銀腺楊84K莖段瞬時轉化體系的建立*
2021-05-21 08:22:38李曉軍黃李超盧孟柱
林業科學 2021年4期
李曉軍 安 軼 黃李超 曾 為 盧孟柱
(浙江農林大學林業與生物技術學院 省部共建亞熱帶森林培育國家重點實驗室 杭州 311300)
楊樹(Populus)是主要造林樹種之一,是目前再生生物資源和森林碳匯的重要組分,具有重要的經濟和生態價值。另外,楊樹生長快、適應性強、成材早、木材蓄積量大,且遺傳背景豐富、易于無性化擴繁和遺傳轉化,成為木本植物研究的模式物種(Janssonetal., 2007; Polleetal., 2013)。隨著毛果楊(Populustrichocarpa)、胡楊(P.euphratica)、新疆楊(P.bolleana)、銀腺楊84K(P.alba×P.glandulosa‘84K’)全基因組測序的完成(Tuskanetal., 2006; Maetal., 2013; 2019; Huangetal., 2020),為楊樹這一木本模式植物功能基因組的研究提供了基礎。對木本植物來說,高大的樹體需要龐大的維管組織來支撐,同時運輸水分和營養物質。木材就是這些龐大的維管組織的產物,來源于維管形成層的活動。所以了解維管形成層活動的調控機制及木質部的分化是利用現代生物技術對木材材性進行遺傳改良的基礎。但獲得穩定轉基因植株的效率低、周期長,這些限制阻礙了對木本植物木材相關基因功能的大規模高效率鑒定。因此,建立一種快速、便捷、高效的方法,用以揭示木材形成分子機制,變得尤為重要。
瞬時轉化技術可以避開冗長的轉基因過程,只需將目的基因導入組織或個體,不涉及再生過程,轉化之后直接進行基因表達分析或表型鑒定,具有操作簡單、周期短、通量高的優點,可作為穩定轉化的補充,特別適合應用于植物自身或異源基因功能及代謝途徑的大規模遺傳研究,并且可對無性繁殖困難的木本植株及同源植物系統(Korolevaetal., 2005; Yooetal., 2007)進行基因功能分析。近年來已有木本植物建立了瞬時轉化體系,用于品種的選育與開發,基因槍轉化(Nowaketal., 2004; Fizreeetal., 2019)、PEG介導原生質體轉化(Yooetal., 2007)、植物病毒載體介導轉化(Kumagaietal., 1995)和基于農桿菌(Agrobacterium)介導(Yangetal., 2000; Simmonsetal., 2009)的轉化方法已成功應用于梨(Pyruscommunis)(Spolaoreetal., 2001)、甜橙(Citrussinensis)(de Oliveiraetal., 2009)、白樺(Betulaplatyphylla)、剛毛檉柳(Tamarixhispida)、黃檗(Phellodendronamurense)(Zhengetal., 2012)、桑(Morusalba)(Wuetal., 2015)、桂花(Osmanthusfragrans)(Hanetal., 2016)、柿(Diospyroskaki)(Moetal., 2019)、蘋果(Malusdomestica)(Lvetal., 2019)等。在楊樹中,通過PEG介導法成功獲得了瞬時轉化的葉片原生質體(Tanetal., 2013; Guoetal., 2015); 通過基因槍轉化法成功獲得了瞬時轉化的葉片表皮細胞和保衛細胞(Nowaketal., 2004); 通過農桿菌侵染技術獲得了瞬時轉化的葉片(Takataetal., 2012)和芽(Yangetal., 2009),這都為木本植物的基因調控研究提供了快速可靠的技術支持。
然而,上述瞬時轉化技術只局限于原生質體、離體的葉片或芽,不利于研究木本植物形成層活動和木質部分化相關基因的功能。因此,本研究以銀腺楊84K 1年生休眠莖段為材料,通過水培觀察,表明其具有典型的形成層活動和木質部分化。以人工增強型綠色熒光蛋白(eYGFP)基因作為報告基因,建立農桿菌介導的真空滲透侵染莖段的轉化體系,實現在形成層區域的外源基因表達,為木材形成和生長發育相關基因的研究提供了一種快速、高效的轉化方法,以促進木本植物木材形成機制的研究。
