大量研究,關于杜鵑花雜交后代性狀變異分析的文獻卻很少,僅見劉曉青等[16]對23個雜交組合F1代主要觀賞性狀的遺傳表現進行了簡單分析,馬紹宇等[17]對高山杜鵑1個雜交群體F1代主要觀賞性狀的遺傳分析,de Keyser et al.[18]對4個常綠杜鵑雜交組合F1代的花及葉片特征進行了QTL定位圖像分析。因此,本研究選擇了抗逆性較好的杜鵑花品種作為親本進行雜交,并獲得雜交子代開花植株。對3組杜鵑花品種間雜交F1代主要形態性狀進行觀測,分析雜交后代主要形態性狀變異特點及規律,為杜鵑花新品種選育和雜交親本的合理選配提供參考。
1 材料與方法
1.1 試驗材料
杜鵑花品種‘紫鶴’(Rhododendronpulcherum‘Zihe’)與‘紅蘋果’(Rh. ‘Red Apple’)、‘白鶴’(Rh. pulcherum ‘Baihe’)與‘粉紅泡泡’(Rh. ‘Pink Bubble’)、‘紫波’(Rh. ‘Zibo’)與‘麒麟’(Rh. ‘Kirin’)3組雜交組合的親本及穩定開花的雜交F1子代植株。雜交于2011年在浙江省寧波市北侖萬景杜鵑良種園進行,雜交子代在2015年3月開花。雜交親本的主要形態特征見圖1和表1。

a.‘紫鶴’;b.‘紅蘋果’;c.‘白鶴’;d.‘粉紅泡泡’;e.‘紫波’;f.‘麒麟’。
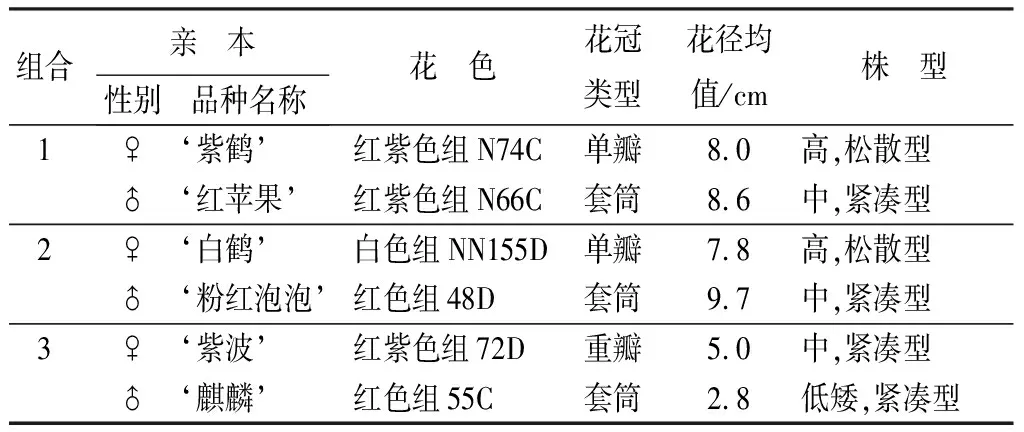
表1 杜鵑品種間雜交親本的主要形態特征
1.2 試驗方法
表型性狀的選取:2018—2019年對3組雜交組合的親本及部分F1子代的表型性狀進行調查和統計。根據中華人民共和國林業行業標準LY/T 1852—2009《植物新品種特異性、一致性、穩定性測試指南 杜鵑花屬 映山紅亞屬和羊躑躅亞屬》中相關內容選取5個數量性狀,4個質量性狀,共計9個性狀進行統計,其中花性狀6個,葉性狀3個(見表2)。
