飼料中丁酸鈉添加水平對草魚生長、脂代謝及健康的影響
2021-05-14 04:03:48晏顯芳于海波周繼術(shù)
畜牧獸醫(yī)雜志 2021年6期
關(guān)鍵詞:血清
晏顯芳,張 明,郭 盼,李 楊,于海波,吉 紅,周繼術(shù)
(西北農(nóng)林科技大學,陜西 楊凌 712100)
草魚(Ctenopharyngodonidella)是一種重要的淡水養(yǎng)殖魚類,大約占全球淡水養(yǎng)殖業(yè)的16%(Wang et al, 2015)。目前,在世界范圍內(nèi)的淡水養(yǎng)殖魚類中,草魚的產(chǎn)量最高(張露等,2016)。草魚由于其生長速度快、肌肉品質(zhì)細膩鮮美、養(yǎng)殖效益好等優(yōu)點深受廣大消費者和養(yǎng)殖戶的青睞。然而,隨著草魚高密度養(yǎng)殖模式的普及和高能飼料的普遍使用,其肝脂代謝紊亂及腸道損傷情況越來越多見,肝臟變性、脂肪肝等疾病頻繁發(fā)生,嚴重影響魚體的健康狀況。
丁酸屬短鏈脂肪酸,可作為腸道上皮的直接能量來源,還具有重要的生理調(diào)控作用;不僅可促進小腸黏膜的生長發(fā)育,增強黏膜的通透性,修復受損腸道上皮細胞,增加絨毛的寬度,提高小腸消化吸收功能,還能促進腸道上皮細胞增殖,并修復腸道受損部位,從而促進腸道健康。由于丁酸具有揮發(fā)性及特殊氣味,在生產(chǎn)上常用丁酸鈉替代。已有研究表明,丁酸鈉能改善鯉魚、羅非魚與鯽魚的腸道健康,但在草魚飼料中尚未見相關(guān)報道。研究還發(fā)現(xiàn),丁酸通過降低胰島素的敏感性,從而降低小鼠白色脂肪組織中的脂質(zhì)積累;而這種抑制脂質(zhì)蓄積的作用未能在小鼠的肝臟和肌肉組織中發(fā)現(xiàn)。丁酸在魚類或在草魚中是否也具有相應的調(diào)節(jié)脂質(zhì)蓄積的作用,相關(guān)報道較為缺乏。
為探索丁酸鈉對草魚生長、脂代謝及健康狀況的影響,本研究在草魚飼料中添加不同水平的丁酸鈉,通過56 d的飼養(yǎng)試驗及檢測相關(guān)指標,以期探索丁酸對草魚生長、脂質(zhì)代謝、腸道組織結(jié)構(gòu)、免疫應答的影響,為丁酸鈉在草魚飼料中的適宜添加水平提供基礎(chǔ)資料。
1 材料和方法
1.1 試驗飼料
試驗飼料所用的基礎(chǔ)飼料原料由魚粉、豆粕、菜粕、棉粕、豆油等組成。丁酸鈉由新奧生物科技有限公司(福建省廈門市)提供。在基礎(chǔ)飼料中分別添加0、500、1000 mg Kg-1丁酸鈉,制成等氮(約36%粗蛋白)和等能(約6%粗脂)的三組試驗飼料(SB0、SB500及SB1000)。將飼料原料粉碎,過60目篩,混勻后,由飼料顆粒機制成直徑2 mm的顆粒飼料。飼料在室溫下風干12 h后-20℃保存?zhèn)溆谩o暳吓浞匠煞趾统R?guī)營養(yǎng)成見表1。
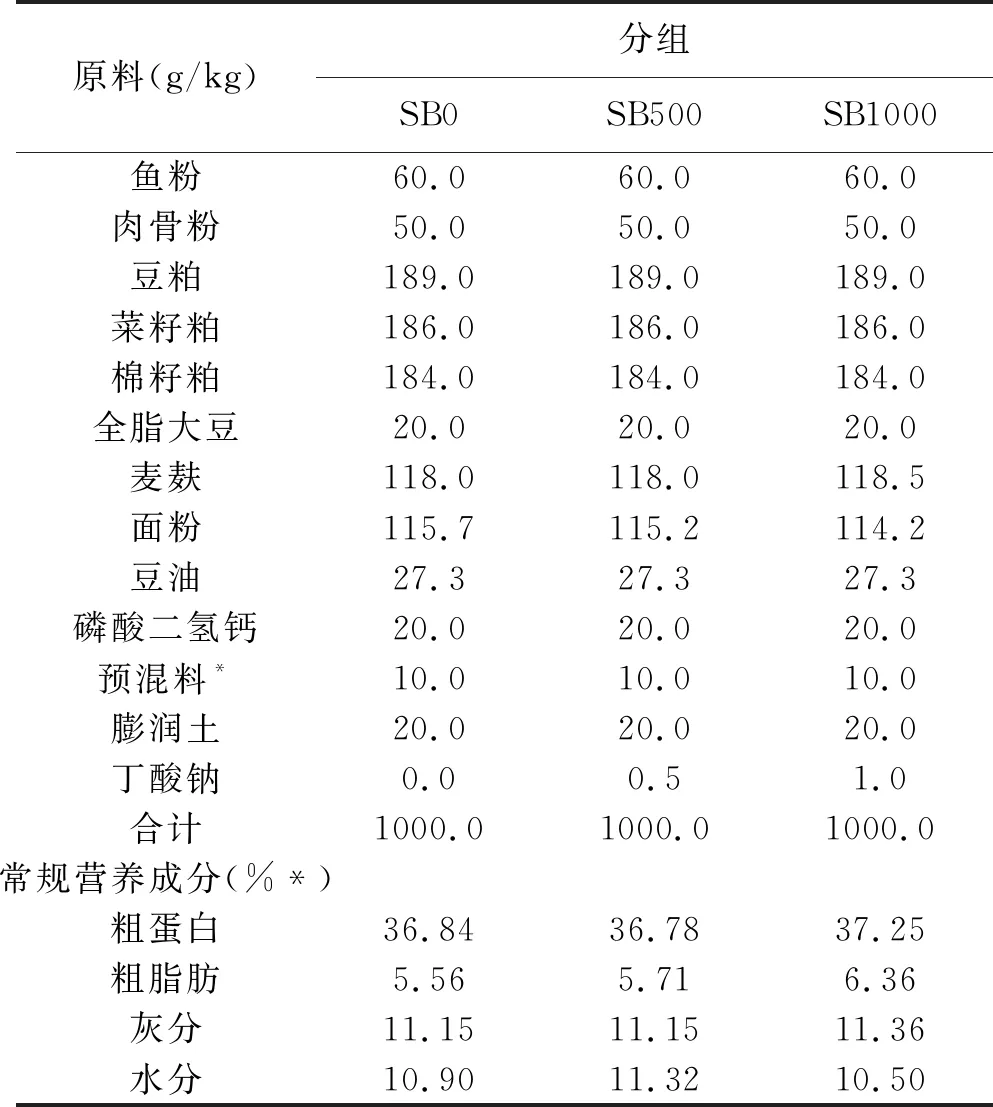
表1 試驗飼料配方組成以及常規(guī)營養(yǎng)成分(風干基礎(chǔ))
1.2 試驗魚的分組及飼養(yǎng)管理
將153尾草魚(14.10±0.60 g)隨機分為3個組,每組三個重復,每個重復17尾,養(yǎng)殖于9個循環(huán)流水養(yǎng)殖缸(130 L)中,分別飼喂以上三組試驗飼料。預飼三周后,進行正式飼喂。飼養(yǎng)試驗中,每天投飼3次(8∶30、12∶30、16∶30),投飼率為2%~3%,飼養(yǎng)周期為56 d。保持流水水量5 L min-1,定期檢測相關(guān)水質(zhì)參數(shù)。試驗期間,水溫25.50±2℃,pH 7.65±0.20,溶解氧7±0.50 mg L-1,硝酸鹽0.09±0.01 mg L-1。
