ToCV單獨侵染和TYLCV&ToCV復合侵染番茄植株上煙粉虱寄主適應性及寄主植物營養成分含量和防御反應變化
2021-05-11 08:33:14丁天波
昆蟲學報 2021年3期
丁天波, 周 雪, 楊 楠, 楊 煬, 唐 瑤, 褚 棟
(青島農業大學植物醫學學院, 山東省植物病蟲害綜合防控重點實驗室, 山東青島 266109)
煙粉虱Bemisiatabaci是一種至少包含44個隱種的世界性農業害蟲(Kanakala and Ghanim, 2019)。除了通過直接刺吸植物汁液和分泌蜜露導致煤污病等方式危害植物生長以外,該蟲作為媒介昆蟲還可以傳播至少111種植物病毒(Jones, 2003),給多種蔬菜和經濟作物生產造成嚴重的經濟損失。例如,煙粉虱傳播的番茄黃化曲葉病毒(tomato yellow leaf curl virus, TYLCV),自2006年在我國上海首次被報道后,短時間內蔓延至多地,損失慘重(Wuetal., 2006)。近年來,煙粉虱等粉虱傳播的另一種植物病毒——番茄褪綠病毒(tomato chlorosis virus, ToCV),自2012年在我國內陸地區首次被發現(Zhaoetal., 2013)后,迅速蔓延至山東、河北、河南、海南等16個省份和地區(Tangetal., 2017; 魏可可等, 2018; 劉勇等, 2019)。研究發現ToCV在我國的快速流行同煙粉虱種群的擴散有著密不可分的聯系(劉永光等, 2014; 代惠潔等, 2016)。目前,山東、江蘇等多個地區均報道TYLCV和ToCV復合侵染(TYLCV&ToCV復合侵染)發生,給當地番茄種植帶來毀滅性傷害(趙黎明等, 2014; 吳淑華等, 2016; 陳利達等, 2020)。ToCV單獨侵染以及TYLCV&ToCV復合侵染與煙粉虱的互作研究,對于深入理解植物病毒的擴散流行機制具有重要指導價值(Dingetal., 2019)。目前,對于煙粉虱與這些病毒的互作機制知之甚少。
植物病毒可以對媒介昆蟲的寄主適應性產生不同的影響,對其自身的傳播具有重要的意義。Belliure等(2005)發現番茄斑萎病毒(tomato spotted wilt virus, TSWV)有利于其媒介昆蟲西花薊馬Frankliniellaoccidentalis的存活和生長發育;同健康煙草相比,煙草花葉病毒(tobacco mosaic virus, TMV)侵染的煙草對煙蚜Myzuspersicae的種群增長則具有明顯的抑制作用(王佳等, 2017)。諸多研究表明,植物病毒能夠改變寄主植物的營養條件和防御反應,并且是媒介昆蟲寄主適應性變化的重要原因。木薯花葉病毒(cassava mosaic virus, CMV)侵染能夠顯著提高木薯植株韌皮部汁液中氨基酸的含量,其媒介昆蟲煙粉虱的種群增長速率亦明顯加快(Colvinetal., 2006);而西葫蘆感染黃瓜花葉病毒(cucumber mosaic virus, CMV)后,葉片和韌皮部的糖分和氨基酸比例均顯著下降,從而導致蚜蟲的種群適合度降低(Maucketal., 2010, 2014);中國番茄黃化曲葉病毒(tomato yellow leaf curl China virus, TYLCCNV)和其衛星的復合侵染能夠顯著抑制寄主植物的茉莉酸(jasmonic acid, JA)防御途徑,從而對煙粉虱的寄主適應性產生有利影響(Zhangetal., 2012)。
媒介昆蟲是蟲媒植物病毒流行擴散的重要“橋梁”,開展植物病毒對媒介昆蟲的影響及機制研究顯得尤為必要。本研究從田間番茄病毒病發生現狀出發,重點圍繞ToCV單獨侵染和TYLCV&ToCV復合侵染對煙粉虱MED隱種的寄主適應性影響及生理機制展開研究,結合存活率和產卵量兩個參數解析不同番茄病毒侵染對煙粉虱MED隱種的寄主適應性影響;通過測定番茄植株氨基酸和總糖含量比較分析了ToCV單獨侵染和TYLCV&ToCV復合侵染對寄主植物營養狀況的影響;最后,綜合茉莉酸和水楊酸(salicylic acid, SA)途徑關鍵基因表達模式解析不同番茄病毒侵染對寄主植物防御的影響。研究結果將從植物病毒單獨侵染和復合侵染兩個角度進一步理解煙粉虱與病毒的互作機制。
1 材料與方法
1.1 供試昆蟲
供試煙粉虱MED隱種于2012年采自山東省濟南市,并長期以棉花Gossypiumhirsutum(品種:魯棉28)為寄主在人工氣候室中連續飼養。其中飼養溫度為27±1℃,相對濕度為60%±5%,光周期為16L∶8D。
1.2 供試病毒來源