數據采集方法:數量性狀采用直尺直接測量法,并重復3次取平均值。統計花相關數據時,選擇同一植株不同位置的3朵花進行測量記錄;統計葉相關數據時,選擇同一植株上3片從上向下數第3層成熟無病害葉片進行測量記錄。質量性狀采取觀察分類法,其中花色以《英國皇家園藝協會的色譜》為標準,分為白色、紫粉、紅粉、紅色、紫紅、紫色、復色共7類,花瓣形狀和花冠類型以中華人民共和國林業行業標準LY/T 1852—2009《植物新品種特異性、一致性、穩定性測試指南 杜鵑花屬 映山紅亞屬和羊躑躅亞屬》進行分類和數量化處理,數據數量化編碼見表2。

表2 杜鵑花品種表型性狀分類及編碼
1.3 數據分析
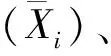
中親值:MPS=(P1+P2)/2,P1、P2為親本各表型性狀實際測量值。
2 結果與分析
2.1 花性狀變異
2.1.1 花色
3個雜交組合的花色變異系數平均值為38.65%,達到較大變異水平,不同組合之間差異大(見表3)。其中‘紫波’與‘麒麟’雜交子代的花色變異系數最大,為52.71%,雙親花色分別是紫紅和紅粉,子代花色分離廣泛,出現了白色、紫粉、紅粉、紅色、紫紅、復色(整體白色,瓣尖淡粉色)共計6種花色,所占比例分別是6.50%、25.80%、29.00%、12.90%、6.50%、19.40%,與父本花色相近的紅粉色花數量最多。‘白鶴’與‘粉紅泡泡’雜交子代的花色變異系數為36.06%,屬于較大變異。雙親花色為白色和紅粉,子代花色為白色、紫粉和紅粉。子代花色中占比最多的是和父本花色相同的紅粉,為48.40%,新花色“紫粉”占子代總數的29.00%。‘紫鶴’與‘紅蘋果’雜交子代的花色變異系數為27.17%,屬于中等變異,親本花色都是紫紅色,子代花色為紫粉、紫紅和紫色,其中以與親本相同的紫紅最多,占所有子代總數的66.70%。