1.3 采樣
飼喂結(jié)束后,饑餓24 h,然后用三胺甲基磺酸鹽(MS-222, 0.06 g L-1)麻醉。進行尾靜脈采血,保存在4℃冰箱中,6 h后離心(1 000 rpm,10 min)制備血清。將每尾魚稱重并解剖,分別剖取草魚肝胰臟、腹脂、腸、腎和脾,分別稱重。按以下公式進行魚體生長及生物學性狀的計算。
特定生長率Specific growth rate(SGR)= (ln終重- ln初重)/飼養(yǎng)時間(d)×100;
投飼量Feed intake(F1,%,魚/d) =飼料總消耗量(g);
飼料系數(shù)Feed conversion ratio(FCR)=飼料投飼量(g)/魚體增重量(g);
肥滿度Condition factor(CF,g/cm3%)=體重(g)/體長(cm)3×100;
肝臟指數(shù)Hepatopancreas index(HSI,%)=肝胰臟重量(g)/魚體重(g)×100;
脾臟指數(shù)Spleen index (SI,%)=脾臟重量(g)/體重(g)×100;
腸長比Relative intestine length (RIL, %)=腸長(cm)/魚體長(cm)×100;
內(nèi)臟指數(shù)Viscerosomatic index(VSI,%)=內(nèi)臟重量(g)/體重(g)×100;
腸體質(zhì)量比Intestinal index (II, %)=腸重(g)/體重(g)×100;
腹臟脂肪指數(shù)Intraperitoneal fat index(IFI, %)=腹腔內(nèi)脂肪重量(g)/體重(g)×100;
存活率Survival rate (SR, %)=末期魚數(shù)量/初始魚數(shù)量(g)×100;
將每箱3尾魚的肝胰臟和小腸凍存于液氮中,并于-80℃保存,用于分析與免疫應答相關(guān)的基因表達。每箱取3尾魚的腸道收集到5 mL離心管中,冷凍于液氮中,-80℃保存,用于腸道菌群分析。另外2尾魚切除肝胰臟、腸和肌肉,保存于-20℃,進行常規(guī)營養(yǎng)成分分析。同時,將另外3尾魚的肝胰臟和中腸固定在多聚甲醛溶液中進行HE染色及組織學觀察與分析。
1.4 血清生化指標測定
采用全自動生化分析儀(日立7180,東京,日本)對草魚血清進行分析。血清生化指標包括丙氨酸氨基轉(zhuǎn)移酶(ALT)、天門冬氨酸氨基轉(zhuǎn)移酶(AST)、總蛋白(TP)、白蛋白(ALB)、球蛋白(GLO)、葡萄糖(Glu)、膽固醇(Chol)、甘油三酯(TG)。
1.5 飼料和魚肉的常規(guī)營養(yǎng)成分檢測
檢測并分析了飼料和魚體肝胰臟、肌肉和腸道的常規(guī)營養(yǎng)成分,其中粗蛋白含量用凱氏定氮法,粗脂肪含量用乙醚抽提法,粗灰分含量馬弗爐550℃灼燒法進行測定,具體方法見AOAC(2000)。
1.6 肝胰及腸道組織的HE染色觀察
將固定后的肝胰臟和小腸于自來水中洗滌12 h,然后常規(guī)乙醇脫水和二甲苯平衡,隨后,按照之前所述的標準組織學程序,用紗布包裹樣品(Liu et al, 2016) ,再使用旋轉(zhuǎn)切片機(RM2235,德國萊卡)連續(xù)切割厚5 μm的石蠟切片,有蘇木精和伊紅染色(H&E)染色。使用倒置顯微鏡(OPTEC)拍攝切片組織樣品。
