MCL1激活MAPK-MEK信號通路促進食管癌細胞EC9706增殖
2021-05-26 02:24:44崔艷麗何利珍
醫學理論與實踐 2021年10期
關鍵詞:檢測
崔艷麗 何利珍
河南省焦作市第二人民醫院檢驗科 454000
食管癌是一種常發的嚴重腫瘤,引起食管癌的因素較多,以不良飲食習慣、經濟社會地位和常吃粗糧等相關性比較高[1]。臨床上治療食管癌多以手術、放化療為主,且發生率和死亡率在國內均較高。為了提高癌癥患者的生存率和存活時間,逐漸開始嘗試一些分子靶向療法,但在食管癌中還沒有成熟的分子靶點,需要進一步揭示調控食管癌增殖的分子機制,找到好的分子治療靶點。
髓樣細胞白血病因子1(Myeloid cell leukemia-1,MCL1)最初被發現是細胞分化相關的一個基因,在白細胞分化過程中被發現其能夠誘導單核/巨噬細胞分化途徑。后來發現MCL1是一種腫瘤發生相關的蛋白,在多種消化道腫瘤中高表達,但是其在食管癌中表達情況及是否影響食管癌細胞增殖還不確定。本項目基于對臨床病例材料進行檢測,發現MCL1在食管癌組織中高表達,并通過構建MCL1的真核表達載體,探討其對食管癌細胞增殖的影響。
1 材料與方法
1.1 組織樣品收集 食管癌樣本為2018年在本院手術切除癌組織樣本(男9例,女3例,平均年齡68.3歲)。對照樣本為對應的癌周組織。將癌組織樣品及癌周組織碾磨,以Triol法提取RNA,然后用實時相對定量法(qRT-PCR)檢測食管癌組織中MCL1基因轉錄情況,所用定量引物序列為:Q-MCL1-F:5’-GAC GAG TTG TAC CGG CA GTC-3’,Q-MCL1-R:5’-TTT GTT ACG CCG TCG CTGA-3’。