2.1.2 花冠裂片內紋飾形態
花冠裂片內紋飾形態是杜鵑花的重要觀賞性狀之一,3個雜交組合變異程度差異較大(見表4)。其中,‘紫鶴’與‘紅蘋果’和‘白鶴’與‘粉紅泡泡’2個組合的親本花冠裂片內均有斑點,但是只有‘白鶴’與‘粉紅泡泡’雜交組合出現少量無斑點子代,占總數的6.45%,變異系數為12.90%。父本‘麒麟’花冠裂片內無斑點,母本‘紫波’花冠裂片內有斑點,子代無斑點和有斑點的植株分別占總數的38.71%和61.29%,多數子代具有與母本性狀類似的花冠裂片有斑點的特性。
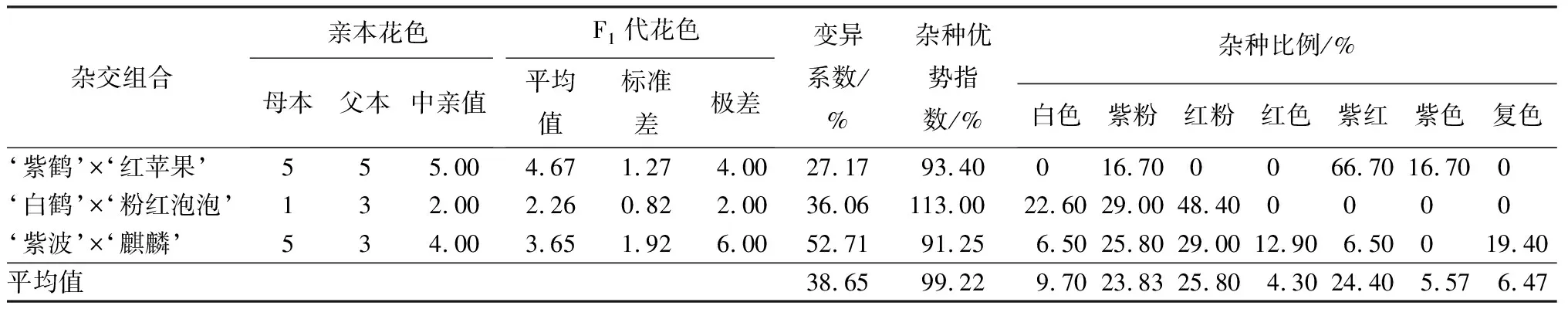
表3 杜鵑花品種間雜交F1代花色變異情況
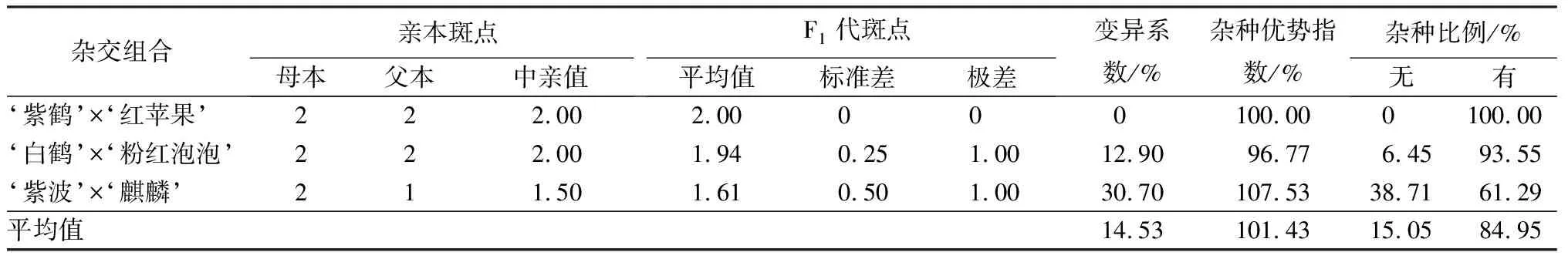
表4 杜鵑花品種間雜交F1代花冠裂片內斑點變異情況
2.1.3 花冠類型
3個雜交組合的花冠類型變異系數差別較大(見表5)。‘紫鶴’與‘紅蘋果’和‘白鶴’與‘粉紅泡泡’2個組合的母本均為單瓣,父本均為套筒,但子代花冠類型的分離情況完全不同。‘紫鶴’與‘紅蘋果’子代全部為與父本相同的套筒花,變異系數為0,而‘白鶴’與‘粉紅泡泡’組合的變異系數為35.70%,子代中單瓣和套筒占總數的比例分別是61.29%和38.71%,多數子代表現為與母本相同的單瓣花冠類型。‘紫波’與‘麒麟’組合的母本花冠類型為重瓣,父本為套筒,子代花冠類型中單瓣、套筒、半重瓣套筒和重瓣的比例分別是16.13%、35.48%、38.71%、9.68%,其中單瓣和半重瓣套筒是與親本不同的花冠類型,共占子代總數的54.84%,變異系數為36.78%,屬于較大變異。
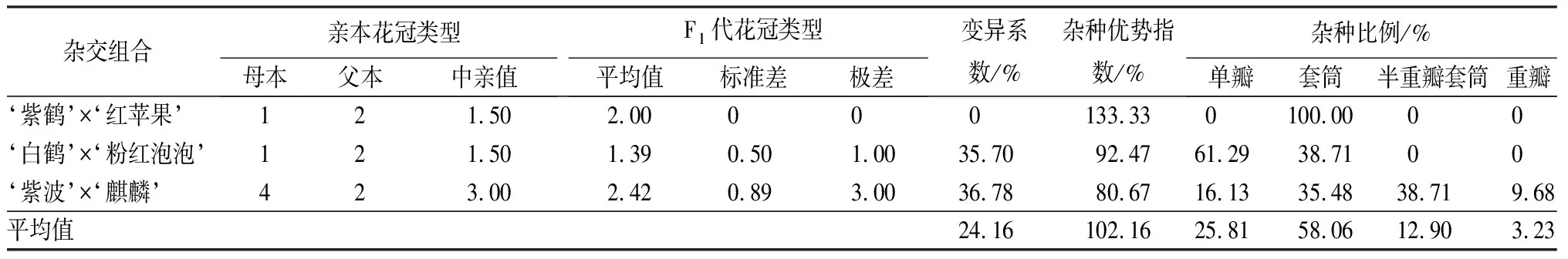