1.7 實時定量檢測基因相對表達量
分別從3尾魚的肝胰臟和腸組織中檢測tlr22和myd88基因的表達。使用TRNzol試劑(TaKaRa,中國大連)提取草魚肝胰臟和腸組織中總RNA。用所需量的無酶水溶解RNA,在260 nm~280 nm用電泳2%瓊脂糖凝膠和UV-Vis光譜儀檢測RNA的質(zhì)量和數(shù)量。在20 μL的反應體積內(nèi),將清洗后的總RNA反向轉(zhuǎn)錄1 μg制備所有的cDNAs。qRT-PCR使用CFX 96實時PCR檢測系統(tǒng)(Bio-Rad, Hercules, CA, USA)進行。PCR反應的總體積為20μL,其中每個引物0.6μL(10μM),cDNA 1μL,2×SYBR Premix Ex TaqTMII 10 μL,無酶雙蒸水7.8 μL。循環(huán)條件為:95℃ 3 min, 95℃ 15 s,60℃ 30 s, 65℃ 5 s,38次循環(huán)。目的基因tlr22及myd88的引物序列見表2。采用 (Livak and Schmittgen 2001)中描述的相對定量方法計算基因相對表達量。

表2 實時定量PCR中使用的引物
1.8 腸道微生物區(qū)系檢測
分析腸道菌群時,使用PowerSoil DNA分離試劑盒從魚糞混合樣本中提取總DNA。采用瓊脂糖凝膠電泳檢測DNA質(zhì)量,納米滴檢測DNA純度,然后用引物338F(5’- ACTCCTACGGGAGGCAGCA-3’)和806R(5’- GGACTACHVGGGTWTCTAAT-3’)擴增16S rRNA基因的V3-V4區(qū)。擴增產(chǎn)物采用磁珠純化法純化。對純化后的產(chǎn)物進行Solexa PCR,再用1.8%瓊脂糖凝膠在120V恒壓下電泳40 min,使用IlluminaHiSeq 2500 (Biomarker,北京,中國)切除、純化并測序相應的條帶。
1.9 統(tǒng)計分析
試驗數(shù)據(jù)以平均值±標準差(Mean±SD)表示。結(jié)果采用單因素方差分析,所有分析均使用PASW Statistics 18(SPSS,IL,USA)進行,然后進行Duncans檢驗,P<0.05為差異顯著。
2 結(jié)果
2.1 草魚生長性能和生物學參數(shù)
飼養(yǎng)8周后,SB1000組草魚臟體比(VSI)顯著低于對照組(P< 0.05);SB500和SB1000組草魚腹脂指數(shù)(IFI)顯著低于對照組(P<0.05);其它指標,如草魚初始體重(IBW),特定生長率(SGR)、飼料轉(zhuǎn)化率(FCR)、肥滿度、肝胰臟指數(shù)及脾臟指數(shù)等在各組間均無顯著差異(P> 0.05)。
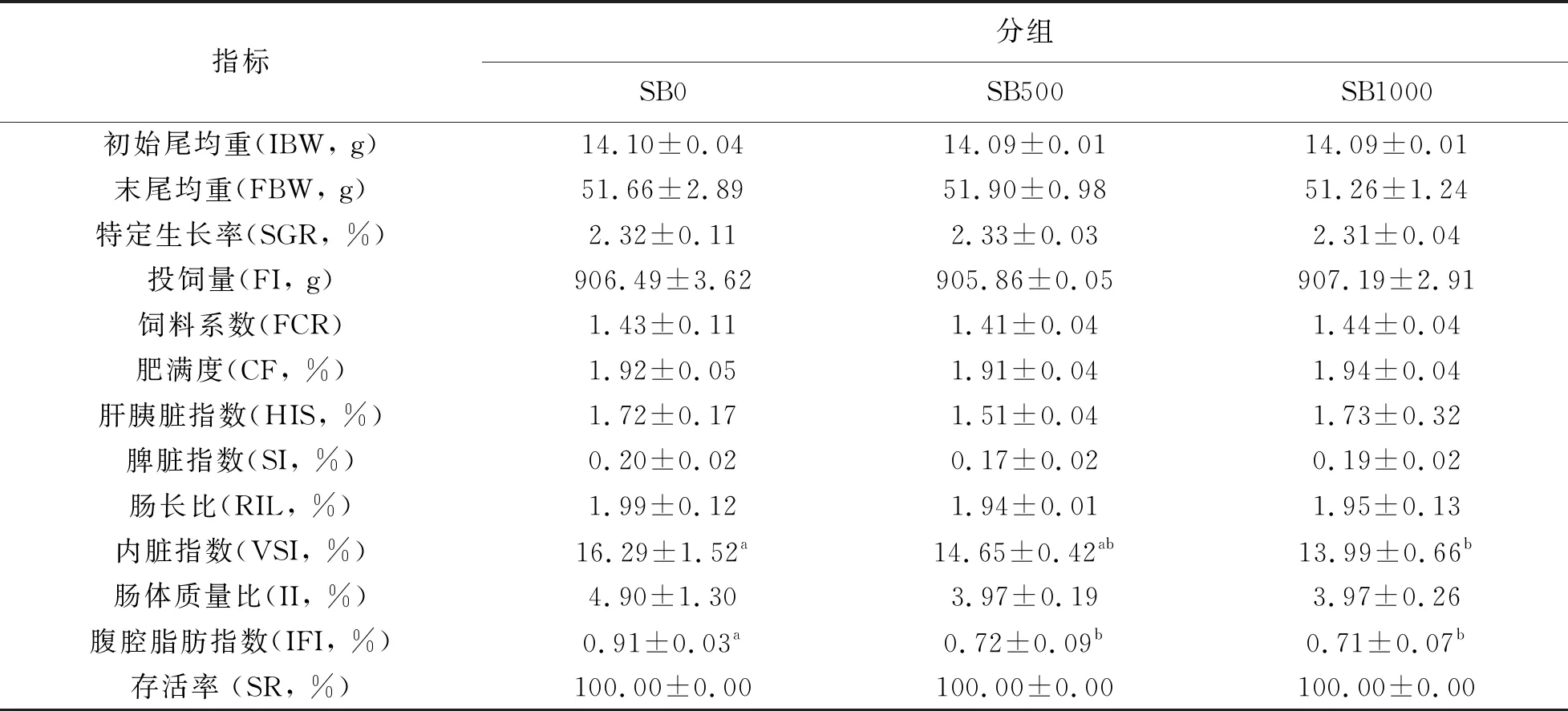
表3 丁酸鹽對草魚生長性能和生物學指標的影響(n=3)
3.2 血清生化指標
血清生化指標檢測表明,SB500組的草魚血清TP和Chol水平顯著低于對照組(P<0.05);SB1000組草魚的血清GLO水平顯著高于SB500組(P<0.05),而大部分草魚血清ALT、AST、ALB、GLU、TG、Chol等,不受丁酸鈉添加水平的影響(P>0.05)。
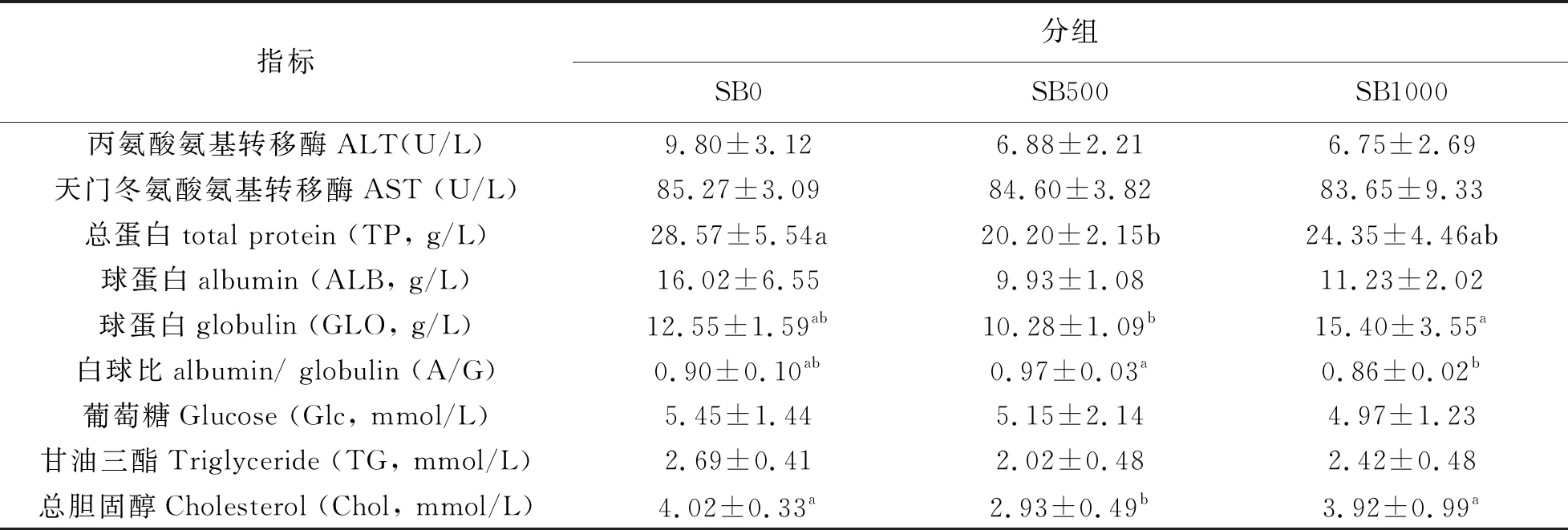