產物大小為266bp。內參引物序列為:Q-β-actin-F:5’-GAG AAA ATC TGG CAC CAC ACC-3’,Q-β-actin-R:5’-GGA TAG CAC AGC CTG GAT AG CAA-3’。產物大小為177bp。
1.2 細胞培養 食管癌細胞EC9706在37℃恒溫二氧化碳培養箱中培養,培養液為添加10%胎牛血清和1%青鏈霉素的RPMI1640培養基。其中,RPMI1640基礎培養基購自Hyclone公司,胎牛血清購自依科賽生物科技有限公司,青鏈霉素購自碧云天生物技術有限公司。
1.3 載體構建 實驗中用到的pCDNA3.1/myc-His載體購自Addgene公司,基因擴增自EC9706細胞。實驗中所用的克隆引物為:MCL1-F: CGG GAT TCA TGT TTG GCC TCA AAA GAA ACG CGG,MCL1-R: GGA ATT CTC TTA TTA GAT ATG CCA AA CCA。產物大小為:1 068bp(含酶切位點及保護堿基)。引物由生工上海生物工程(上海)股份有限公司合成。構建的載體命名為:pCDNA3.1-MCL1-myc-His。
1.4 轉染試驗 EC9706細胞提前接種于12孔細胞培養板,次日細胞生長至80%鋪滿皿底時,實驗組每孔轉染pCDNA3.1-MCL1-myc-His質粒3μg,對照組轉染pCDNA3.1/myc-His空質粒3μg。轉染步驟按照Thermo Turbofect轉染試劑說明書進行,在轉染24h和48h后進行收樣檢測。
