我國辣椒資源pvr2-eIF4E基因的多樣性分析
2021-05-26 13:09:18曹亞從顧曉振張正海于海龍馮錫剛張南南張偉麗景雅欣王海平李錫香BenoMOURY張寶璽王立浩
辣椒雜志 2021年1期
曹亞從 顧曉振 張正海 于海龍 馮錫剛 張南南 張偉麗景雅欣 王海平 李錫香 Beno?t MOURY 張寶璽 王立浩*
(1. 中國農業科學院蔬菜花卉研究所, 北京 100081; 2. 法國農業科學院植物病理研究所,阿維尼翁 84143)
辣椒(Capsicum spp.)屬于茄科辣椒屬,起源于南美洲。1493年,哥倫布首次將辣椒引入歐洲,并于15至16世紀后期引入東亞[1],由于復雜的地理環境以及人們的馴化等活動,形成了豐富的辣椒種質資源。遺傳多樣性使育種者可以利用種質資源中不同的等位基因來培育品種,等位基因挖掘已經被用來在已知基因座上發現未探索的等位基因多樣性,是利用資源多樣性育種的基礎。
我國是世界上辣椒種植面積最大的國家(http://www.FAO.org)。自2015年以來,辣椒已成為我國播種面積最大的蔬菜作物,年播種面積150萬 ~200萬hm2[2]。馬鈴薯Y病毒屬包括多個種,是一種世界性的病害。1956年Simons首次報道在美國加州有PVY侵染辣椒[3]。在中國,20世紀80年代馬鈴薯Y病毒屬對辣椒的危害就有廣泛報道[4-6]。近年來報道危害辣椒的馬鈴薯Y病毒屬病毒有馬鈴薯Y病毒(Potato Virus Y,PVY)、煙草蝕紋病毒(Tobacco Etch Virus,TEV)、辣椒斑駁病毒(Pepper Mottle Virus,PepMoV)、辣椒脈斑病(Pepper Veinal Mottle Virus,PVMV)等[7-14],嚴重影響辣椒的生產效益,尤其在高溫地區,是辣椒的重要病害,其危害僅次于CMV和TMV[15-16]。
eIF4E是植物對抗病毒的重要決定因子[17-20],酵母雙雜交及ELISA實驗表明病毒基因組連接蛋白(VPg)或其前體(NIa)可以與eIF4E或其同源異構體Eif(iso)4E相互作用,而互作缺失的病毒則喪失侵染性[21-22]。在辣椒中同樣發現了與馬鈴薯Y病毒屬病毒PVY和TEV抗性相關的eIF4E[23-25],根據發現的抗性及發現時間先后順序,Kyle和Palloix[26]將其命名為pvr2,后證明pvr1與pvr2為等位基因。pvr2-eIF4E序列多樣性豐富,感病等位基因為pvr2+,繼pvr2 1和pvr2 2被發現后[24,27-29],Charron等[30]對25份辣椒材料 (Capsicum annuumL.)的研究發現了pvr2的10個等位基因,在此10個等位基因中,有7個前人未報道,分別命名為pvr2 3-9。Ibiza等[31]采用EcoTILLING的方法,在233個辣椒栽培品種中發現了16個pvr2的等位基因,其中有13個等位基因前人未報道,分別命名為pvr2 10-22。pvr2-eIF4E序列的多樣性導致其功能差異,體現在對不同PVY小種(PVY0、PVY0,1、PVY0,1,2)[29]和TEV的抗性差異。