供試ToCV單獨侵染和TYLCV&ToCV復合侵染番茄植株采自于山東省青島市,通過室內飼養煙粉虱對番茄Solanumlycopersicum(品種:中雜9號)植株進行病毒接種,成功后置于人工氣候室中培養,作為供試病毒來源長期保存。ToCV單獨侵染和TYLCV&ToCV復合侵染番茄植株通過以下步驟獲得:將25頭煙粉虱MED隱種雄成蟲置于ToCV單獨侵染或TYLCV&ToCV復合侵染番茄植株上取食獲毒48 h,將其取出,放入微蟲籠中并置于番茄苗(4-5真葉期)從下往上數第2片真葉上,待傳毒48 h后,移出所有煙粉虱,將已接種番茄苗置于網籠中隔離培養,1個月后,根據發病癥狀并結合RT-PCR病毒檢測結果判斷接種番茄的攜毒情況,接種成功的番茄植株作為后續試驗用病毒植株;對照健康番茄植株獲取方式同上,即用煙粉虱MED隱種雄成蟲取食健康番茄48 h后,再置于健康番茄苗相同葉片上48 h。
1.3 煙粉虱MED隱種成蟲在不同番茄植株上的存活率和產卵量比較
參照Su等(2015)的方法,分別取20頭健康初羽化煙粉虱MED隱種雌成蟲放入微蟲籠中,并置于1.2節制備的ToCV單獨侵染、TYLCV&ToCV復合侵染和健康番茄植株從下往上數第4片真葉上,待7 d后,統計在不同番茄植株葉片上煙粉虱的死亡數,計算其存活率,并統計煙粉虱7 d的產卵量。每種處理保證至少3個生物學重復。
1.4 番茄植株中氨基酸含量測定
分別剪取1.2節ToCV單獨侵染、TYLCV&ToCV復合侵染和健康番茄植株新鮮葉片,用液氮進行研磨,稱取0.5 g研磨組織,置于水解管中,并加入16 mL HCl溶液(6 mol/L),真空脫氣30 min后,充氮封管,110℃下水解24 h后,取出冷卻,使用ddH2O轉移并定容至50 mL。準確吸取1 mL水解液,真空脫酸抽干,加入1 mL ddH2O抽干(重復2次),并加入1 mL HCl溶液(0.02 mol/L)充分溶解后,用0.22 μm濾膜進行過濾,將濾液置于高速氨基酸分析儀(日立8900)上進行水解氨基酸和總氨基酸含量測定。每種處理至少保證3個生物學重復。
1.5 番茄植株中總糖含量測定
不同番茄植株中可溶性總糖含量測定參照蒽酮比色法(陳毓荃, 2002; 丁劍等, 2017),并稍作改進。 分別稱取1.2節ToCV單獨侵染、TYLCV&ToCV復合侵染和健康番茄植株新鮮葉片1.0 g,放入三角瓶中,加入25 mL ddH2O后,在沸水浴中煮沸20 min,待自然冷卻后,使用濾膜過濾并用ddH2O定容至100 mL即得樣品提取液。將500 μL樣品提取液、500 μL ddH2O和4 mL蒽酮試劑混勻,在沸水浴中煮沸10 min,待自然冷卻后,取250 μL樣品混合液在多功能酶標儀(SpectraMax? Plus, Molecular Devices)上620 nm處記錄吸光值,對照孔以ddH2O代替樣品提取液。最終結合葡萄糖標準曲線,計算不同番茄樣品的可溶性總糖含量,每種處理至少保證3個生物學重復。
1.6 番茄植株JA和SA信號通路關鍵基因表達模式分析
分別剪取1.2節ToCV單獨侵染、TYLCV&ToCV復合侵染和健康番茄植株從下往上數第4片真葉,經過液氮研磨后,使用TRIzol(Thermo Fisher Scientific)完成總RNA的提取,并對所提取總RNA的質量和濃度進行測定。不同樣品均選取1 μg總RNA,利用PrimeScript RT Reagent Kit (Perfect Real-time)(TaKaRa)并根據說明書完成cDNA第1鏈的合成。每種處理至少保證3個生物學重復。
番茄JA信號通路關鍵基因(FAD7和PIII)及SA信號通路關鍵基因(NPR1和PR1)的實時熒光定量引物參照Li等(2017),根據前人研究選取ubiquitin3(UBI)基因作為內參(Shietal., 2013),具體引物信息見表1。 RT-qPCR反應體系: SYBR Premix ExTaqTMII(TaKaRa)10 μL, cDNA 1 μL, 上下游引物(10 μmol/L)各1 μL, ddH2O 7 μL。反應條件: 95℃ 30 s; 95℃ 5 s, 60℃ 30 s, 40個循環。結合目的基因和內參基因的Ct值,每個基因的相對表達量通過2-△△Ct法計算(Livak and Schmittgen, 2001),每個處理至少保證3個生物學重復和2個技術重復。

表1 實時熒光定量PCR所用引物信息Table 1 Primer sequences used for RT-qPCR
1.7 數據分析
所有實驗數據以平均值±標準誤表示,運用SPSS 16.0軟件對數據進行單因素方差分析(one-way ANOVA),采用Tukey氏檢驗對不同處理數據之間的差異顯著性進行分析(P<0.05)。
2 結果