表4 丁酸鹽對草魚血清生化指標的影響(平均值±SD,n=3)
3.3 魚體常規(guī)營養(yǎng)成分測定
在草魚肌肉和肝胰臟中,SB500和SB1000組的粗蛋白和粗脂質(zhì)含量均顯著低于對照組(P<0.05)肌肉粗灰分含量在SB500和SB1000組中顯著升高(P<0.05)。SB1000組小腸粗蛋白含量高于對照組(P<0.05);但丁酸鹽的添加對小腸粗脂質(zhì)含量無明顯降低作用(P> 0.05)。
3.4 組織學觀察
肝胰臟HE染色觀察顯示,與對照組相比,丁酸鹽的添加(SB500及SB1000組)使草魚肝胰臟中的白色空泡狀的脂滴面積和數(shù)量有減小的趨勢。
腸道HE染色觀察顯示,丁酸鹽的添加,草魚腸道絨毛高度呈上升趨勢;通過對草魚腸道絨毛高度的測定,結(jié)果顯示,SB500和SB1000等丁酸鹽的添加水平,均顯著增加了草魚腸道中腸絨毛高度(P<0.05)。
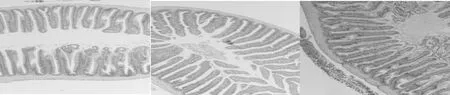
圖2 草魚腸道HE染色觀察(n=3)
3.5 基因相對表達量檢測
丁酸鈉的添加顯著提高了基因tlr22和myd88在肝胰臟中的相對表達量(P<0.05),這種表達量的升高在SB500和SB1000組間無顯著差異(P>0.05)(圖4)。丁酸鈉的添加對基因tlr22和myd88在腸道組織中相對表達量無顯著影響,各組間基因tlr22和myd88mRNA相對表達量無顯著差異(P>0.05)(圖5)。
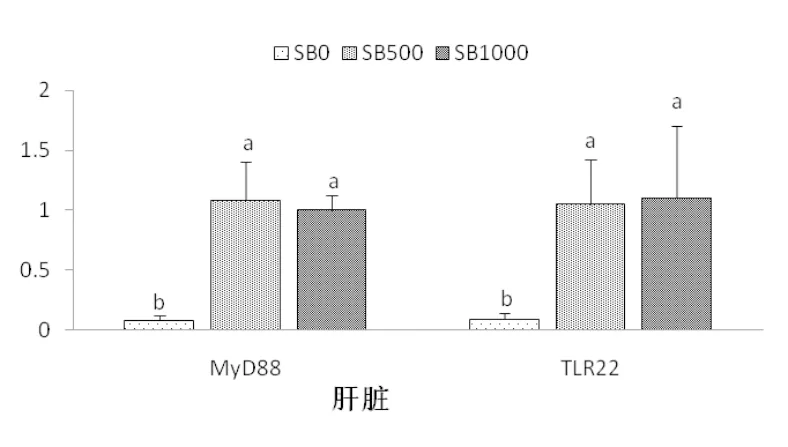
圖4 草魚肝臟基因tlr22和myd88的相對表達量(n=3)
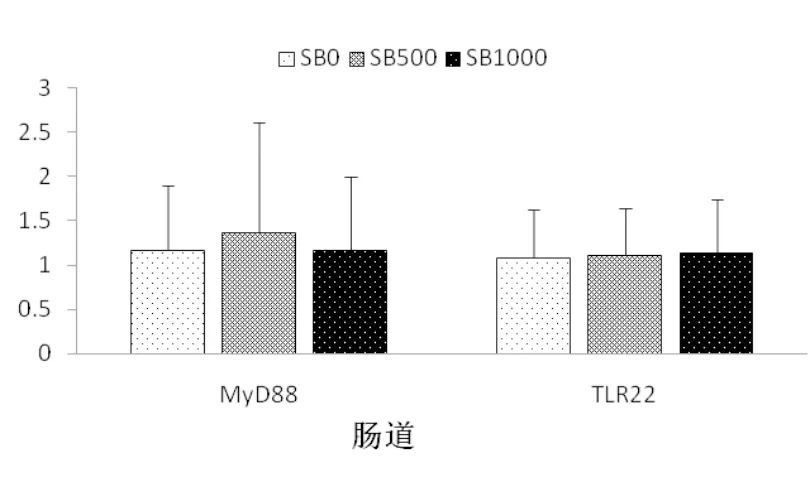
圖5 草魚腸道組織tlr22和myd88的基因相對表達量(n=3)
3.6 腸道菌群
結(jié)果顯示,不同水平丁酸鹽對草魚腸道細菌群落的總序列無顯著影響(P>0.05)。SB1000組腸道菌群ACE指數(shù)和操作分類學單位(OUTs)均顯著高于對照組(P<0.05)。與對照組相比,SB1000組腸道細菌群Simpson多樣性指數(shù)無顯著差異,但該組的Shannon多樣性指數(shù)顯著低于對照組(P<0.05,表6)。

表6 草魚腸道微生物菌群的細菌多樣性指數(shù)(n=3)
對草魚腸道內(nèi)菌群在分類為門上的分析表明,腸道內(nèi)優(yōu)勢菌種為梭桿菌(Fusobacteria)和變形桿菌(Proteobacteria),其中梭桿菌占總菌數(shù)的30%~70%,且丁酸鈉的添加有增加梭桿菌占比的趨勢,但與對照組相對差異并不明顯(P>0.05)。變形桿菌為第二優(yōu)勢菌,占比10%~40%,且該菌受丁酸鈉的影響,表現(xiàn)為SB1000顯著降低了變形桿菌在總菌群中的占比(P<0.05,圖6)。
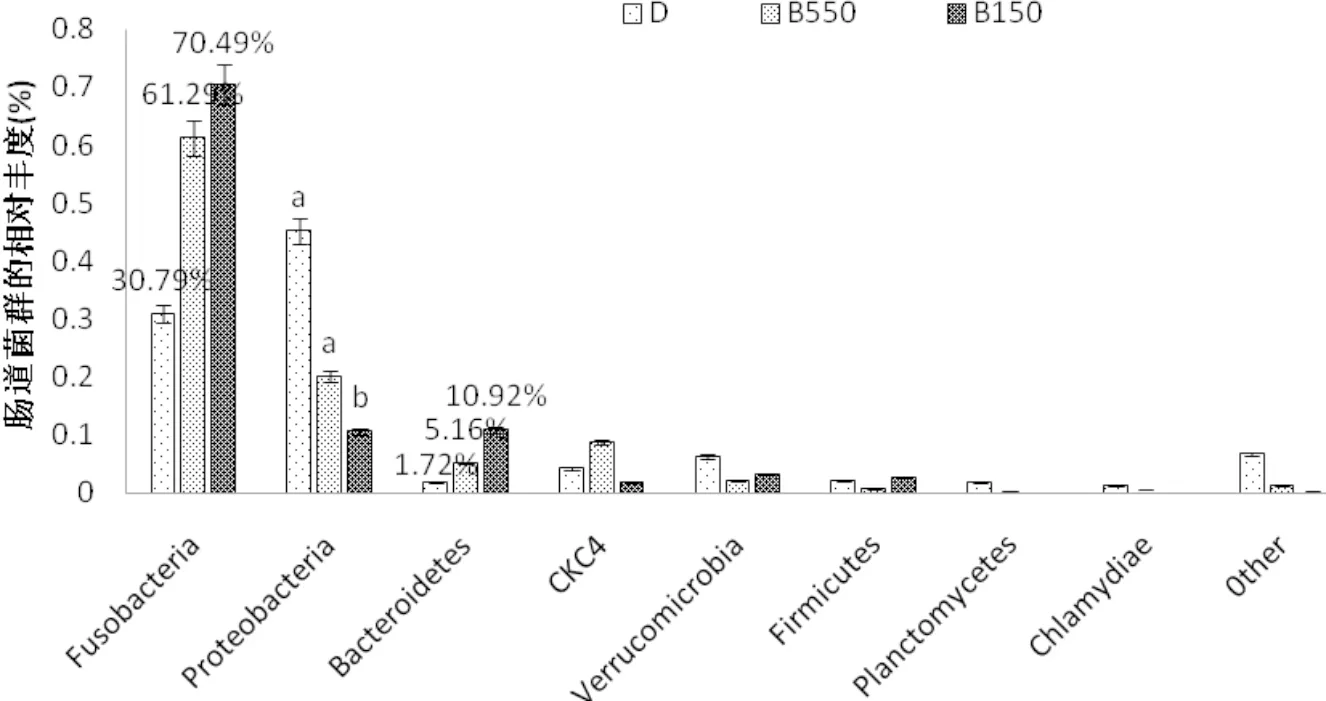
圖6 草魚腸道微生物菌群在門類水平上的相對豐度(n=3)
對草魚腸道內(nèi)菌群在分類為屬上的分析表明,醋酸桿菌cetobacterium為草魚腸道優(yōu)勢菌種,占比為44%~70%,擬桿菌Bacteroidetes為第二優(yōu)勢菌,占比在10%以內(nèi)(圖7)。結(jié)果表明,丁酸鈉的添加對醋酸桿菌與與擬桿菌的占比無顯著影響,但有提升的趨勢。SB0組(44.68%)相比,雖然納入SB500(60.46%)和SB1000(70.24%)后Cetobacterium的相對豐度有增加的趨勢,但差異無統(tǒng)計學意義(P>0.05)。與SB0組(0.39%)相比,補充theSB500組(2.94%)和SB1000組(9.68%)似乎增加了擬桿菌的相對豐度,但增加無統(tǒng)計學意義(P>0.05)。