對pvr2-eIF4E的多態性研究發現,其外顯子的多態性比例高于內含子[30],因此pvr2-eIF4E的研究重點位于其外顯子。pvr2-eIF4E基因具有5個外顯子,研究表明外顯子1的多態性最豐富[30-31],因此本課題組前期對外顯子1的多態性進行了研究[32],在1 904份材料中共發現17個單倍型,14個有義突變位點。
我國有著豐富的辣椒種質資源,但是pvr2-eIF4E作為辣椒抗PVY和TEV育種工作中有效而穩定的抗源,并未在豐富的資源中得到廣泛而深入的研究。本研究通過全面鑒定pvr2-eIF4E的所有外顯子,挖掘出我國辣椒資源中的PVY和TEV抗性材料,為PVY和TEV抗性材料的挖掘和利用提供研究基礎。
1 材料與方法
1.1 材料
我國種質資源中期庫1 863份辣椒材料,其中187份材料引種自國外,引種地區包括美洲、歐洲、非洲、亞洲的26個國家;1 676份材料來自于中國33個省、市、自治區。
1.2 基因組DNA提取
辣椒材料播種于穴盤,待長到約6片真葉時,取嫩葉片,采用改良CTAB方法提取葉片基因組DNA[33]。采用Biospec-nano顯微分光光度計(Shimadzu Corporation)測定DNA的濃度及質量。基因組DNA稀釋到25 ng/L備用。
1.3 擴增引物設計
通過美國國家生物技術信息中心NCBI網站(http://www.ncbi.nlm.nih.gov/)公布的Yolo Wonder的pvr2-eIF4E的編碼序列確定目的基因序列(AY122052.1)。將序列比對到CM334 v1.55版 本 基 因 組[34](http://passport.pepper.snu.ac.kr/?t=PGENOME),得到pvr2-eIF4E在CM334基因組中的基因為CA04g00860。
利用在線工具Primer 3(http://primer3.ut.ee/)設計引物擴增外顯子,擴增引物見表1。
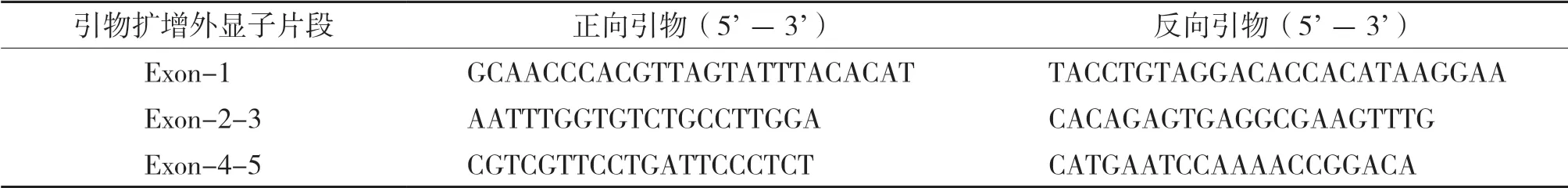
表1 pvr2-eIF4E外顯子測序引物序列Table 1 Sequences of pvr2-eIF4E exon sequencing primers
1.4 產物擴增及測序