1 材料與方法
1.1 試驗材料
于2019年11月,在河北任丘中國林業科學研究院林業研究所試驗基地,采集1年生銀腺楊無性系84K休眠期枝條。截取直徑約1 cm、長度約10 cm的莖段,封口膜包裹莖段兩端以防止失水,并低溫(4 ℃)保存。
1.2 載體構建和根癌農桿菌轉化及鑒定
構建表達載體pK2GW7-eYGFP。首先根據Chin等(2018)所用載體克隆eYGFP表達序列35S-COR47-5′-UTR-eYGFP-HSP-T878(1 991 bp),并通過酶切位點StuⅠ,使用同源重組方法將此序列構建到pK2GW7載體中(pK2GW7載體由浙江農林大學韓瀟博士惠贈)。引物為F1(5′-CTGACCCACAGA TGGTTAGAGAGGTGAGACTTTTCAACAAAGGGTAAT TT-3′)和R1(5′-GATGAGACCTGCCGCGTAGGTG TGTACAGATATATGTTGAATTATTGA-3′)。轉化大腸桿菌(Escherichiacoli)后,在含壯觀霉素(50 mg·L-1)的LB固體培養基中培養12 h,挑選陽性單克隆進行PCR鑒定。鑒定引物為F2(5′-CAGATGGTTAGAGAGGTGAGACTTTT-3′)和R2(5′-GACCTGCCGCGTAGGTGTGTACAGATAT-3′)。鑒定成功后提取質粒并利用凍融法將表達載體轉化至根癌農桿菌(Agrobacteriumtumefaciens)GV3101感受態細胞中,并PCR鑒定。表達載體pK2GW7轉化至根癌農桿菌GV3101感受態細胞用于試驗對照。
1.3 形成層活動的觀察
將休眠莖段從低溫環境取出,將其垂直培養在水中。對水培0、5、10、15、20、25、30天的莖段1/2處進行切片(40 μm,LEICA VT1200S)、20倍數顯微鏡下觀察形成層活動(LEICA DM6)。培養條件為: 16 h光照/8 h黑暗,23~25 ℃,光照強度50 μmol·m-2s-1。
1.4 遺傳轉化操作
采用凍融轉化法,將pK2GW7質粒和含eYGFP基因的雙元載體pK2GW7的質粒轉入農桿菌GV3101,然后將農桿菌于固體LB培養基(50 mg·L-1壯觀霉素)上培養2天,培養溫度為28 ℃。挑取單克隆進行PCR檢測。檢測正確后按1∶100(V/V)的比例取活化菌液接種于200 mL YEB液體培養基(50 mg·L-1利福平和50 mg·L-1壯觀霉素)中,并置于28 ℃、200 r·min-1搖床內過夜,搖菌至OD600= 0.8。分別對不帶載體、帶有pK2GW7、帶有pK2GW7-eYGFP的3種GV3101農桿菌菌液,在激光共聚焦顯微鏡(ZEISS,LSM 880)下觀察,均未發現任何信號,故排除農桿菌本底表達的影響。收集pK2GW7、pK2GW7-eYGFP菌體,菌體用重懸液(1/2MS,25 g·L-1蔗糖,10 mmol·L-1MES, 200 μmol·L-1AS,pH5.6)重懸。轉化前將低溫保存的84K楊莖段在含25%蔗糖的1/2 MS溶液中浸泡3 h,可促進轉化(Zhengetal., 2012)。將浸泡過的莖段豎直放入裝有重懸液的三角瓶中,莖段形態學下端浸入重懸液中,形態學上端與真空儀器通過橡膠管連接,在0.08 MPa壓強下,懸浮液從莖段形態學下端抽濾至形態學上端,轉化完畢的莖段進行水培,培養條件為: 16 h光照/8 h黑暗,23~25 ℃,光照強度50 μmol·m-2s-1。
1.5 農桿菌介導的銀腺楊84K莖段瞬時轉化單因素試驗