1.5 Western blot檢測 上述1.4中細胞樣品,棄去培養基,用添加蛋白酶抑制劑的RIPA裂解液100μl在冰上裂解細胞15min,然后添加6×loading buffer, 蛋白樣品95℃煮樣10min后備用。聚丙烯酰胺凝膠電泳(100V,1.5h)分散蛋白后,將膠上蛋白轉膜(250mA,100min)轉移至PVDF膜上,脫脂奶粉封閉2h后,以抗磷酸化MEK1(p-MEK1)抗體(1∶1 000,購自Abclone公司)、以抗MEK1抗體(1∶1 000,購自Abclone公司)、β-actin抗體(1∶5 000,購自Abclone)為一抗分別孵育PVDF膜4℃過夜;PBST溶液清洗PVDF膜3次后;以HRP標記的羊抗鼠/兔二抗(1∶5 000,購自Abclone公司)室溫孵育1h;然后用PBST溶液清洗PVDF膜3次;ECL化學發光液(購自Millipore公司)顯色,化學發光儀拍照。
1.6 細胞計數 將細胞接種至12孔板,待細胞生長至70%左右時轉染MCL1真核表達載體和對照的空載體,分別在轉染后0、24和48h,用胰酶消化細胞,以血球計數板計數,每組3個樣品,每個樣品重復計數3次。
1.7 細胞劃痕試驗 細胞生長至80%左右時轉染超表達MCL1的質粒,并同期轉染空質粒作為對照。在次日細胞長滿皿底時,以無菌移液器吸頭在細胞皿內劃線,分別在劃線后0、24和48h后拍照,檢測細胞遷移能力變化情況。
1.8 細胞周期檢測 上述1.4中細胞樣品,以PBS清洗細胞后,胰酶消化后再次以PBS清洗細胞1遍,離心棄去PBS,以預冷的70%乙醇在4℃固定細胞48h,然后離心棄去乙醇,以PBS懸浮細胞,加入細胞周期用PI溶液,在37℃孵育30min,流式細胞儀檢測細胞周期。