以葉片基因組DNA為模板,分別對外顯子1、外顯子2-3和外顯子4-5進行擴增,采用北京全式金生物技術有限公司的DNA高保真聚合酶試劑盒(TransStart ? Top taq DNA Polymerase),擴增體系為30 μL:2 μL 基因組DNA,0.6 μL高保真聚合酶,3 μL 緩沖液(10 X),2.4 μL dNTP,上下游引物各1.2 μL,19.6 μL ddH2O。PCR反應程序為:94 ℃ 3min;94 ℃ 30 S,58 ℃ 30 S,72 ℃ 60 S,35 個循環;72 ℃ 10 min。PCR產物送上海生工生物工程有限公司進行Sanger測序,測序引物采用擴增正向引物。對含有新發現等位基因的材料進行3次重復測序。
1.5 測序結果分析
將測序結果在Cexpress軟件中打開,序列對齊后根據Yolo Wonder的編碼序列,確定各材料的堿基突變位點,用MEGA5.0對測序序列進行多重比對,統計各材料間氨基酸差異位點。
1.6 PVY抗性接種鑒定
對含有新發現等位基因的材料進行PVY抗性接種鑒定。鑒定工作委托法國農業科學院完成,接種PVY病毒株系為LYE84.2和SON41,其中LYE84.2為PVY0,SON41為PVY0,1,2。
1.7 PVY抗性材料遺傳關系分析及核心種質構建
利用Gu等[35]SSR數據,提取含有抗性等位基因材料的數據,將其轉化為矩陣,通過DARWin軟件(https://darwin.cirad.fr/)對含有抗性等位基因的材料構建進化樹。
根據進化樹將材料進行分組,對每種pvr2等位基因在每個組中提取遺傳距離最遠的兩份材料,分組中少于2份的,則提取實際材料數。
2 結果與分析
2.1 pvr2-eIF4E基因序列多樣性分析
在PCR擴增測序的1 863份辣椒材料中,發現的變異均為SNP,共檢測到10個有義氨基酸突變位點(表2),共檢測到10種等位基因,其中有1 117份材料與感病材料Yolo Wonder有相同的氨基酸序列,為pvr2+;有8種等位基因與前人已報道等位基 因pvr2 1、pvr2 2、pvr2 3、pvr2 4、pvr2 6、pvr2 7、pvr2 9、pvr1 +編碼序列一致,共744份材料;有1份材料為新發現的等位基因,命名為pvr223,在第1外顯子的第185位堿基發生突變,導致編碼苯丙氨酸的密碼子(UGG)突變為終止密碼子(UAG)(圖1)。以感病材料Yolo Wonder編碼區序列為對照,1 863份材料中共有746份發生有義氨基酸突變,約占總數的41%。
突變位點的統計結果顯示(表2),突變位點主要位于第1外顯子,有7個位點發生有義突變,在第2、3、4外顯子分別有1個有義突變,在第5外顯子未發現有義突變。突變位點最多的位點位于第1外顯子的第67位氨基酸,有644份材料發生有義突變;其次為第205、71、79位氨基酸,分別有232、97、20份材料發生有義突變,其余位點發生突變的材料很少。
2.2 pvr2-eIF4E等位基因抗性分析