根據楊樹遺傳轉化體系(Hanetal., 2000; Anetal., 2020)和桂花、朱槿(Hibiscusrosa-sinensis)、蘋果、剛毛檉柳等其他木本植物遺傳轉化體系(Zhengetal., 2012; Trivellinietal., 2015; Hanetal., 2016; Lvetal., 2019),本試驗選擇了影響農桿菌介導的84K楊莖段瞬時轉化效率的轉化因子進行研究: 乙酰丁香酮(AS)濃度、農桿菌濃度、真空滲透侵染時間、真空滲透侵染次數、侵染后培養時間。所有因子均設某單一因素為變量進行篩選,以報告基因eYGFP瞬時表達反映各因素對瞬時轉化率的影響(瞬時轉化率 = 觀察到eYGFP綠色熒光信號的莖段數目/總轉化莖段數目),每個處理3次重復,每個重復至少6個莖段(P< 0.05; ANOVA)。
1.6 農桿菌介導的銀腺楊84K莖段瞬時轉化因子正交試驗設計
為探究不同因素對84K楊莖段瞬時轉化的相互影響作用,本試驗進行了不同因素正交試驗以選擇最優的因子組合。以eYGFP瞬時轉化率為考察指標,采用L9(34)正交試驗進行試驗設計(表1)。正交試驗每個處理3次重復,每個重復至少6個莖段。采用SPSS17.0軟件對數據進行分析。
1.7 銀腺楊84K莖段瞬時轉化效率分析
轉化后的84K楊莖段可用LUYOR-3415RC手持熒光燈進行eYGFP初步觀察分析。在莖段1/2處進行切片,切片厚度為40 μm,并使用共聚焦顯微鏡(ZEISS,LSM 880)在493~558 nm的藍色激發光下觀察熒光信號。統計觀察到的eYGFP熒光信號,計算瞬時轉化率。
2 結果與分析
2.1 解除休眠后莖段形成層的活動及木質部分化
對休眠的莖段進行水培。如圖1A所示,培養0天時,形成層排列緊密,呈皺縮狀; 隨著培養天數增加,第5天時的形成層細胞吸水逐漸恢復扁平狀態,形成層寬度和層數較0天時變化不顯著(圖1B、C); 培養10天時,形成層層數顯著增加,寬度增加不顯著; 培養15天時,可觀察到新形成的次生木質部,相較之前形成的木質部細胞,其細胞壁較薄。其中可見體積膨大的細胞,為新形成的導管。隨著培養天數增加,新形成的次生木質部區域逐漸增加,并伴隨著大量新導管產生。培養20~30天期間,形成層寬度增加顯著,層數為5或6層。這表明1個月內水培休眠枝條可以模擬正常木材形成過程,新生的木質部細胞與前期形成的木質部沒有顯著差異。
2.2 載體構建及農桿菌轉化鑒定
為建立銀腺楊84K莖段瞬時轉化系統,根據Chin 等(2018)所用載體克隆增強型eYGFP表達序列。將擴增產物通過同源重組方法連接到pK2GW7載體中,質粒轉入農桿菌感受態GV3101中待用。pK2GW7-eYGFP載體如圖2所示。
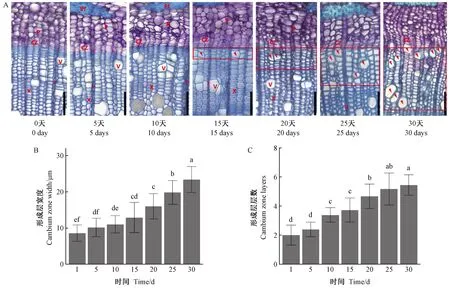
圖1 室溫水培解除休眠后莖段形成層的活動和木質部分化Fig. 1 Cambium activity and xylem differentiation of dormancy-released stem segments cultured in water at room temperature A: 不同培養天數形成層變化。CZ: 形成層; P: 韌皮部; Pf: 韌皮纖維; r: 射線; V: 導管; X: 木質部。紅框代表新形成的次生木質部; 箭頭代表新形成的導管。標尺為100 μm。A: Changes of cambium zone following cultured days. CZ: Cambium zone; P: Phloem; Pf: Phloem fibre; r: Ray; V: Vessel; X: Xylem. Red rectangles indicate the newly secondary xylem; Arrowheads point to new vessels. Scale bars: 100 μm. B, C: P < 0.05; one-way ANOVA.下同。The same below.