1.9 qRT-PCR檢測 上述1.4中細胞樣品,棄去培養基并以PBS清洗1遍后,每孔加入1ml Trizol試劑(購自Takara公司),提取細胞總RNA,然后以反轉錄試劑盒(購自天根生化科技有限公司)反轉錄獲得cDNA,再以熒光定量試劑盒(天根生化科技北京有限公司)檢測相關下游基因表達情況。Q-C-MYC-F:5’-CCG ACC AGC TGG AGA TGG TGA-3’,Q-C-MYC-R:5’-AGC CTG GTA GGA GGC CAG CTT-3’;Q-C-FOS-F:5’-CTA TGC AGC AGA CTG GG AGC-3’,Q-C-FOS-R:5’-AGA CGT GTA AGC AGT GC AGC-3’;Q-CCND1-F:5’-AGC TGT GCA TCT ACA CC GAC-3’,Q-CCND1-R:5’-GAA ATC GTG CGG GGT CA TTG-3’。
1.10 統計學方法 采用 Graphpad Prism5 統計軟件進行分析。數據以均數±標準差表示,組間比較采用獨立樣本t檢驗;檢驗水準α=0.05。
2 結果
2.1 MCL1在食管癌組織中高表達 2018年1—12月在本院收集的12例手術切除的食管癌組織(Cancer tissue) 樣品及其癌周組織(Control)樣品,通過qRT-PCR檢測,將對應的數據等比例換算后,發現在其中10份臨床樣品中,MCL1在食管癌組織中表達量高于對應的癌周組織,僅2例差異不顯著,如圖1所示。
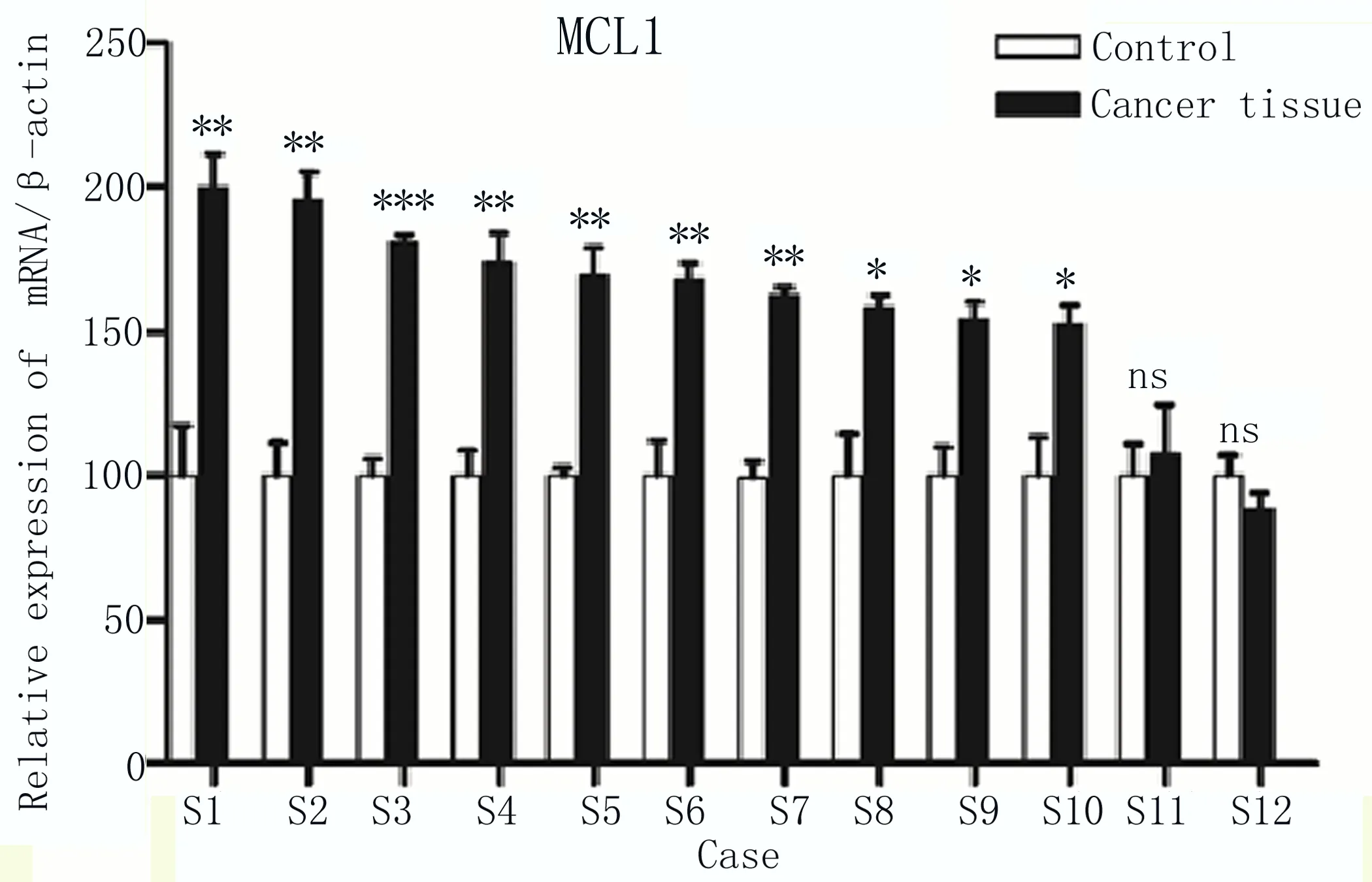
圖1 相對定量PCR檢測12例癌組織(Cancer tissue)和癌周組織(Control)中MCL1表達量 兩組比較,*P≤0.05;**P≤0.01;***P≤0.001