在1 863份辣椒材料發現的10種pvr2-eIF4E等位基因中,已知有2種等位基因為感病,7種等位基因對PVY和TEV部分小種具有抗性(表2),1種等位基因的抗性未知。
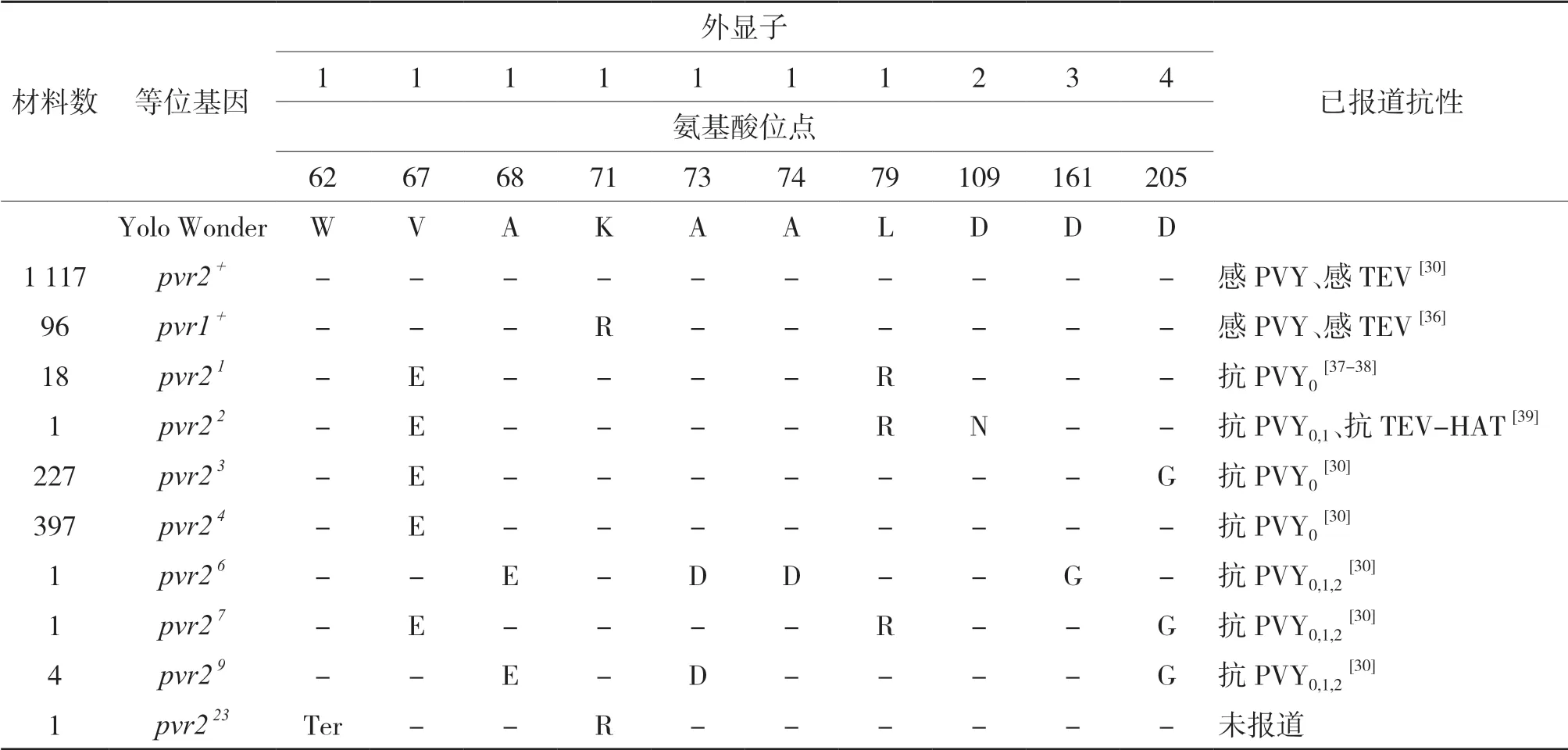
表2 pvr2-eIF4E基因多樣性分析Table 2 Analysis of pvr2-eIF4E gene diversity
采用PVY株系LYE84.2(PVY0)和SON41(PVY0,1,2),對含有新發現等位基因pvr223的辣椒材料進行PVY抗性接種鑒定。結果顯示,含有pvr223的辣椒材料易感此2種PVY株系(表3)。

表3 人工接種鑒定pvr223等位基因材料對PVY抗性的統計數據Table 3 Summary on the resistances of materials with pvr223 allele to PVY measured by artificial inoculation
在研究的1 863份辣椒材料中,共有649份材料含有pvr2抗性等位基因,占總材料數的34.84%。649份材料均對PVY0具有抗性,占總材料數的34.84%;其中有6份材料含有高抗PVY等位基因(pvr2 6、pvr2 7、pvr2 9),對PVY0,1,2具有抗性,占總材料數的0.32%;有1份材料含有已報道抗TEV的等位基因pvr2 2。
2.3 含有抗PVY的pvr2-eIF4E等位基因材料地理分布
抗性材料主要來源于我國國內,有586份,占抗性材料的90.29%;有63份抗性材料來源于國外,占抗性材料的9.71%(圖2)。國內的抗性材料主要來源于湖北,有134份,占國內抗性材料的22.87%,其次來自于湖南、云南和四川,分別有52、49和42份,占國內抗性材料的8.87%、8.36%和7.17%。收集自國外的抗性材料主要來源于美國,有25份,占國外抗性材料的39.68%,其次來自于泰國、印度和荷蘭,分別有9、5和4份,占國外抗性材料的14.29%、7.94%和6.35%。
含有高抗PVY等位基因(pvr2 6、pvr2 7、pvr2 9)的材料分別來自于中國山西(1份)、美國(2份)、墨西哥(1份)、西班牙(1份),另有1份引自國外,引進地不詳。含有已報道抗TEV的pvr22材料來自英國。

圖2 含有抗PVY等位基因材料的地理分布圖 A,在不同國家之間的分布;B,在中國各地的分布Figure 2 Geographical distribution of materials containing PVY-resistant alleles. A, distribution among diあerent countries;B, distribution throughout China.