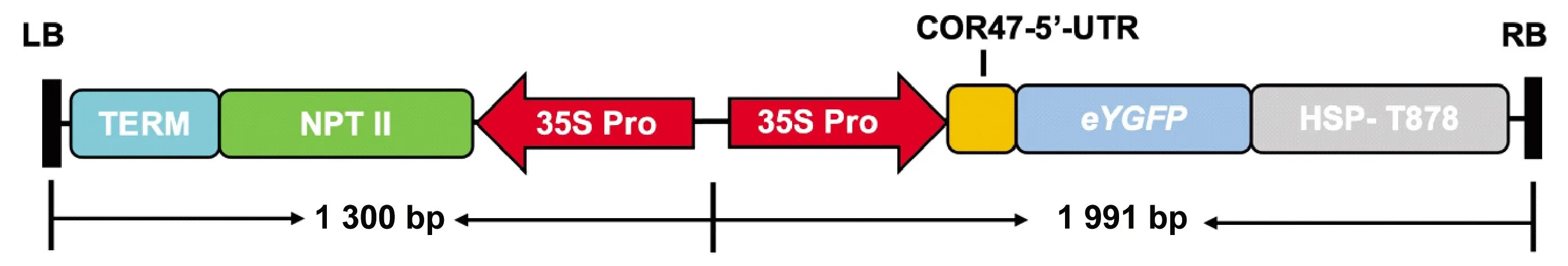
圖2 pK2GW7-eYGFP表達載體示意Fig. 2 Schematic diagram of the pK2GW7-eYGFP vector
2.3 不同因素對84K楊莖段轉化的影響
2.3.1 乙酰丁香酮(AS)對eYGFP基因瞬時轉化率的影響 添加AS與不添加AS相比,eYGFP基因瞬時轉化效率明顯增加。當AS濃度為100、200、300 μmol·L-1時,轉化效率無顯著變化(圖3A)。故添加AS進行后續試驗,選擇AS濃度為200 μmol·L-1。
2.3.2 菌液濃度對eYGFP基因瞬時轉化率的影響 農桿菌濃度OD600為0.3時瞬時轉化eYGFP基因成功率最低,隨著農桿菌濃度增加,轉化率增加; 當農桿菌濃度OD600增加到0.9時,瞬時轉化效率達到最高。隨著農桿菌濃度提高到1.2或1.5時,轉化效率降低(圖3B)。說明菌液濃度過高或過低均會影響轉化效率。農桿菌濃度過高,加快了莖段的腐爛速度,導致莖段生理狀態較差從而影響轉化率。
2.3.3 真空滲透侵染時間對eYGFP基因瞬時轉化率的影響 如圖3C所示,瞬時轉化率在真空滲透侵染時間5~15 min時較低,在達到20 min時顯著增加,在延長到30 min時反而下降。表明,真空滲透侵染達到一定時間后處于侵染飽和狀態,抽吸時間過長反而引起莖段結構損傷,影響轉化率。
2.3.4 真空滲透侵染次數對eYGFP基因瞬時轉化率的影響 隨著真空滲透侵染次數增加(每2次真空滲透侵染間隔72 h),轉化率呈現出先增后降的趨勢。當用農桿菌重懸液真空滲透侵染2次時,瞬時轉化率最高; 減少或增加真空滲透侵染次數,轉化率未顯著增加; 且當真空滲透侵染5次時瞬時轉化率最低(圖3D)。表明真空滲透侵染會不同程度地損傷莖段,抽吸次數越多損傷越大,導致轉化率越低。