2.2 構建表達MCL1的真核表達載體 通過PCR的方法,用保真酶將MCL1基因擴增出來(圖2A);雙酶切后裝至pCDNA3.1/myc-His載體上,酶切驗證構建的目的載體pCDNA3.1-MCL1-myc-His,結果顯示獲得和預期大小的條帶(圖2B),測序后序列無突變。這些結果提示載體pCDNA3.1-MCL1-myc-His構建成功。
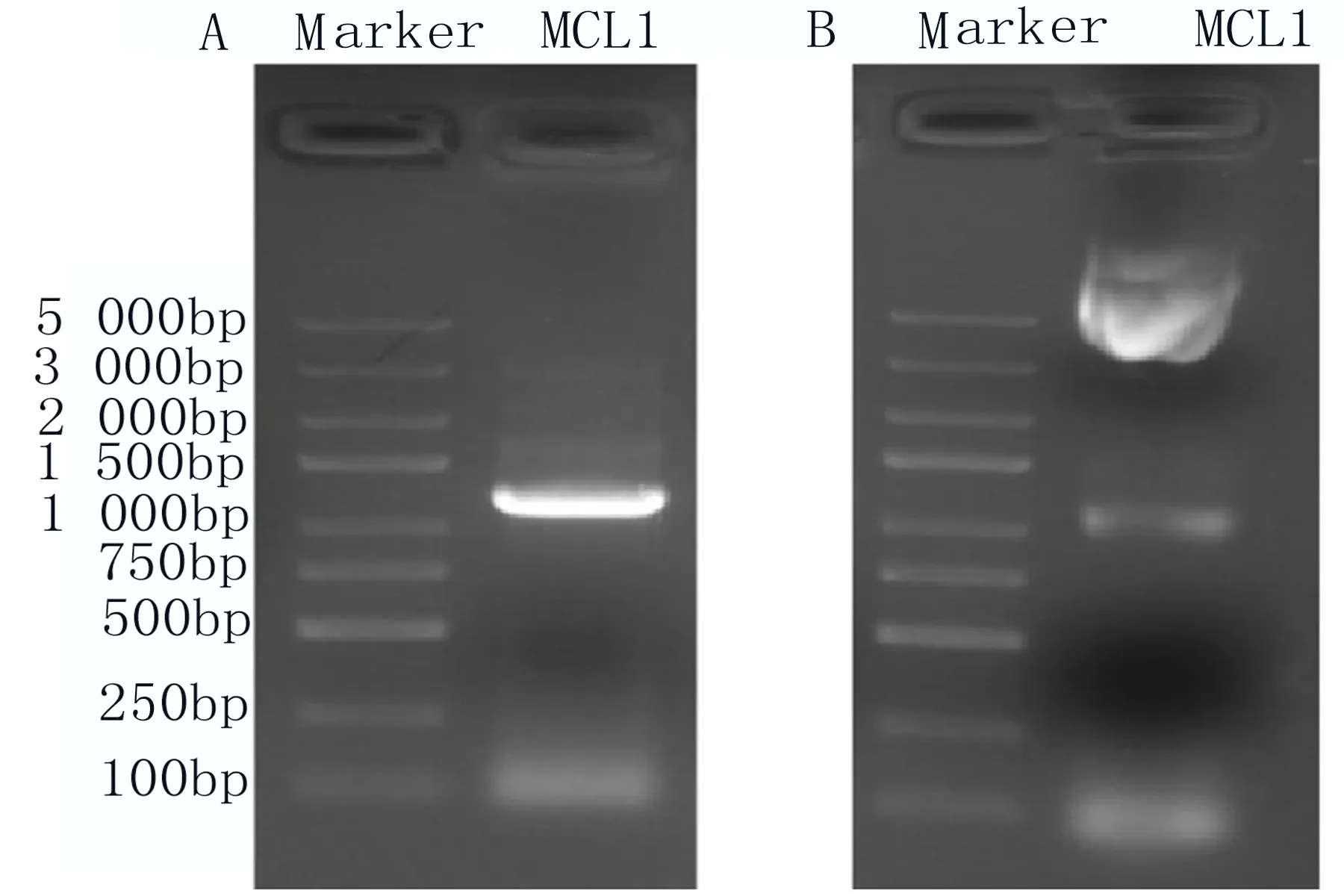
圖2 核酸電泳結果
2.3 細胞計數檢測超表達MCL1促進EC9706細胞增殖 將構建好的MCL1真核表達載體pCDNA3.1-MCL1-myc-His轉染至EC9706細胞內,同時以轉染pCDNA3.1/myc-His空質粒為對照組,在轉染后24和48h分別檢測兩組的細胞數量,結果顯示超表達MCL1組的細胞數量高于對照組,其中在轉染48h,差異顯著,如圖3所示。
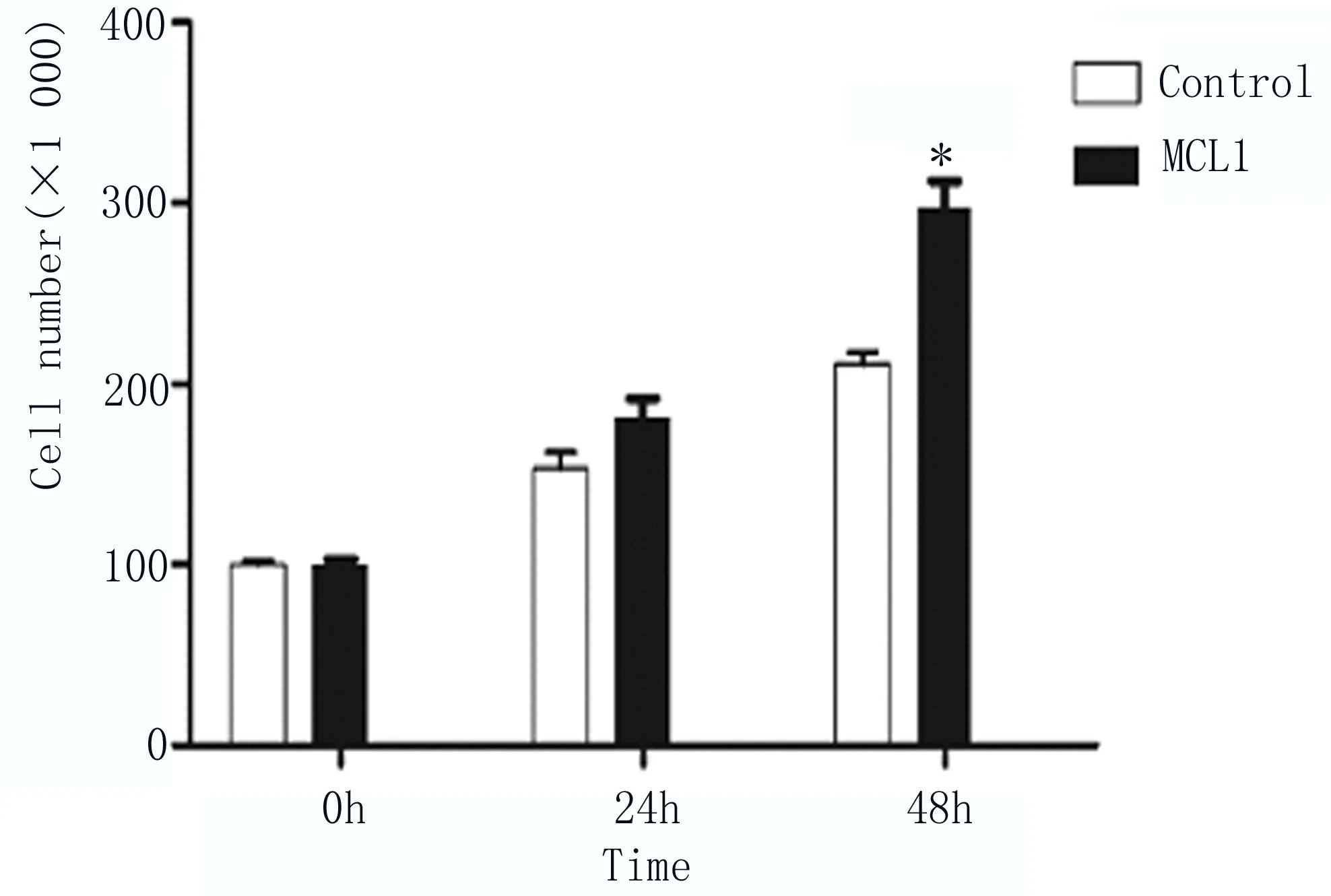
圖3 細胞計數檢測不同時間點細胞數量差異 兩組比較,*P≤0.01
2.4 細胞周期檢測超表達MCL1促進EC9706細胞增殖 以流式細胞儀檢測細胞周期,結果顯示,在轉染24h后對照組(Control)和超表達MCL1組(MCL1)的G1期比例分別為(72.44±0.56)%、(70.05±1.05)%(P≤0.05);在轉染質粒48h后G1期細胞比例分別為(73.95±0.05)%、(71.55±0.55)%(P≤0.01);對應的S+G2期細胞比例在超表達MCL1后高于超表達空質粒的對照組。以上結果提示超表達MCL1能夠增加S+G2期細胞比例,提高了細胞增殖能力,如圖4所示。