表5 杜鵑花品種間雜交F1代花冠類型變異情況
2.1.4 花瓣形狀
3個雜交組合親本均為橢圓形花瓣,子代中橢圓形花瓣的數量占比最大,是77.42%。但是不同雜交組合的子代變異系數差別較大(見表6)。其中,‘紫鶴’與‘紅蘋果’組合的變異系數為0,子代均為橢圓形花瓣;‘白鶴’與‘粉紅泡泡’組合的變異系數為16.01%,屬于中等變異,并出現了新的闊橢圓形花瓣性狀,占總數的12.90%;‘紫波’與‘麒麟’組合子代性狀中窄橢圓形、橢圓形、闊橢圓形和卵形分別占總數的3.23%、45.16%、29.03%和22.58%,新出現的花瓣性狀占總數的54.84%,變異系數為31.73%,屬于較大變異。
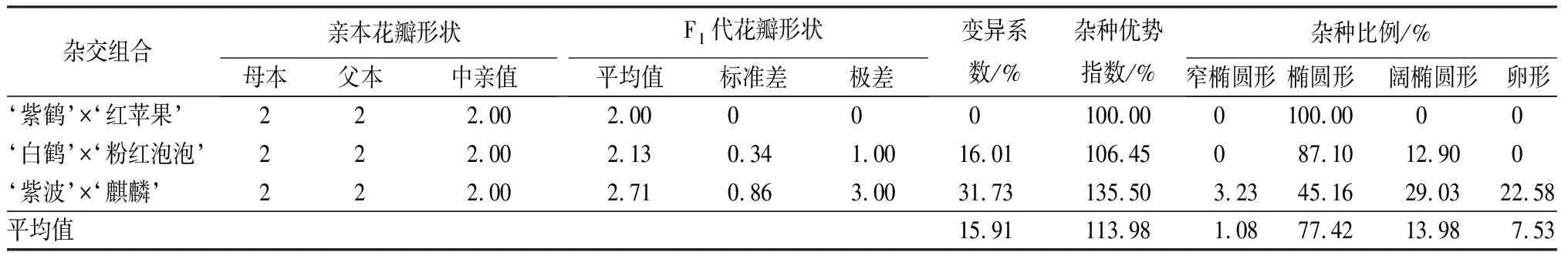
表6 杜鵑花品種間雜交F1代花瓣形狀變異情況
2.1.5 花徑
3個雜交組合花徑的變異系數較小且相近,為9.58%~11.41%,但是雜種優勢度各不相同(見表7)。‘紫波’與‘麒麟’雜交子代的雜種優勢指數最大,為125.90%,呈現明顯的超親本現象,高親比例達35.48%,沒有出現低于雙親的個體;其次‘紫鶴’與‘紅蘋果’組合雜種優勢指數為104.98%,超親本子代占60.00%,,其中高親比例達46.67%;‘白鶴’與‘粉紅泡泡’組合雜種優勢指數為92.98%,子代花徑介于雙親的為61.29%,低于雙親的為35.48%,高于雙親為3.23%。從子代花徑分布情況可見,不同雜交組合分布各不相同,超親子代較多,但是沒有明顯偏向于父本或母本。
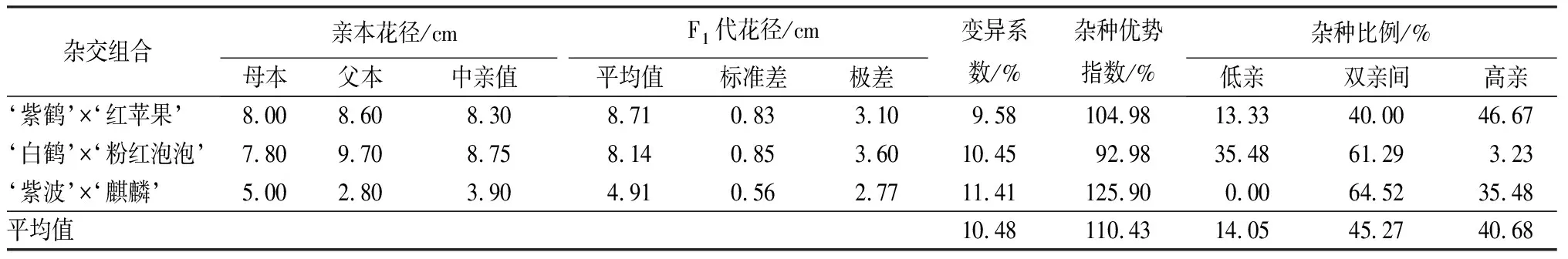
表7 杜鵑花品種間雜交F1代花徑變異情況
2.1.6 花梗長度