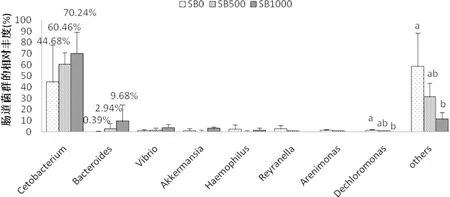
圖7 草魚腸道微生物菌屬水平的相對豐度(n=3)
3 討論
4.1 丁酸鹽對草魚生長的影響
研究發(fā)現(xiàn),飼料中添加2 000 mg·kg-1的丁酸鈉可以提高草魚幼魚的生長、抗氧化能力和腸道吸收能力(Liu et al,2016)。在本試驗研究發(fā)現(xiàn),丁酸鈉在草魚飼料中的添加,各組之間草魚的生長性能沒有顯著差異,這與丁酸鹽在草魚(Liu et al,2016)、大西洋鮭魚(Bjerkeng et al, 1999)和非洲鯰魚(Owen et al.,2006)的研究結(jié)果一致。本試驗結(jié)果與Liu等(2016)在草魚中的報道不相一致的原因,可能與該研究用了較高的丁酸鈉添加水平(2 000 mg kg-1)有關(guān),而本研究中最高的丁酸鈉添加水平僅為1 000 mg kg-1。
4.2 丁酸鹽對草魚脂質(zhì)蓄積的影響
本研究顯示,SB500和SB1000組的腹腔脂肪指數(shù)顯著低于對照組(表2),同時丁酸鈉的添加使草魚具有較低的內(nèi)臟指數(shù),顯示出丁酸鈉具有抑制腹腔脂肪蓄積的作用。這在C57BL/6小鼠的白色脂肪組織中,丁酸鹽也顯示了相似的抑制脂肪蓄積效果(Zhouet al,2016);這可能與短鏈脂肪酸抑制脂肪分解并誘導“褐變”,增加產(chǎn)熱能力,從而導致機體脂肪減少(Sahuri-Arisoylu et al,2016)有關(guān),也可能與短鏈脂肪酸介導的GPR43被激活,而該GPR43的激活抑制了脂肪細胞中的胰島素信號,從而抑制了脂肪組織中脂肪的堆積(Kimura et al, 2013)有關(guān)。
在本試驗結(jié)果顯示,SB500和SB1000的魚的肝胰腺和肌肉中發(fā)現(xiàn)脂質(zhì)積累顯著減少(表5,圖1)。這與許多研究的發(fā)現(xiàn)相一致,如小鼠研究(Jin et al , 2016;Zhou et al, 2016;Sahuri-Arisoylu et al,2016;Kimura et al, 2013;Khan et al, 2016)。Sahuri-Arisoylu等(2016)報道,短鏈脂肪酸在肝臟中減少脂質(zhì)積累,改善肝功能,并提高線粒體效率。短鏈脂肪酸介導的GPR43激活抑制胰島素介導的脂肪積累,從而通過抑制多余能量的積累和促進脂肪消耗來調(diào)節(jié)能量平衡(Kimura et al, 2013)。SB可以對直接和/或間接參與脂質(zhì)代謝的各種調(diào)控基因和/或蛋白質(zhì)進行微調(diào)(Khan et al, 2016)。因此,不同水平的SB可以提高線粒體效率,從而導致肝區(qū)和肌肉的減少。
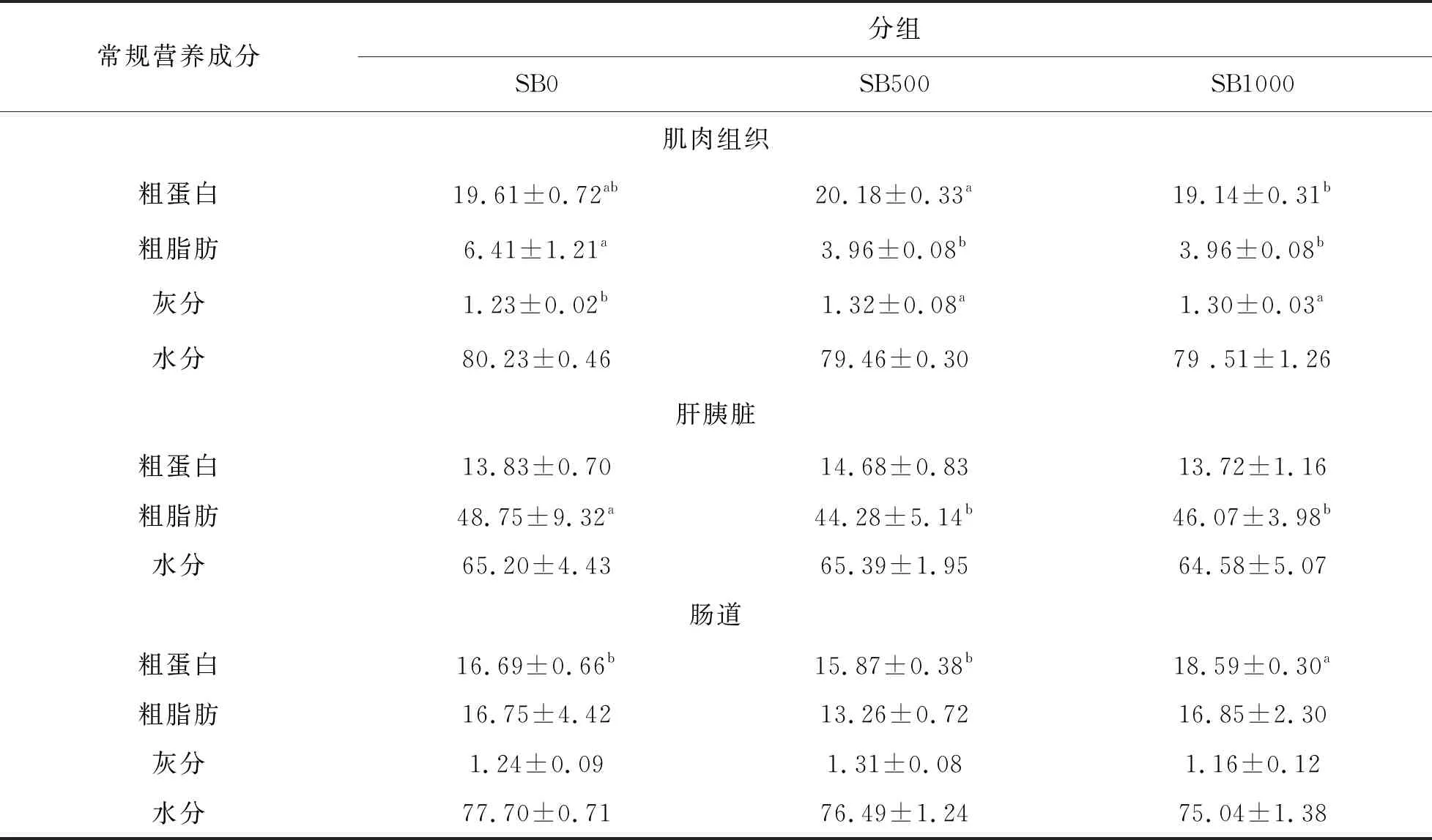
表5 丁酸鹽添加水平對草魚組織中常規(guī)營養(yǎng)成分的影響 (鮮樣,%;n=3)

圖1 草魚肝胰臟HE染色觀察