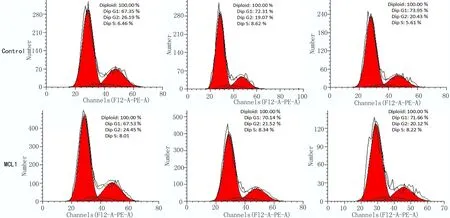
圖4 流式檢測超表達MCL1后細胞周期變化
2.5 超表達MCL1促進EC9706細胞遷移 細胞轉染MCL1后,次日細胞將近鋪滿皿底,以移液器吸頭劃線,并將培養基換成無血清培養基,在劃線后0、24和48h拍照。結果顯示,超表達MCL1的細胞顯示更好的遷徙能力,尤其在48h時差異較顯著,如圖5所示。
2.6 抑制MAPK信號通路能夠抑制EC9706細胞增殖 用抑制劑U0126(100nM)處理細胞,Western blot檢測MEK含量降低,磷酸化MEK含量增加(圖6);細胞計數檢測發現用U0126抑制劑處理細胞24和48h后,細胞數量較僅加溶劑的對照組(Control)明顯減少,如圖7所示。以上結果顯示抑制MAPK-MEK信號通路能夠抑制EC9706細胞增殖。
2.7 超表達MCL1激活MEK信號通路 在細胞內轉染pCDNA3.1-MCL1-myc-His來超表達MCL1蛋白,以轉染空載體pCDNA3.1/myc-His的細胞為對照組(Control),Western blot檢測發現MCL1能夠減少MEK含量,增加p-MEK含量(圖8);qRT-PCR檢測MEK下游靶基因C-MYC、C-FOS和CCND1,結果顯示,EC9706細胞中超表達MCL1后能夠激活MEK下游靶基因的轉錄, 差異顯著有統計學意義,如圖9所示。

圖5 細胞劃痕實驗檢測超表達MCL1后對細胞遷移的影響
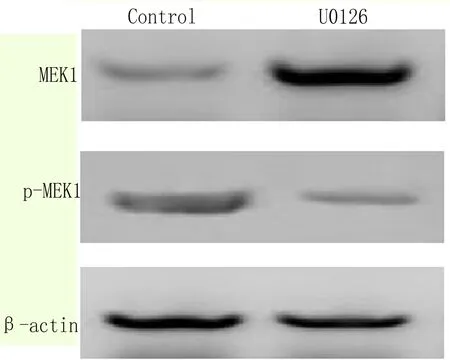
圖6 Western blot 檢測U0126處理細胞后細胞內MEK1含量變化
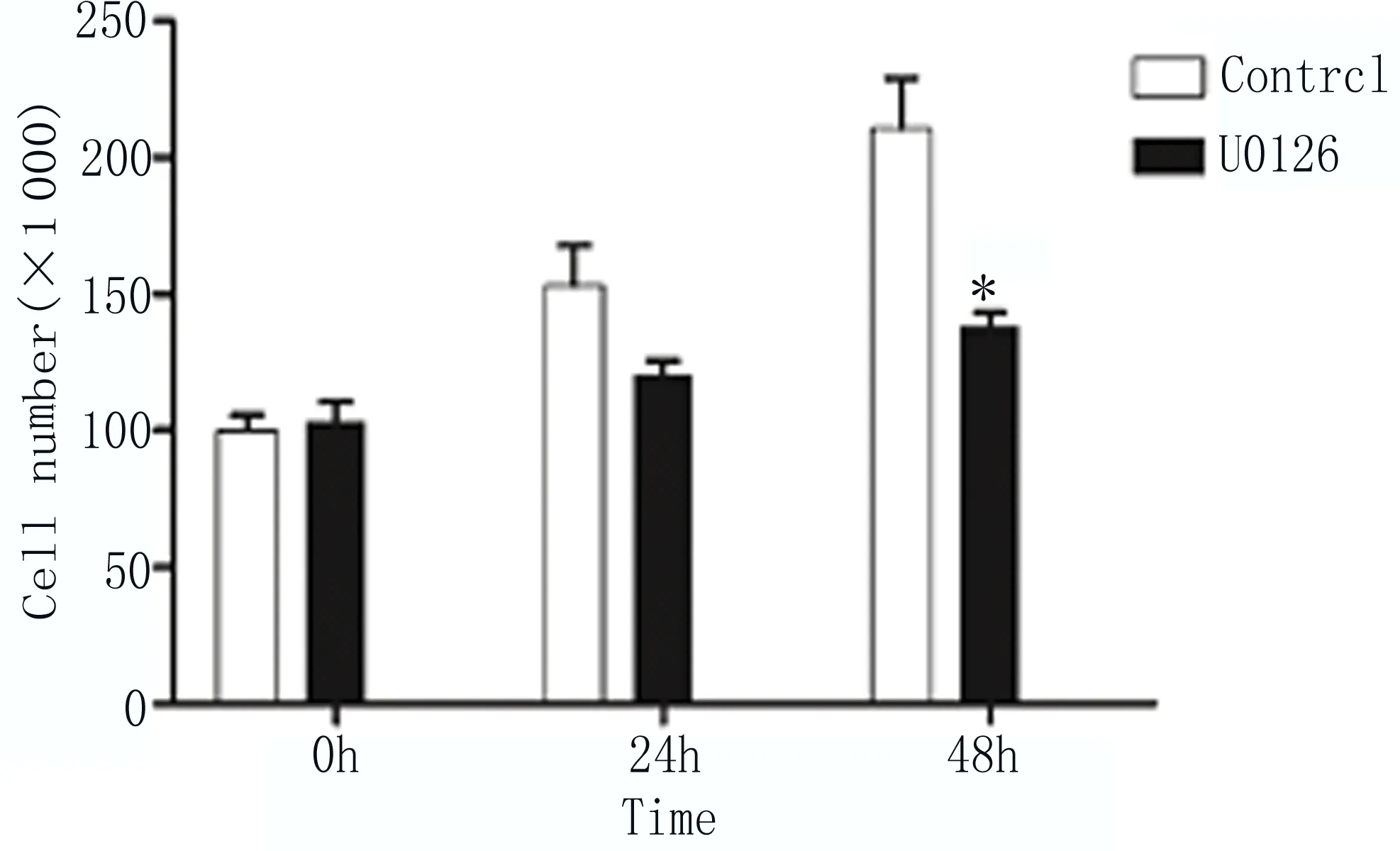