3個雜交組合花梗長度的變異系數差異不大,平均變異系數為17.59%,達到了中等變異水平(見表8)。但是3個組合的雜種優勢指數差異較大,‘紫鶴’與‘紅蘋果’組合的雜種優勢指數最大,為140.14%,組合中高親子代占子代總數的86.67%;‘紫波’與‘麒麟’組合的雜種優勢指數最小,為86.15%,組合中96.77%的子代介于親本之間;‘白鶴’與‘粉紅泡泡’組合雜種優勢指數為129.89%,但是這個組合子代高親比例最高,為90.91%。從子代花梗長度分布情況可見,親本花梗長度差異越小,子代高親數量越多;親本花梗長度差異較大時,子代多介于親本之間。
2.2 葉部性狀的遺傳
葉部性狀包括葉長、葉寬和長寬比3個性狀(見表9和表10)。‘紫鶴’與‘紅蘋果’、‘紫波’與‘麒麟’2組雜交組合的子代葉片生長情況類似,表現為葉長、葉寬的雜種優勢指數呈下降趨勢,長寬比的雜種優勢指數呈上升趨勢,表明這2個雜交組合子代的葉片整體變小,變得更加窄長。雜交組合‘白鶴’與‘粉紅泡泡’與其他2組相反,葉長和葉寬的雜種優勢指數呈上升趨勢,長寬比呈下降趨勢,表明這組雜交組合的子代葉片整體變大,變得更加寬短。從葉長、葉寬和葉長寬比子代分布情況看,葉部性狀沒有偏向于親本中的任何一個。
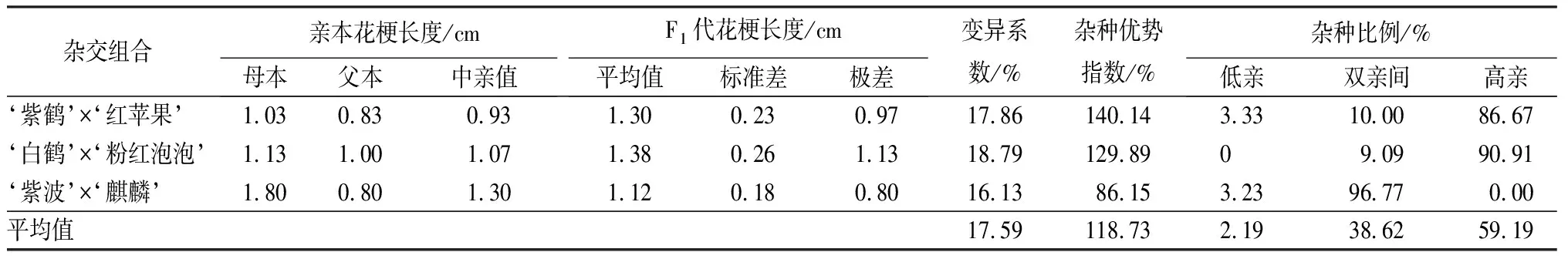
表8 杜鵑花品種間雜交F1代花梗長度變異情況
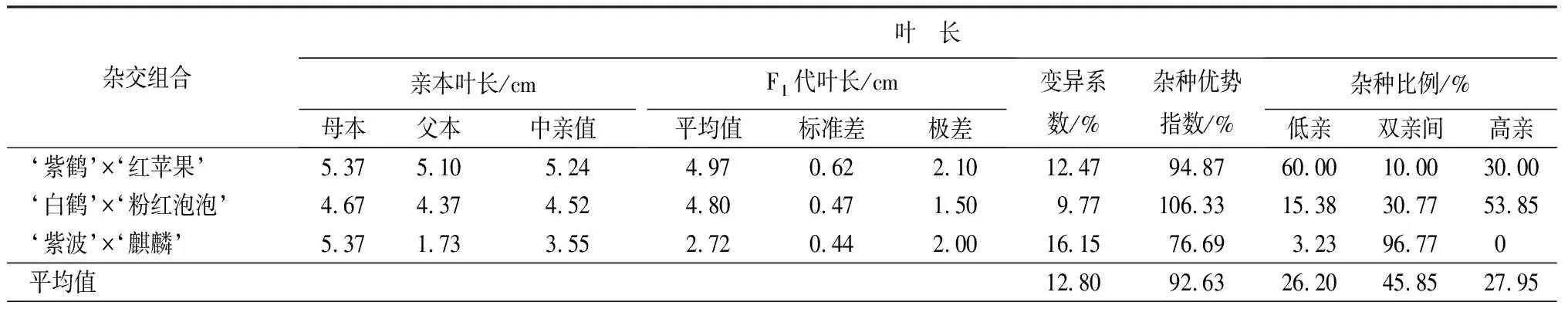
表9 杜鵑花品種間雜交F1代葉片性狀變異情況
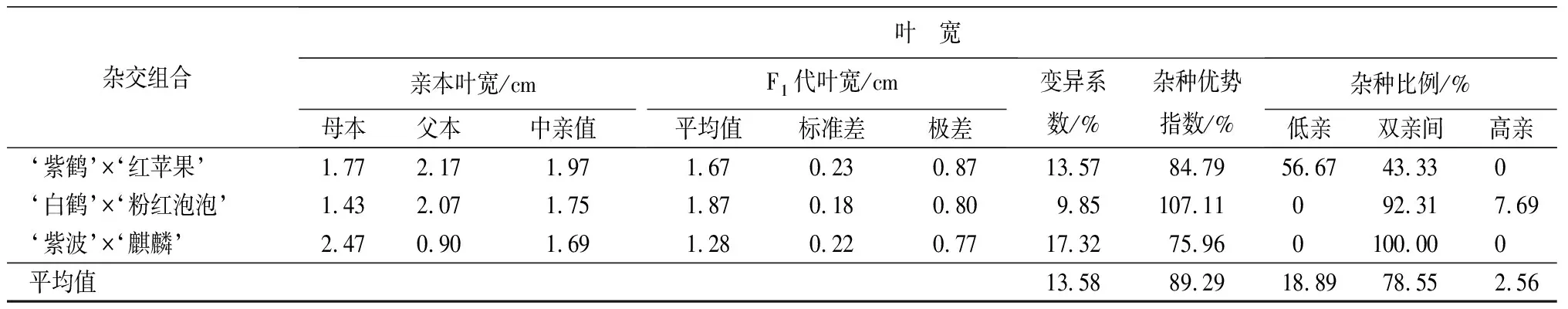
