水稻響應冷脅迫基因OsHI-XIP過量表達載體構建及遺傳轉化體系建立
2021-05-24 12:06:40熊文濤沈雨民陳明亮羅世友熊煥金吳小燕肖葉青
江西農業學報 2021年5期
熊文濤,沈雨民,陳明亮,羅世友,熊煥金,吳小燕,肖葉青
(江西省農業科學院 水稻研究所/水稻國家工程實驗室(南昌)/國家水稻改良中心 南昌分中心,江西 南昌 330200)
東鄉野生稻(以下簡稱“東野”)是迄今為止發現的分布緯度最北的普通野生稻,其蘊藏了其他普通野生稻和栽培稻所不具備的強耐冷性[1-2]。近年來,研究人員陸續從東野后代群體中篩選到多個綜合農藝性狀優良的耐冷株系[3-4]。其中,從東野和粳稻0298的雜交后代篩選的耐冷粳稻東野1號的稻蔸能成功越冬,該品種已通過江西省農作物新品種審定[5]。目前,相關研究報道了東野的3個耐冷基因,以及低溫響應調控因子ICE1的同源基因OrbHLH1[6];通過QTL定位區段找到了候選基因LTT7[7]和Cold1[8],其中Cold1起源于中國普通野生稻,經過人工馴化及篩選,現普遍存在于東北亞的粳稻品種中[8]。
贛香A是本課題組選用1504(贛早秈49號)、IR58025B、金23B、新露B和江農早2號B等5個具不同優良特性的保持系做親本,采用雜交、復交和系統選擇等方法選育出來的新品種[9]。該品種的綜合農藝性狀優良,如落色好、不早衰、千粒重較大、有香味等,而且對稻瘟病具有很強的抗性。
為了能夠將東野中的耐冷基因導入贛香B中,從而提高贛香B的耐冷性,我們在前期的研究中以贛香B為受體親本,以東野為供體親本,進行回交,利用分子標記進行輔助選擇,從回交后代中選擇性狀優良的單株進行耐冷性鑒定,創制了攜帶耐冷基因的贛香B品種[10]。之后對贛香B和耐冷性較好的株系IL742在常溫和低溫下的表型進行測序及表達譜差異分析,找到了東野中與冷脅迫相關的6個基因[11]。為了進一步探究這些冷脅迫相關基因的分子機理,本研究構建了這些基因的過量表達載體,并利用農桿菌介導法獲得了轉基因陽性植株,可為進一步研究這些基因的分子機理提供可靠的材料。
1 材料與方法
1.1 試驗材料
轉化受體材料為贛香B的成熟種子;轉化菌株為農桿菌EHA105;雙元轉化載體pCUbi3190由江西省農業科學院超級稻中心提供。
1.2 超表達載體的構建
以從越光材料中提取的cDNA為模板,用引物Os05g15770FKpnI(5′-GGGGTACCATGGCGTCCCGACGCCTTG-3′)和Os05g15770RSpeI(5′-GACTAGTTCACAGAACCTGATCCAGGAGACC-3′)以及KODFX酶(Takara)進行PCR擴增,獲得了OsHI-XIPCDS序列。用KpnI和SpeI酶切PCR純化產物及pCUbi1390表達載體,然后通過DNA連接酶進行連接,轉入大腸桿菌DH5a體內,通過篩選克隆、酶切檢測以及測序分析獲得了正確的OsHI-XIP超表達載體。最后電激轉化農桿菌EHA105,于-80 ℃下保存備用。
1.3 愈傷組織的誘導及繼代