圖7 細胞計數檢測U0126處理細胞后細胞增殖情況 兩組比較,*P≤0.05
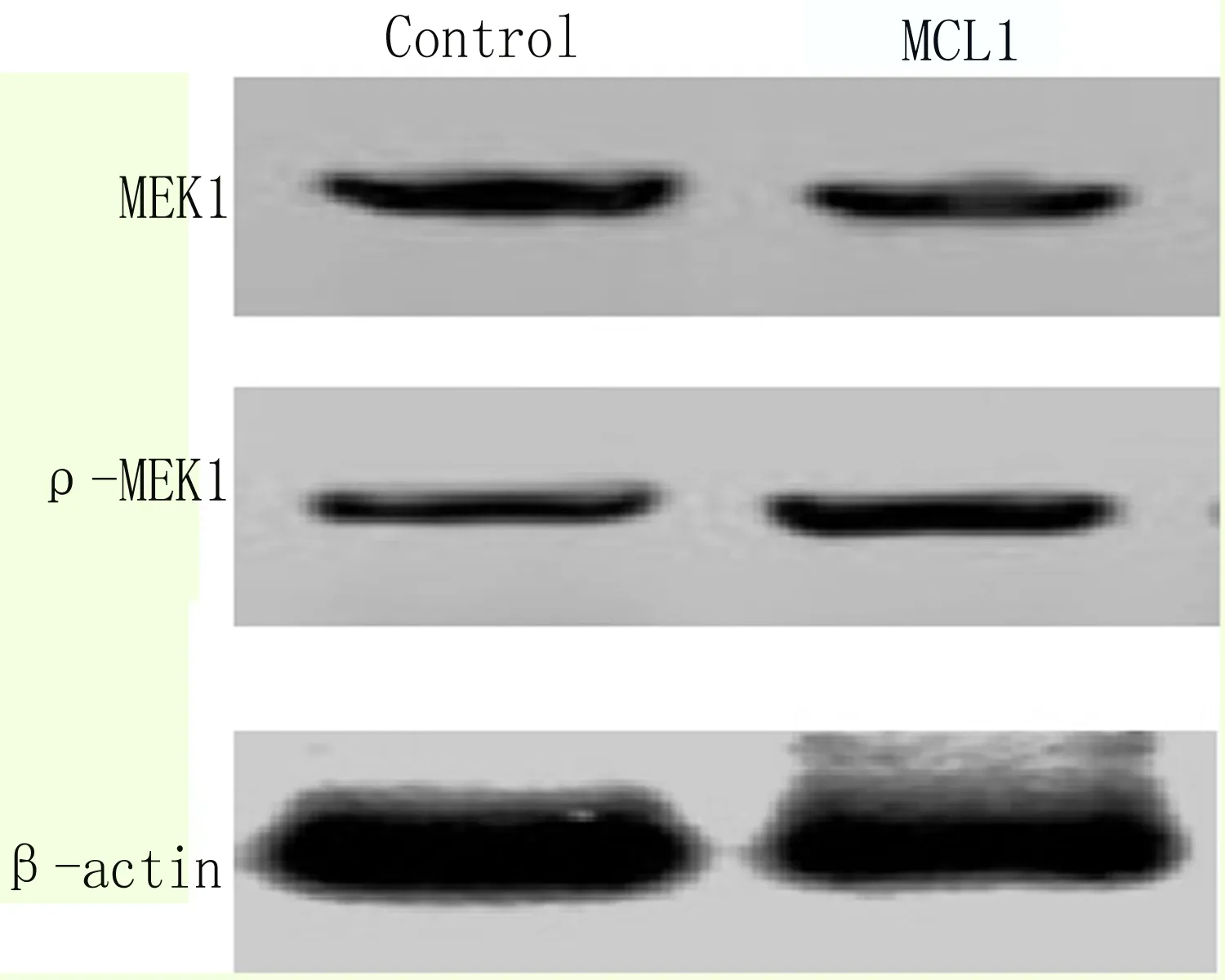
圖8 Western blot 檢測超表達MCL1后細胞內MEK含量變化
3 討論
MCL1能夠和Bcl2互作,在抗細胞凋亡方面發揮作用,癌細胞通常由正常細胞轉化產生,因此,抗凋亡和調控細胞分化的BCL1基因可能和癌細胞的生成存在相關性。除了在骨髓瘤中檢測其和癌癥的預后相關外,在前列腺癌[2]、胃癌[3]等均和癌癥的發生存在相關性。但是,MCL1是否可以作為食管癌的標記基因,在食管癌中的表達規律還不清楚。本研究通過對近期臨床上收集的12份食管癌組織樣品進行檢測,發現大部分癌組織中均高表達MCL1基因。因此,筆者推測在食管癌發生中,MCL1也可能發揮重要作用。

圖9 相對定量PCR檢測超表達MCL1后EC9706細胞內 C-MYC、C-FOS和CCND1基因表達量變化 兩組比較,*P≤0.05;**P≤0.01
癌細胞和正常細胞的一個差異是癌細胞能夠持續增殖,前期研究表明,MCL1是維持T細胞存活重要的基因[4]。在前列腺癌細胞中,MCL1是一個治療靶點,抑制MCL1的表達能夠誘導前列腺癌細胞增殖[5]。這些結果表明MCL1在調控癌細胞增殖時發揮重要作用,為檢測在食管癌中過表達MCL1是否影響細胞增殖和遷移,實驗中通過構建表達MCL1的真核表達載體,驗證了超表達的MCL1能夠促進EC9706細胞增殖并促進其遷移。這些結果提示,MCL1對食管癌細胞EC9706的增殖也很重要。