在水平試驗中,SB1000促進了蛋白在腸內(nèi)的沉積。丁酸鹽似乎增加了幾種必需氨基酸和核苷酸衍生物的可用性。此外,使用丁酸鹽作為燃料可能會降低葡萄糖和氨基酸氧化,從而增強腸細胞的能量供應(Robles et al,2013)。據(jù)我們所知,SB是一種膳食HDAC抑制劑,HDACs調(diào)節(jié)組蛋白乙酰化,一種控制染色質(zhì)結(jié)構(gòu)的表觀遺傳修飾(Mathew et al,2010)。此外,丁酸鹽的添加逆轉(zhuǎn)了極端日糧(Benedito-Palos et al , 2016)喂養(yǎng)的魚的肌肉中蛋白分解(CDH15、CAPN3、PSMA5、PSMB1、UBE2N)的上調(diào)表達(Benedito-Palos et al,2016)。因此,SB似乎是通過下調(diào)肌肉中蛋白分解相關(guān)基因的表達而增加蛋白質(zhì)含量的。然而,在飼喂分級SB水平的草魚中,肌肉的粗蛋白和脂質(zhì)沒有顯著差異(Liu et al,2016)。這些不同的結(jié)果可能是由于魚的大小和養(yǎng)殖條件的差異所致,相關(guān)問題尚需進一步研究。
4.3 丁酸鹽對草魚機體健康
4.3.1 丁酸鹽對草魚血清生化指標的影響 在水平試驗中,血清AST和ALT的活性一般與肝損傷有關(guān),其升高時是肝壞死的標志(Wang et al, 2015)。本研究結(jié)果顯示,丁酸鈉的添加水平對血清谷丙轉(zhuǎn)氨酶和谷草轉(zhuǎn)氨酶活性無顯著影響,顯示出該SB添加水平對草魚肝胰臟健康無負面影響。
研究發(fā)現(xiàn),血清總蛋白和白蛋白反映了機體對營養(yǎng)物質(zhì)的吸收、合成和分解。血清總蛋白和白蛋白的減少是機體營養(yǎng)不良、肝臟疾病和炎癥的反應(Artacho et al, 2007)。血液中GLO含量是保持機體健康和維持正常免疫功能的主要因素,體現(xiàn)了先天免疫機能(Talpur et al,2014)。本試驗顯示,與飼喂對照飼料的魚相比,SB500組的血清總蛋白水平顯著降低,但SB1000組魚體血清總蛋白升高,表明添加較高水平SB對草魚機體健康更有益。本研究結(jié)果顯示,與SB500組相比,SB1000組的草魚血清GLO水平明顯更高,間接說明在先天免疫方面,SB1000飼料可能優(yōu)于SB500飼料。Kurihara等(1985)發(fā)現(xiàn),SB可以誘導成骨細胞MC3T3-E1細胞中GLO的合成。Ahmed等人(2015)報道,在3 g/10 kg日糧中添加SB會導致OreochromisNiloticus的總蛋白、白蛋白和GLO增加。此外,我們發(fā)現(xiàn),A/G(蛋白-球蛋白比值)在SB1000組中明顯低于SB500組。SB1000組的GLO水平可能是所有組中最高的。在研究中,我們發(fā)現(xiàn),在SB500組中,Chol水平顯著降低,提示草魚體內(nèi)脂質(zhì)運輸水平顯著降低。各研究組血清TG無差異。因此,SB的低水平可能會抑制草魚的脂溶或破壞膽固醇的合成能力。此外,SCFAs激活GPR43促進其他組織中多余能量的利用,而不是將多余能量作為脂肪儲存在脂肪組織中,從而維持代謝穩(wěn)態(tài)(Kimura et al,2013)。然而,在類型試驗中,本研究表明,不同類型的SB處理對草魚血清生化指標沒有影響,這間接說明不同類型的SB可能不會影響草魚血清生化指標。在小鼠(Zhou et al, 2016)和裂頭海鱸 (Estensoro I et al,2016)中發(fā)現(xiàn)相似的結(jié)果,血清TG、Chol和Glc無顯著差異。Ju 等(2015)發(fā)現(xiàn)補充不同類型的SB不會影響血清TP、白蛋白和GLO含量。因此,對于SB對血清生化指標的影響似乎沒有共識。