雜交組合葉 寬親本葉寬/cm母本父本中親值F1代葉寬/cm平均值標準差極差變異系數/%雜種優勢指數/%雜種比例/%低親雙親間高親‘紫鶴’ב紅蘋果’1.772.171.971.670.230.8713.5784.7956.6743.330 ‘白鶴’ב粉紅泡泡’1.432.071.751.870.180.809.85107.11092.317.69‘紫波’ב麒麟’2.470.901.691.280.220.7717.3275.960100.000平均值13.5889.2918.8978.552.56
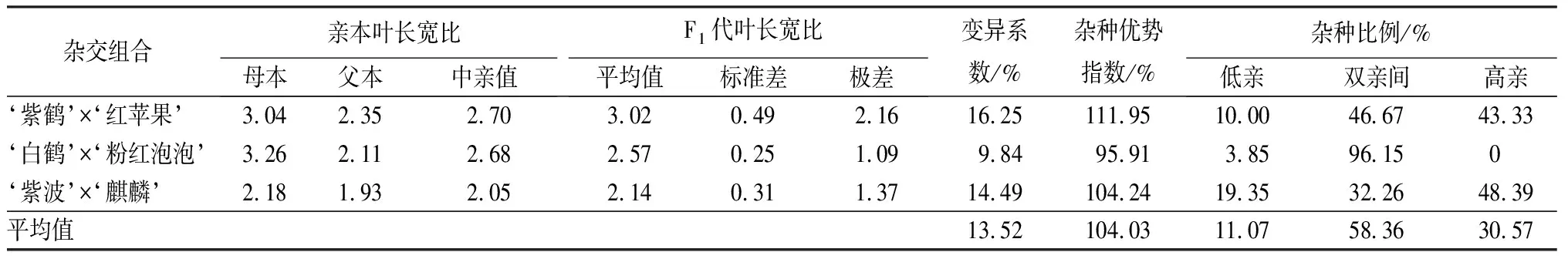
表10 杜鵑花品種間雜交F1代葉長寬比變異情況
2.3 雜交組合F1代數量性狀間的相關性
據相關性分析結果(見表11),3個雜交組的數量性狀之間具有較強的相關性,營養生長和生殖生長密切相關。花徑與花梗長度、葉長、葉寬、葉長寬比都呈極顯著相關,其中與葉長的相關性最強,相關系數為0.819。花梗長度與葉長、葉寬呈極正相關,與葉長寬比呈正相關。葉長與葉寬、長寬比均呈正相關。
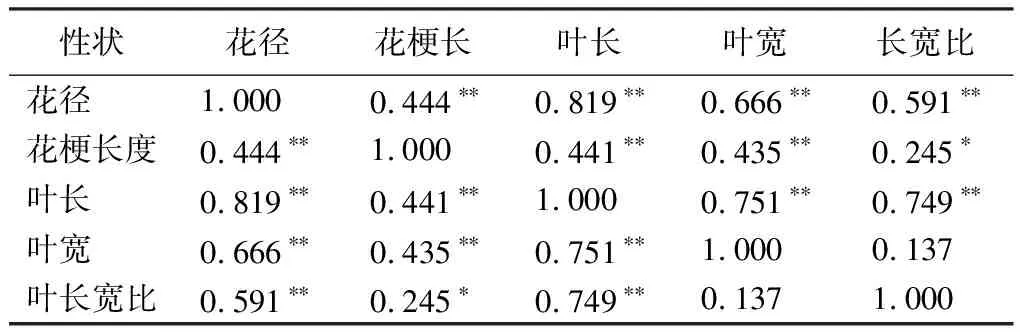
表11 雜交組合F1代數量性狀間的相關性
3 結論與討論
花色是杜鵑花育種中最重要觀賞性狀。本研究中花色劃分為白色、紅粉、紅色、紫紅、紫粉、紫色和復色共7類,按照色系歸為白色系、紅色系、紫色系和復色系共4個色系。3個雜交組合子代花色均出現了豐富的變異,當雙親花色差異越大時,子代花色變異也更為豐富,而且與父本花色類似的子代占比最高,說明父本花色對子代的花色影響較大。同時結果也表明,不同雜交組合的子代花色分離出現明顯差異。雜交組合‘紫鶴’ב紅蘋果’中親本花色均屬紫色系,子代均呈現深淺不同的紫色或紫紅色。雜交組合‘白鶴’ב粉紅泡泡’中母本是白色系,父本是紅色系,子代中白色花色占比為22.6%,遠低于粉紅色子代的比例,與劉曉青等[16]推測白色是杜鵑花的弱性基因觀點一致。該組合子代還出現了親本沒有的紫色系花色,可能紫色的基因來源于父本‘白鶴’。