2.4 含有抗PVY的pvr2-eIF4E等位基因材料遺傳關系分析
基于SSR標記的分型數據,對649份辣椒抗性材料的遺傳關系進行了分析(圖3)。根據遺傳關系可以將含有pvr2抗性等位基因的材料分為3個群,如圖3所示,其中群I包含本研究中鑒定到的所有抗性等位基因,又分為3個亞群;群II包含4種抗性等位基因(pvr2 1、pvr2 3、pvr2 4、pvr2 9),又分為3個亞群;群III包含3種抗性等位基因(pvr2 1、pvr2 3、pvr2 4),又分為4個亞群。
2.5 PVY抗性核心種質構建
結合SSR標記分型數據構建的遺傳關系和pvr2等位基因,抽取PVY抗性核心種質,每個亞群中抽取每個pvr2等位基因2份,抽取在進化樹上遺傳距離最遠的2份,少于2份的則抽取實際數據材料。PVY抗性核心種質材料共抽取53份,具體見表4。
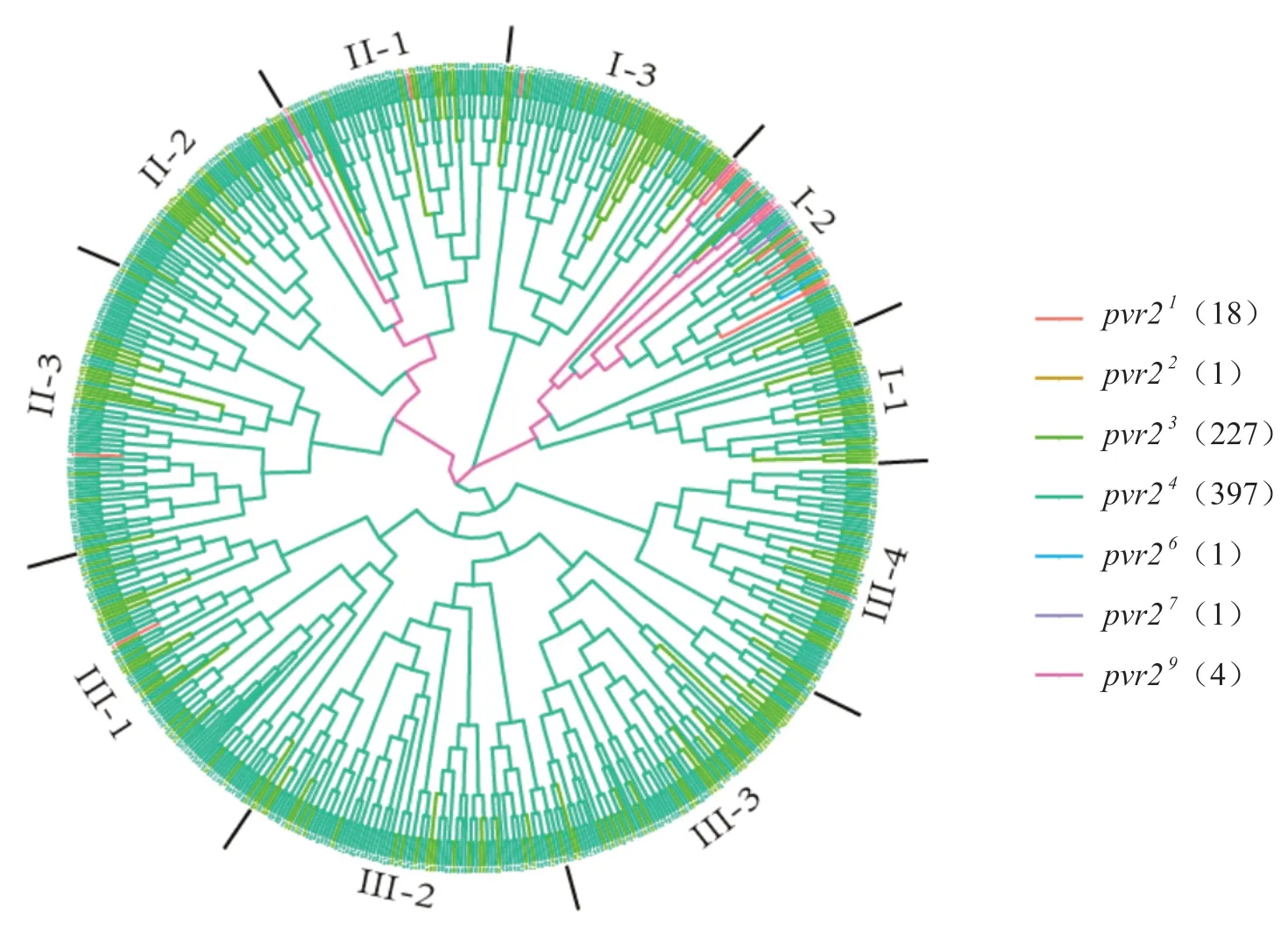
圖3 含有抗性pvr2等位基因材料的遺傳關系圖Fig. 3 Genetic relationship of accessions containing resistant pvr2 alleles