4.3.2 丁酸鹽對草魚腸道健康及微生物菌群的影響 腸是魚類營養(yǎng)物質(zhì)吸收的主要場所,而腸絨毛在這一過程中發(fā)揮著重要作用。本試驗結(jié)果顯示,飼喂不同水平丁酸鈉,草魚中腸的絨毛高度均顯著上升(圖3),說明不同水平SB均能改善草魚的腸道結(jié)構(gòu)。Liu(2014)在鯉魚和Hassanin(2015)在兔中也報道了類似的結(jié)果。SB似乎可以為腸上皮細胞提供能量,促進水和鈉的吸收,從而增加腸上皮細胞的增殖和十二指腸黏膜的生長(Zuo et al,2013;Guilloteau et al, 2010;Huet al,2007)。
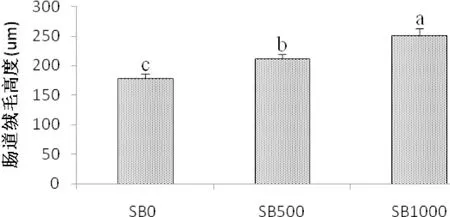
圖3 丁酸鈉添加水平對草魚腸道絨毛高度的影響(μm)
4.3.3 丁酸鹽對草魚免疫機能的影響 骨髓分化因子88(myd88)是一個類似適配器樣的蛋白質(zhì),參與受體(IL-1R)及toll樣受體(tlr22)介導的核激活因子(NF-κB)(Lin et al, 2015)。在本試驗中,myd88和 tlr22的mRNA相對表達量在SB500及SB1000組的肝胰臟中均較高(圖4),表現(xiàn)出丁酸鈉的添加促進了草魚的免疫應答能力,這在歐洲海鱸(Terova et al,2016)及雄性大鼠(Mattace et al, 2013)中也有相似的發(fā)現(xiàn)。組蛋白去乙酰化酶(histone deacetylase,HDAC)是一類蛋白酶,對染色體的結(jié)構(gòu)修飾和基因表達調(diào)控發(fā)揮著重要的作用。Sheikh 等(2016)報道了myd88的乙酰化作用,通過調(diào)節(jié)tlr依賴信號通路到細胞因子基因,進而影響HDAC抑制劑的抗增殖作用。因此推測,丁酸鹽改善草魚肝胰臟免疫應答的功能可能與丁酸鈉是HDAC抑制劑有關(guān)。
研究顯示,丁酸鹽改善草魚免疫應答的功能還具有組織特異性,因為在本試驗中,丁酸鈉的添加對草魚腸道m(xù)yd88和tlr22的相對表達量無顯著影響(圖5),表現(xiàn)出丁酸鈉不影響腸道的免疫應答機能,這在歐洲海鱸(Terova et al,2016)的研究中也有類似發(fā)現(xiàn),但與在鯉魚的研究結(jié)果不一致,在鯉魚的研究中發(fā)現(xiàn),鯉魚飼料中添加丁酸鈉可以顯著改善腸道的免疫應答(Liu et al, 2014)。飼料丁酸鈉對不同魚類腸道組織的免疫反應影響的差別,與魚的種類、魚的大小、養(yǎng)殖條件及丁酸鈉的添加水平差異有關(guān),相關(guān)問題尚需進一步研究。