2.3.5 培養天數對eYGFP基因瞬時表達的影響 真空滲透侵染后,將莖段進行水培。水培天數對瞬時轉化率也有較大影響(圖3E)。當水培天數較短時,如3天和6天,均觀察不到綠色熒光信號; 當培養到第9天時,可觀察到綠色熒光; 水培至12天時瞬時轉化率最高。隨后,隨培養時間增加,轉化率降低。

圖3 不同因素對84K楊莖段真空滲透轉化的影響Fig. 3 Effects of key factors on transient transformation of poplar 84K stem segments via vacuum infiltration
2.4 農桿菌介導的84K楊莖段瞬時轉化體系的優化
為優化農桿菌介導的84K楊莖段瞬時轉化效率,根據上述單因素試驗結果,通過L9(34)的正交試驗設計(表1),共進行了9組不同水平處理的轉化因子組合試驗。表明水培天數、真空滲透侵染時間均對eYGFP瞬時轉化率有顯著影響(P< 0.05),即水培天數和真空滲透侵染時間顯著影響農桿菌介導銀腺楊84K莖段瞬時轉化率,而菌液濃度和真空滲透侵染次數的影響不顯著(表2)。4個因素對轉化率的影響為: 侵染后水培天數>真空滲透侵染時間>菌液濃度>真空滲透侵染次數。
通過表1顯示,農桿菌介導的真空滲透瞬時轉化銀腺楊84K莖段的最優組合是農桿菌重懸液濃度為OD600= 0.9、真空滲透侵染20 min、真空滲透侵染2次、侵染后水培15天。
2.5 瞬時轉化后eYGFP的表達觀察