因為父本‘白鶴’是錦繡杜鵑品種之一,據Riek et al.[20]分析,錦繡杜鵑的親本之一為毛白杜鵑(Rh.mucronatum),雖然花色為白色花,但攜帶紫色基因。‘紫波’ב麒麟’雜交子代花色變異最為豐富,子代除了親本紫色、紅色系花色外還出現了少量的白色系和復色系,而劉曉青等[16]認為親本中有白色系和復色系,子代才會產生白色系和復色系,與本研究結果不一致。花色呈現與花朵中的花色苷種類有關,根據HPLC分析,紅色花含有2~4個主要的花色苷類型,而紫色花有2~6個類型[20],所以紫色花后代花色分離更為豐富。杜鵑花花色呈現不但與花色苷種類有關,也與由花色苷形成的色素量有關。杜鵑花花色遺傳是由3對主效等位基因控制的[20],具有質量遺傳特點,但是子代花色深淺差異,說明色素含量是有差異的,又具有數量遺傳的特征,因此推測淺色花或白色花是由于色素含量降低造成的。
表型多樣性是受遺傳多樣性和環境多樣性的共同影響的,既具有穩定性,又具有變異性[21]。本研究中只有‘紫波’ב麒麟’的子代出現少量復色系花色,且‘紫波’ב麒麟’組合的親本形態差異較大,推測復色現象是因為該組合2個親本的親緣關系較遠而產生的。趙蘊陽[22]研究發現遠緣雜交可能會引起轉座子激活,從而導致表觀遺傳不穩定性,進而引起基因表達和表型變異。劉青林等[23]、李軍等[24]、Morita et al.[25]認為轉座子激活引起基因不穩定表達會形成花色嵌合體。雖然復色性狀增加了觀賞植物的多樣性,但是其后代性狀也易發生改變[26-27]。本研究中出現的復色性狀能否穩定遺傳,且遺傳能力如何還不清楚,尚需要進一步研究。