4.3.4 丁酸鹽對草魚腸道菌群的影響 目前,丁酸鹽對腸道菌群影響的研究主要集中在有益或有害細菌豐度的變化上,而對淡水魚腸道微生物菌群多樣性的影響,研究并不充分。本試驗結(jié)果顯示,SB1000組的Shannon指數(shù)顯著降低(表6),說明SB1000可以降低腸道微生物種群多樣性(Wang et al, 2012)。研究表明,飼料中添加SB1000會降低草魚腸道的pH,從而導致腸道細菌的膜損傷(Hoseinifar et al.,2017),隨著孵育時間的延長,細菌細胞不僅會失去重要離子,還會失去細胞質(zhì)中的溶質(zhì),從而導致菌體的生長受到抑制,嚴重情況下導致細菌生存能力的喪失(Hu et al,2007;Guilloteau et al,2010)。此外,Lu等(2008)還發(fā)現(xiàn),補充1 000 mg·kg-1SB可顯著減少腸道的梭狀芽胞桿菌和大腸桿菌的活菌數(shù)。而在SB1000中ACE指數(shù)明顯低于對照組,說明SB1000可以顯著增加腸道細菌豐度。然而,Liu等(2014)發(fā)現(xiàn)添加300mg kg-1的SB不能提高普通鯉魚腸道菌群的豐度,這種差異可能與丁酸鈉對腸道菌群豐度的影響存在劑量反應效應有關(guān)。在本試驗中,鯨桿菌(Cetobacterium)和擬桿菌(Bacteroides)的相對豐度有隨著丁酸鹽的添加而有增加的趨勢,且該兩種菌在SB1000組的相對豐度最高(表6),表現(xiàn)了丁酸鹽提高了該菌在草魚腸道中的定植,這在雞(Biagi et al, 2007)等動物中也發(fā)現(xiàn)了類似的效果。擬桿菌門中的擬桿菌屬在糖類代謝、營養(yǎng)物質(zhì)吸收、健康的維持和上皮細胞的成熟與維持等方面發(fā)揮著非常重要的作用(Hooper et al,1999; Eckburg et al,2005)。鯨桿菌是梭桿菌中的一種,它在魚的消化過程中發(fā)揮著關(guān)鍵的作用(Lyons et al, 2017)。醋酸是鯨桿菌從蛋白質(zhì)或碳水化合物(Finegold et al, 2003)中得到的主要最終產(chǎn)物。微生物群落的微小差異可能是由于腸黏膜形態(tài)和免疫應答的改變所致(Liu et al, 2014)。此外,變形菌(Proteobacteriain)在SB1000組中的相對豐度顯著低于對照組(圖6),這可能與環(huán)境或宿主因素的變化,如低纖維日糧和急性或慢性炎癥,體內(nèi)平衡被破壞,具有選擇性,并導致腸道內(nèi)變形細菌大量繁殖的生態(tài)失調(diào)(Shin et al, 2015)有關(guān)。綜上所述,在飼料中添加丁酸鈉1 000 mg·kg-1,可抑制草魚脂質(zhì)積累,提高其免疫力,促進其健康。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫(yī)科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫(yī)科大學學報(2021年1期)2021-02-07 01:06:36
現(xiàn)代臨床醫(yī)學(2021年1期)2021-01-26 00:56:02
昆明醫(yī)科大學學報(2020年12期)2021-01-26 00:44:04
中華養(yǎng)生保健(2020年4期)2020-11-16 01:31:40
中西醫(yī)結(jié)合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業(yè)科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫(yī)學院學報(2015年5期)2015-12-05 08:22:29