3 討論
pvr2-eIF4E基因產生抗性的確切機制仍有待闡明,對pvr2基因表達數據的分析表明,植物感病與否的表型差異是由pvr2氨基酸變化決定的,而不是因為其他調節eIF4E表達的因子決定[40]。已有報道的蛋白互作研究認為,pvr2-eIF4E基因發揮抗性作用,是由于病毒Vpg蛋白和植物eIF4E蛋白的互作被破壞,病毒Vpg不能結合到eIF4E,從而影響病毒的復制、移動等[36,40]。通過結構模型預測,認為pvr2的突變位點位于蛋白的表面,第62~79位氨基酸被定義為蛋白結構域I,第106~109位氨基酸被定義為蛋白結構域II,這兩個區域可能是pvr2-eIF4E與PVY和TEV互作的關鍵區域,參與對PVY的抗性[41-42]。本研究發現的新等位基因pvr223的突變發生在第1外顯子的第62位氨基酸,由苯丙氨酸突變為終止密碼子,使原本含228個氨基酸的蛋白突變為僅有62位氨基酸的蛋白。雖然新發現等位基因pvr223僅保留了很短的蛋白序列,保留的序列不存在前人報道的與Vpg互作的候選關鍵區域,但仍然感病,因此pvr2-eIF4E可能存在其他抗病相關作用位點及作用機制,或含有pvr2 23-eIF4E的材料存在其他基因編碼與PVY病毒Vpg蛋白互作的蛋白。