花冠裂片內斑點是影響杜鵑花性狀的重要因子之一。本研究中雜交組合‘紫鶴’ב紅蘋果’親本花冠裂片內均含斑點,子代花冠裂片內均含斑點;雜交組合‘白鶴’ב粉紅泡泡’親本花冠裂片內均含斑點,子代出現少數不含斑點植株;雜交組合‘紫波’ב麒麟’父本花冠裂片內不含斑點,母本花冠裂片內含斑點,子代出現不含斑點植株。花斑的出現情況表明,花冠裂片內斑點具有顯性遺傳的特征,這與劉曉青等[16]研究結果一致。此外,百合[28]、三色堇(Violatricolor)[29]等花被上的花斑也具有顯性遺傳特點,所以推測斑點在杜鵑花遺傳中為顯性遺傳。在花冠類型上,3個組合雜交試驗的父本均為套筒,母本為單瓣或重瓣,子代的花冠類型出現了完全不同的分離比例,說明花冠類型的遺傳相對復雜,還需要增加正反交試驗和世代分離,分析其遺傳規律。
本研究通過對杜鵑花的花部和葉部的數量性狀進行相關性統計,發現多數花部性狀和葉部性狀呈正相關關系。其中,花徑和葉長的相關系數最高,為0.819,說明杜鵑花的營養生長和生殖生長具有較強的聯系性;葉長與葉寬、葉長寬比呈極顯著相關,相關性系數分別是0.751和0.749,說明葉長一定程度上決定了葉寬;花徑和花梗長度之間呈極顯著相關,說明花梗長度一定程度決定了花徑。所以在選擇育種過程中,可以利用性狀之間的相關性對決定性狀的基因進行宏觀篩選,通過較容易測量的葉長和花梗長度等性狀推測其他不宜測量的基因,并篩選出目標性狀,以提高育種效率[30-31],還可以利用變異系數高的性狀作為指示性狀對變異系數低且相關性強的性狀進行間接選擇,以提高育種效果[32]。此外,針對成熟期才表現出來的性狀也可以利用這種相關性進行預測[32]。如:杜鵑花實生苗的幼齡期較長,有鱗杜鵑從播種到開花需要3~5年,無鱗杜鵑則需要5~10年,甚至有些種類的杜鵑花需要十幾年[33]。可以通過分析葉片形態的遺傳變異規律和雜種優勢表現,利用葉部性狀和花部性狀的相關性起到早期鑒定和預先選擇的作用[34-35],達到篩選目標性狀,縮短杜鵑育種周期的效果。