將贛香B的成熟種子去殼后用70%乙醇浸泡1 min,隨后用50%次氯酸鈉處理30 min,在轉速180 r/min的搖床上搖晃。用無菌水沖洗8~10次,最后將沖洗好的種子放到滅菌濾紙上晾干5 min。在每個倒有誘導培養基的培養皿中放入12~13粒種子,放入(27±1)℃、黑暗環境下培養。經過14 d的黑暗誘導,挑選胚性愈傷組織,將其平均分成3等份,分別放入新的誘導培養基中在(27±1)℃、黑暗環境下繼代培養4 d。
1.4 農桿菌介導的轉化體系建立
從固體劃線平板上挑取農桿菌EHA105單克隆菌株,放入裝有5 mL YEB液體培養基的50 mL離心管中。將該離心管放置于200 r/min的搖床上,在28 ℃下暗培養20~24 h;之后將離心管中的培養液轉入加有100 mL YEB的500 mL三角瓶中,在相同培養條件下培養。當OD600到達1.0時,將農桿菌細胞在4 ℃條件下以8000 r/min離心15 min,進行富集;最后用懸浮培養基將農桿菌細胞重懸。將重懸后的農桿菌細胞浸入水稻愈傷組織,在轉速50 r/min的搖床上緩和培養20~25 min,用滅菌濾紙吸干愈傷組織上的多余菌液。之后將這些愈傷組織轉入共培養基,在(27±1)℃、黑暗條件下共培養48 h。一旦在愈傷組織中出現了少量農桿菌,即用加入了適當濃度的頭孢噻肟無菌水清洗8~10次,再用滅菌濾紙吸干,然后將其轉移到篩選培養基中,在(27±1)℃、黑暗條件下培養12 d。經過第一輪篩選培養后,挑選正常的愈傷組織,將其轉移到新的篩選培養基上進行第二輪培養。經過10 d的第二輪篩選培養,挑選新長出來的微小愈傷組織,再將其轉移到新的篩選培養基上進行第三輪篩選,在(27±1)℃、黑暗下培養5 d。將第三輪篩選培養基中長出的顆粒狀的微小愈傷組織轉移到分化培養基上,在(27±1)℃、黑暗條件下培養7 d;之后將愈傷轉移至新的分化培養基上,在(27±1)℃、光照條件下培養4 d。最后轉移分化出來的幼苗到生根培養基上,待幼苗長出較發達的茁壯根系時進行移栽煉苗。
1.5 轉基因植株的檢測
剪取植株的幼嫩葉片,參考王關林等的CTAB法提取DNA,采用PCR方法擴增檢測植株內的潮霉素(HPT)基因。所用引物hpt557-F為5′-ACACTACATGGCGTGATTTCAT-3′,hpt557-R為5′-TCCACTATCGGCGAGTACTTCT-3′。反應體系(10 μL): DNA模板30~50 ng、1.1×T3 Super PCR Mix 9.1 μL、10 μmol/L引物各0.2 μL。反應條件:98 ℃ 2 min;98 ℃ 10 s,54 ℃ 10 s,72 ℃ 10 s,32個循環;72 ℃ 2 min,25 ℃ 1 min。
2 結果與分析
2.1 超量表達載體OsHI-XIP+pCUbi1390的獲得
通過PCR擴增、酶切等過程將該基因連接到pCUbi1390載體上(圖1),然后轉入大腸桿菌,提取其質粒進行酶切或者PCR檢測及測序分析。共提取了15個質粒進行PCR檢測,結果只有3個質粒能擴增出目的條帶(894 bp)(圖2)。進一步對這3個質粒進行測序比對發現,OsHI-XIP+pCUbi1390-8質粒序列正確(圖3)。
2.2 農桿菌介導的水稻遺傳轉化
秈稻的遺傳轉化效率普遍較低,嚴重影響了秈稻品種轉基因工作的開展。為了提高秈稻的轉化效率,我們對農桿菌介導的贛香B轉化體系進行了系統研究。統計結果表明:贛香B的侵染愈傷數為500,其中抗性愈傷數為82,轉化效率為16.4%;檢測轉基因苗數為23,其中陽性轉基因苗數為16,陽性苗率為69.6%;這兩個參數顯著高于其它大部分秈稻品種,可見,贛香B作為轉化受體,其轉化效果顯著優于其他秈稻品種,是一個很好的秈稻轉基因受體材料。