瞬時轉化后進行水培,經切片顯微觀察,維管組織發育情況與休眠莖段水培的維管組織發育情況類似。形成層細胞從最初的皺縮狀恢復到扁平狀,形成層層數也隨之增加,培養15天可觀察到新形成的次生木質部細胞。瞬時轉化后的莖段維管發育也可追蹤從形成層活動到木質部形成的全過程。84K楊莖段瞬時轉化后水培15天時的eYGFP表達情況初步觀察如圖4所示。圖4A為正常光下莖段整體情況,莖段下部剝掉部分樹皮,露出形成層區域。經激發光照射在已轉化的莖段形成層區域可觀察到eYGFP綠色熒光信號(圖4B中紅箭頭所示),紅色為自發熒光。正常光下莖段橫截面如圖4C所示,照射激發光后在莖段橫截面上也可觀察到明顯的eYGFP綠色熒光信號(圖4D)。

表1 84K楊莖段瞬時轉化因子L9 (34)正交試驗設計(均值±標準誤)Tab.1 Factors and levels of L9 (34) orthogonal experiments(Mean ± SE)
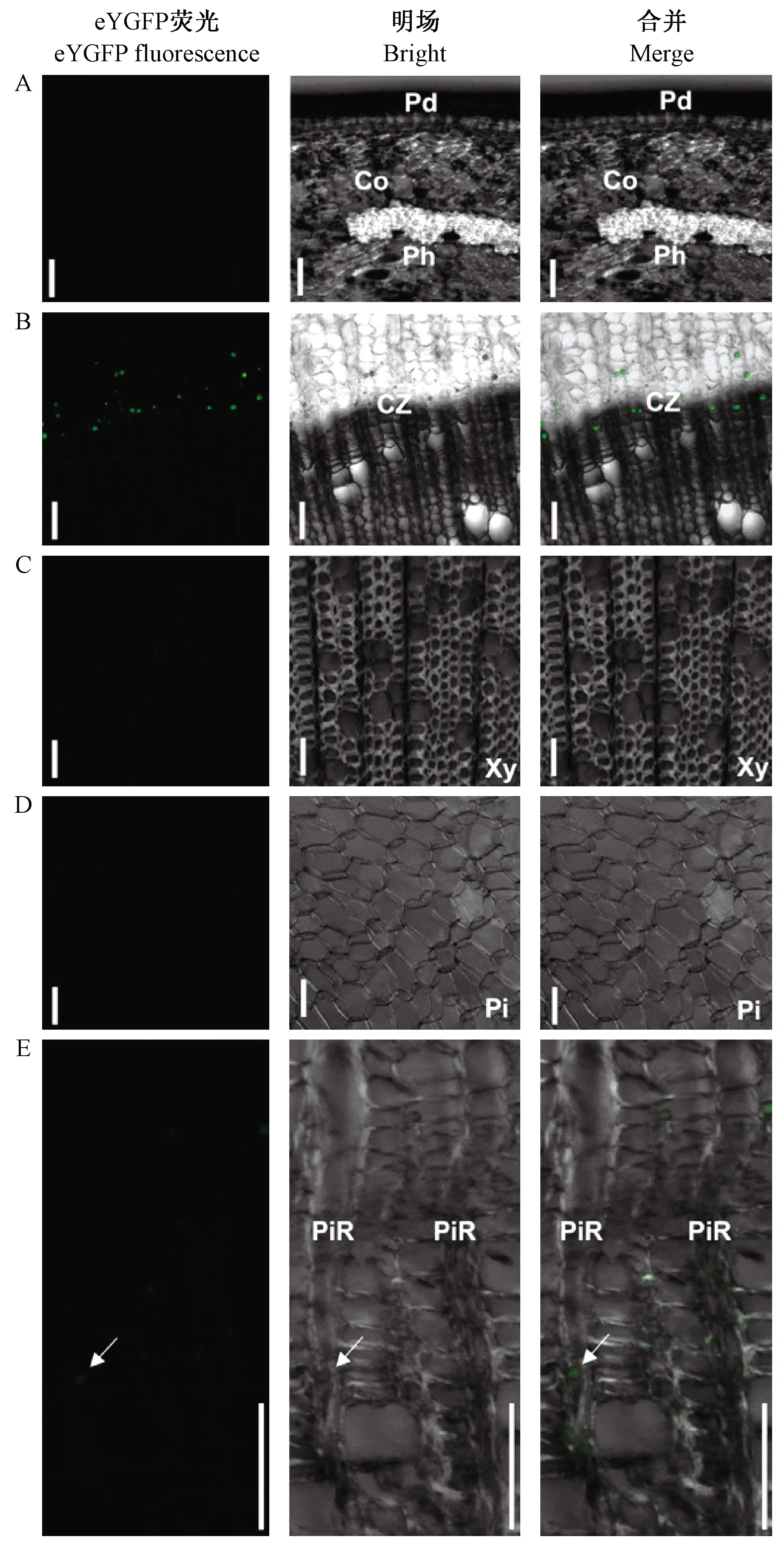
圖5 瞬時轉化后eYGFP在不同部位的表達情況Fig. 5 eYGFP expression in different regions A: 周皮(Pd)、皮層(Co)、韌皮部(Ph); B: 形成層區域(CZ); C: 木質部(Xy); D: 髓(Pi); E: 髓射線(PiR)。標尺= 50 μm。A: Periderm(Pd), Cortex(Co), and Phloem(Ph); B: Cambium zone(CZ); C: Xylem(Xy); D: Pith(Pi); E: Pith rays(PiR). Scale bars = 50 μm.