能夠調控EC9706細胞增殖的因素較多,一些中藥有效成分如六君子湯,乙酸乙酯部位可以調控IL-6/STAT3信號通路進而調控EC9706增殖[6];壁虎活性肽[7]和兗州卷柏總黃酮[8]能夠通過調節其線粒體凋亡通路相關蛋白促進EC9706細胞凋亡抑制其增殖。這些結果表明,有多種藥物可以調控EC9706細胞增殖。而MCL1調控細胞增殖的機制研究也比較多,有研究顯示,在黑色素瘤細胞中,同時靶向MCL1和ERK1/2信號通路能夠改善患者的預后[9];在膠質瘤細胞中,沉默MCL1基因后能夠導致PI3K/AKT信號通路的抑制,進而促進膠質瘤細胞的凋亡[10]。但是,對于MCL1調控EC9706細胞增殖的機制還不清楚。有研究顯示靶向MCL1和MEK的抑制劑共同使用,利于非小細胞肺癌[11]、急性髓系白血病細胞[12]和胰腺癌細胞[13]凋亡,這些結果提示,MCL1可能和MEK共同發揮作用對癌細胞抑制明顯,推測二者起協同互補作用,但是MCL1和MEK是否存在相互調節還沒有研究。本實驗中在EC9706細胞中超表達MCL1,檢測到下游MEK蛋白磷酸化增加,這些結果提示MCL1不僅和MEK是兩個協同互補的靶點,相互之間還存在調控關系。
MEK信號通路激活,除了本身磷酸化水平的增加,其下游靶分子也能被激活。MEK下游靶分子有C-FOS、cyclinD1和C-MYC等。C-FOS、C-MYC和 cyclinD1等是處于MEK下游調控細胞增殖的基因[14],實驗中也發現這些MEK下游基因表達量提高,進一步證明MCL1能夠激活MEK信號通路。
總之,實驗證明MCL1促進細胞增殖能夠通過靶向MAPK-MEK信號通路發揮作用,為MCL1調控癌細胞增殖機制提供新的理論依據。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