辣椒傳入我國較晚,遺傳背景比較狹窄,本研究鑒定的辣椒資源多收集于“七五”、“八五”期間,多數未利用多抗性材料經過抗性轉育,因此,雖然我國有大量含PVY抗性等位基因的材料,但是多樣性不夠豐富,多抗性差:含有抗多個PVY小種的基因型等位基因pvr2 6、pvr2 7、pvr2 9材料僅有6份,且有5份是引種自國外;含有已報道抗TEV基因型等位基因pvr2 2的材料僅有1份。近年來,各地多有報道PVY和TEV病毒在辣椒上引起的危害[7-9,11,13,15,43-44],我國現存種質資源中缺少PVY和TEV高抗材料,需要引起重視,今后要加強引入豐富的pvr2抗性等位基因材料,用以培育豐富類型的多抗PVY和TEV辣椒品種。
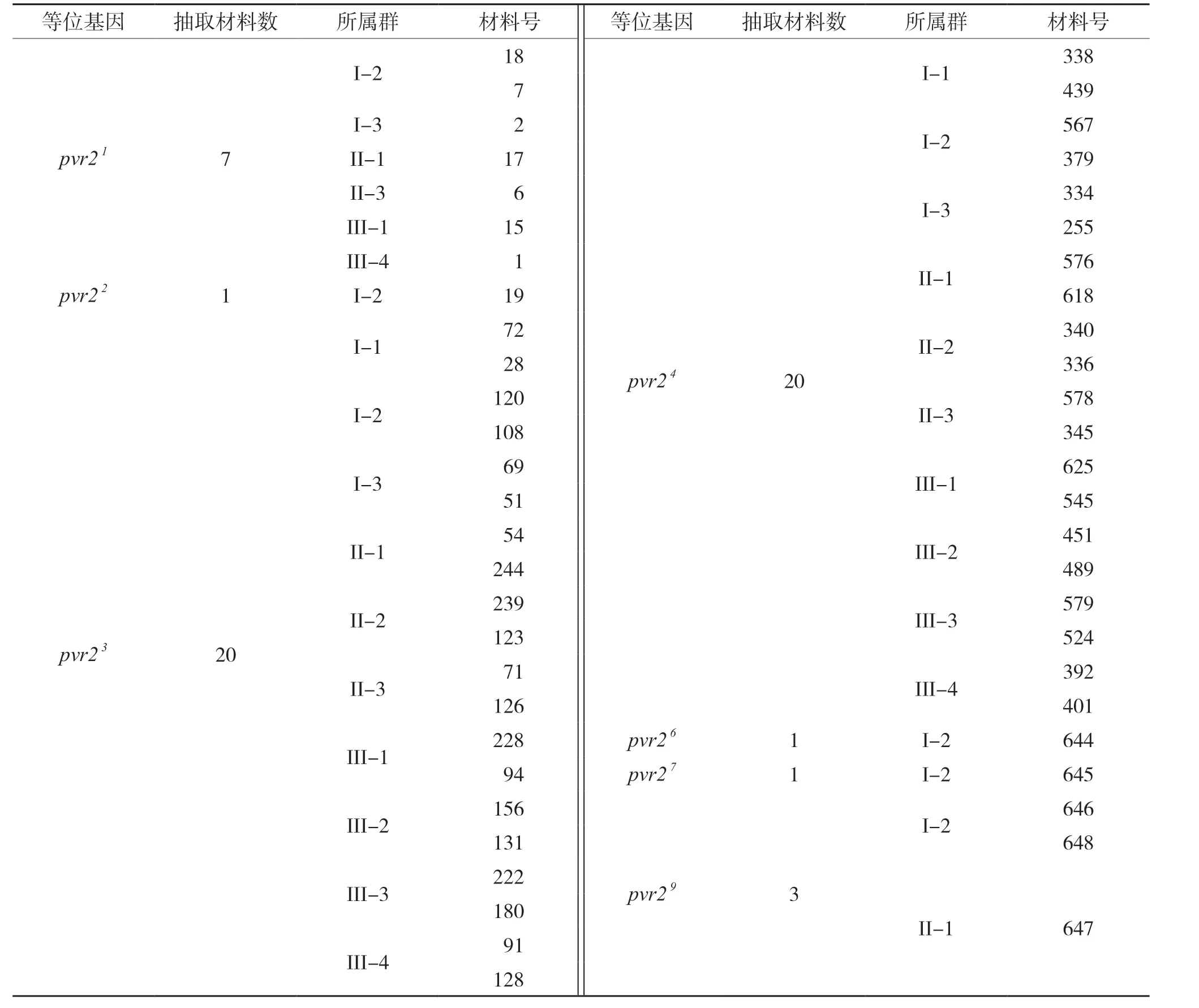
表4 PVY抗性核心種質材料抽取情況Table 4 Extraction of PVY resistant core germplasm