為進一步探究84K楊莖段轉化后eYGFP綠色熒光信號具體部位,對莖段1/2處進行切片觀察。通過共聚焦顯微鏡觀察不同部位eYGFP的表達情況,在周皮、皮層、韌皮部、木質部、髓中未觀察到綠色熒光信號(圖5A、C、D)。而在形成層區域細胞中可觀察到較多的綠色熒光信號(圖5B),同時在髓射線中也可觀察到較少的微弱熒光信號(圖5E)。結果顯示瞬時轉化后eYGFP的表達主要集中在形成層區域。說明外源基因可以在形成層細胞中表達,有利于鑒定它們在形成層活動及分化為木質部中的作用。
3 討論
現代生物技術的迅猛發展已推動林木研究從傳統育種轉向分子育種,而利用現代生物技術對木材進行遺傳改良,首先必須了解木材形成的復雜過程。目前,研究木本植物的木材相關基因大多以穩定的遺傳轉化植株為材料。穩定遺傳轉化技術需要把目的基因整合到宿主基因組中,使宿主具有目的基因所表現的性狀、功能和表型。多數穩定的農桿菌介導轉化需要再生體系,這是一個繁瑣且漫長的過程。由于木本植物具有雜合性高、繁殖時間長、轉化效率低等特點,不能快速獲得大量轉基因植株,且仍有相當一大部分木本植物再生體系沒有建立,遺傳改良受到限制(Gambinoetal., 2012; van Nockeretal., 2014)。與穩定遺傳轉化體系相比,瞬時轉化技術不涉及再生過程,可在轉化之后直接進行表型鑒定,周期短且轉化效率高(Uenoetal., 1996)。然而,瞬時轉化大多以葉片(Tanetal., 2013; Guoetal., 2015)為轉化材料,雖轉化體系較為成熟,但轉化過程損壞了轉化材料使其無法繼續正常生長。而且由于轉化材料組織的限制,更不利于木本植物木材發育相關基因的研究。
本試驗在不破壞植物材料的前提下,通過真空滲透侵染的方法建立了農桿菌介導的銀腺楊84K莖段瞬時轉化體系。試驗通過研究菌液濃度、真空滲透侵染時間、真空滲透侵染次數、侵染后水培天數等轉化因子,采用正交設計優化試驗,結果表明真空滲透侵染時間和水培天數對eYGFP瞬時轉化率有顯著影響。真空滲透侵染方法通過真空壓力將農桿菌菌液從莖段下端抽至莖段上端,此操作方便快捷且可批量轉化。此方法增加了84K楊莖段導管的運輸能力和細胞的滲透性,使載體更容易進入植物細胞。切片觀察結果表明eYGFP報告基因主要在恢復活力的形成層區域中表達(圖5)。說明相較于其他的葉或芽等材料的瞬時轉化體系,本體系能提供形成層細胞相關基因的原位表達,這都有利于鑒定它們在形成層活動方面的作用。試驗結果表明,侵染后水培9~20天后目標基因的表達水平都較高(圖3E),農桿菌轉化后直接進行水培,15天之前休眠莖段的主要活動是形成層恢復活力,形成層層數和寬度都有所增加。如果研究的目標基因參與形成層細胞活動,則可在15天之前進行形成層細胞的早期表型變化觀察。而15天左右可觀察到新形成的次生木質部。因此研究木質部發育相關基因的功能,可在15天后再進行維管組織發育的表型變化觀察。本體系的建立可以追蹤木質部形成的全過程,所以可利用它研究形成層及木質部發育的相關基因功能,但需要注意觀察分析時間。
建立高效的瞬時表達系統,有助于植物基因功能的研究。本體系在不破壞植物材料的基礎上,可快速、便捷、高效地獲得瞬時轉化材料,特別有助于研究維管組織發育相關基因。此方法建立對揭示木材形成的分子機制有重要意義,同時也可為研究其他木本植物木材形成相關基因提供借鑒。
4 結論
本研究建立了銀腺楊84K莖段真空滲透瞬時轉化體系: 農桿菌重懸液OD600為0.9,抽真空20 min,根癌農桿菌真空滲透侵染2次,水培15天。外源基因主要在銀腺楊84K莖段形成層細胞中表達,可以快速鑒定其在維管組織分化中的功能,有助于研究木質部發育的調控機制,也可為其他木本植物的木材品質研究提供新思路。