2.3 轉基因植株的PCR檢測及陽性苗的獲得
以獲得的23株ZH11轉化苗(編號為19OSH513_1~23)葉片的總DNA為模板,以hpt557-F/R為引物,進行PCR擴增反應。然后分別取PCR產物5 μL,采用1%瓊脂糖凝膠電泳進行檢測。結果(圖4)顯示,有16株ZH11轉化苗擴增出潮霉素基因(HPT,557 bp),因此初步鑒定獲得了16株PCR陽性植株。最后,種植T0代的轉化苗,獲得了種子;目前已經種植到T1代苗期(圖5)。

圖2 部分OsHI-XIP+pCUbi1390質粒的PCR檢測結果
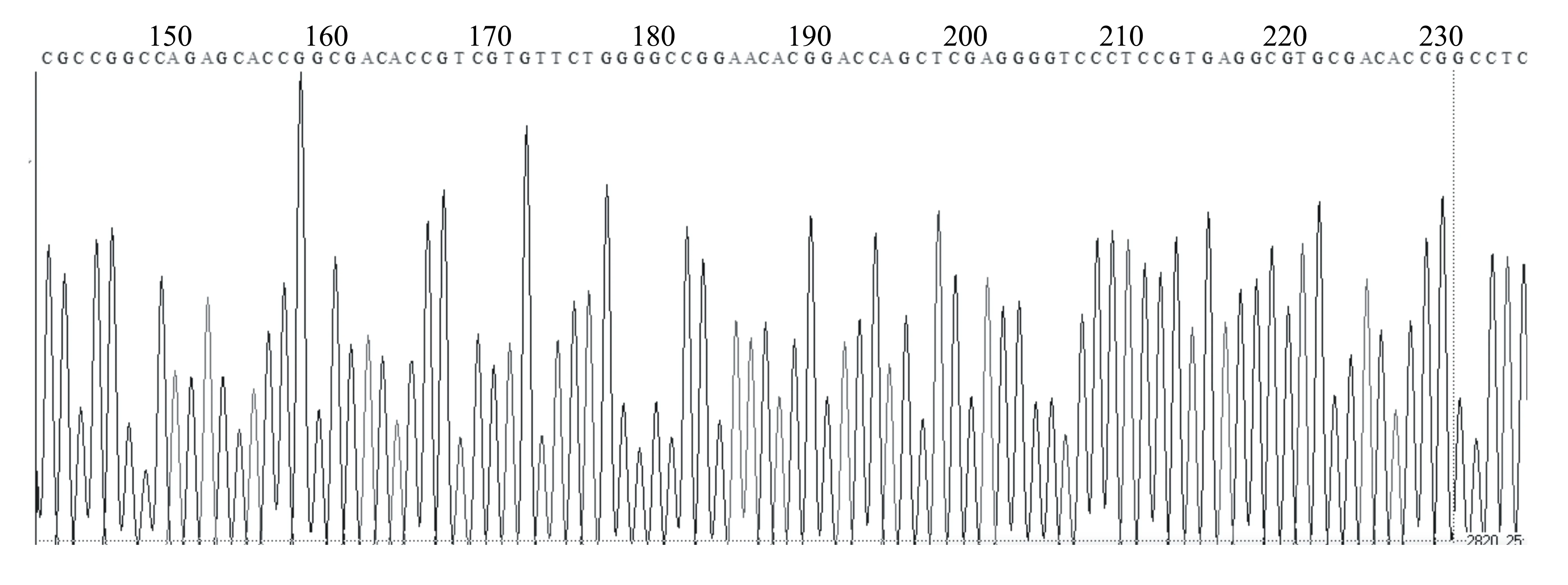
圖3 OsHI-XIP+pCUbi1390-8質粒中基因片段的測序結果
3 討論與結論
贛香B是本課題組選育的優質秈型香稻保持系,其母本為贛香A,具有開花習性好、異交結實率高、抗稻瘟病、米質優良且有香味等特點。目前,利用贛香A已成功配制雜交稻品種或組合15個,并且通過了福建、海南、云南、四川、重慶、貴州、浙江和江西等多個省(市)品種審定,如贛優明占、贛優607、贛香優702、贛優810等。由于贛香B的綜合農藝性狀優良,是研究水稻重要農藝性狀相關基因的重要材料,亟須建立該材料的遺傳轉化體系。目前,大部分秈稻品種的組織培養都面臨不同程度的困難,比如有些品種的愈傷組織不耐繼代,有些品種的抗性愈傷容易褐化死亡,有的品種的抗性愈傷不能分化成幼苗等,進而導致秈稻品種的轉化效率極低。本試驗首次成功建立了贛香B的遺傳轉化體系,而且轉化效率穩定維持在16%以上,顯著高于一般秈稻品種的轉化效率。因此,贛香B可以作為優良的秈稻轉化受體品種,從而建立起水稻高效率的遺傳轉化體系。

B代表空白對照;N代表陰性對照;P代表陽性對照。圖4 轉化再生植株PCR檢測結果

圖5 T1代的田間轉化苗
東野是迄今為止分布緯度最高的普通野生稻,除了蘊藏著豐富的種質資源、具有強抗逆性外,還擁有其他普通野生稻和栽培稻所沒有的強耐冷性,具有很大的育種價值。為了讓贛香B能夠獲得較強的耐冷性,本研究前期已構建以贛香B為受體親本,以東野為供體親本的滲入系群體,并通過冷脅迫處理篩選出了贛香B的耐冷株系。基于耐冷株系和贛香B的轉錄組比較分析,篩選出6個響應冷脅迫的基因,我們重點研究了其中的OsHI-XIP基因。該基因亞細胞定位于內質網上,編碼1個糖基水解酶;過量表達OsHI-XIP基因能夠增強水稻對植食性昆蟲的抗性[12]。目前尚未見該基因耐冷方面的報道。為了進一步研究OsHI-XIP的生物學功能,我們構建了OsHI-XIP的超表達載體,利用農桿菌介導法將OsHI-XIP基因轉入贛香B的愈傷組織中,通過篩選、分化培養獲得了轉基因苗(圖5)。這為后期進一步研究耐冷基因的生物學功能,為研究東野的耐冷機制提供了可能。本研究首次建立了以贛香B為受體的農桿菌介導的秈稻高效遺傳轉化體系,為后期研究優良農藝性狀相關基因的分離、克隆及生物學功能驗證奠定了堅實的材料基礎,提供了強有力的技術支撐,也為水稻分子育種奠定了基礎。