2.1 煙粉虱MED隱種成蟲在病毒侵染和健康番茄植株上的存活率和產卵量比較
煙粉虱MED隱種以病毒侵染和健康番茄作為寄主時,其存活率之間存在顯著性差異(P<0.05)(圖1: A)。當煙粉虱MED隱種成蟲以健康番茄植株為寄主時,7 d后其存活率為62%,分別是以ToCV單獨侵染和TYLCV&ToCV復合侵染番茄植株為寄主時的1.82和1.88倍(P<0.05)(圖1: A)。當煙粉虱MED隱種成蟲以TYLCV&ToCV復合侵染番茄植株為寄主時,其存活率略低于以ToCV單獨侵染番茄植株為寄主時,但差異不顯著(P>0.05)(圖1: A)。

圖1 不同番茄病毒侵染7 d的番茄植株上的煙粉虱MED隱種成蟲存活率(A)和產卵量(B)Fig. 1 Survival rate (A) and number of eggs laid (B) of Bemisia tabaci MED adultson tomato plants infected with different tomato viruses for 7 dHealthy: 健康番茄植株Healthy tomato plants; ToCV: ToCV單獨侵染番茄植株ToCV-singly infected tomato plants; TYLCV&ToCV: TYLCV&ToCV復合侵染的番茄植株TYLCV&ToCV co-infected tomato plants. 圖中數據為平均值±標準誤;柱上不同字母表示經Tukey氏檢驗后差異顯著(P<0.05)。Data in the figure are means±SE, and different letters above the bars indicate significant differences by Tukey’s test (P<0.05). 下圖同The same for the following figures.
煙粉虱MED隱種成蟲在不同病毒侵染和健康番茄上的產卵量如圖1(B)所示,在ToCV單獨侵染和TYLCV&ToCV復合侵染番茄植株上7 d的產卵量分別為261和226粒,均顯著低于在健康番茄上的產卵量(389.3粒)(P<0.05)。煙粉虱MED隱種成蟲在ToCV單獨侵染和TYLCV&ToCV復合侵染番茄植株上的產卵量差異并不顯著(P>0.05)。
2.2 病毒侵染對番茄植株氨基酸含量的影響
不同病毒侵染和健康番茄植株體內總氨基酸含量檢測結果表明(圖2: A),經TYLCV&ToCV復合侵染后,番茄植株中總氨基酸含量明顯低于健康番茄植株中總氨基酸含量,并且顯著低于ToCV單獨侵染后番茄植株中的總氨基酸含量(P<0.05)。ToCV單獨侵染和健康番茄植株中的總氨基酸含量則無顯著性差異(P>0.05)。
進一步對不同病毒侵染和健康番茄植株中17種水解氨基酸含量進行測定和比較分析,結果發現(圖2: B),在ToCV單獨侵染、TYLCV&ToCV復合侵染和健康番茄植株中,谷氨酸(Glu)、天冬氨酸(Asp)和亮氨酸(Leu)含量均較高,半胱氨酸(Cys)、蛋氨酸(Met)和組氨酸(His)含量均較低。TYLCV&ToCV復合侵染番茄植株中絲氨酸(Ser)、甘氨酸(Gly)、丙氨酸(Ala)、纈氨酸(Val)、亮氨酸(Leu)和苯丙氨酸(Phe)的比例顯著降低(P<0.05),14種氨基酸[天冬氨酸(Asp)、蘇氨酸(Thr)、絲氨酸(Ser)、甘氨酸(Gly)、丙氨酸(Ala)、纈氨酸(Val)、異亮氨酸(Ile)、亮氨酸(Leu)、酪氨酸(Tyr)、苯丙氨酸(Phe)、賴氨酸(Lys)、組氨酸(His)、精氨酸(Arg)、脯氨酸(Pro)]含量均顯著低于ToCV單獨侵染番茄植株(P<0.05);其他3種氨基酸[谷氨酸(Glu)、半胱氨酸(Cys)、蛋氨酸(Met)]含量在ToCV單獨侵染和TYLCV&ToCV復合侵染番茄植株中則沒有顯著性差異(P>0.05)。

圖2 不同番茄病毒侵染對番茄植株中總氨基酸(A)和水解氨基酸(B)含量的影響Fig. 2 Effects of the infection of different tomato viruses on the contents of total amino acids (A)and hydrolyzed amino acids (B) in tomato plants
2.3 病毒侵染對番茄植株總糖含量的影響
通過對不同病毒侵染和健康番茄植株內總糖含量進行測定和比較,結果表明(圖3),TYLCV&ToCV復合侵染番茄植株中總糖含量低于健康番茄植株和ToCV單獨侵染番茄植株總糖含量,并且同ToCV單獨侵染番茄植株之間差異顯著(P<0.05)。
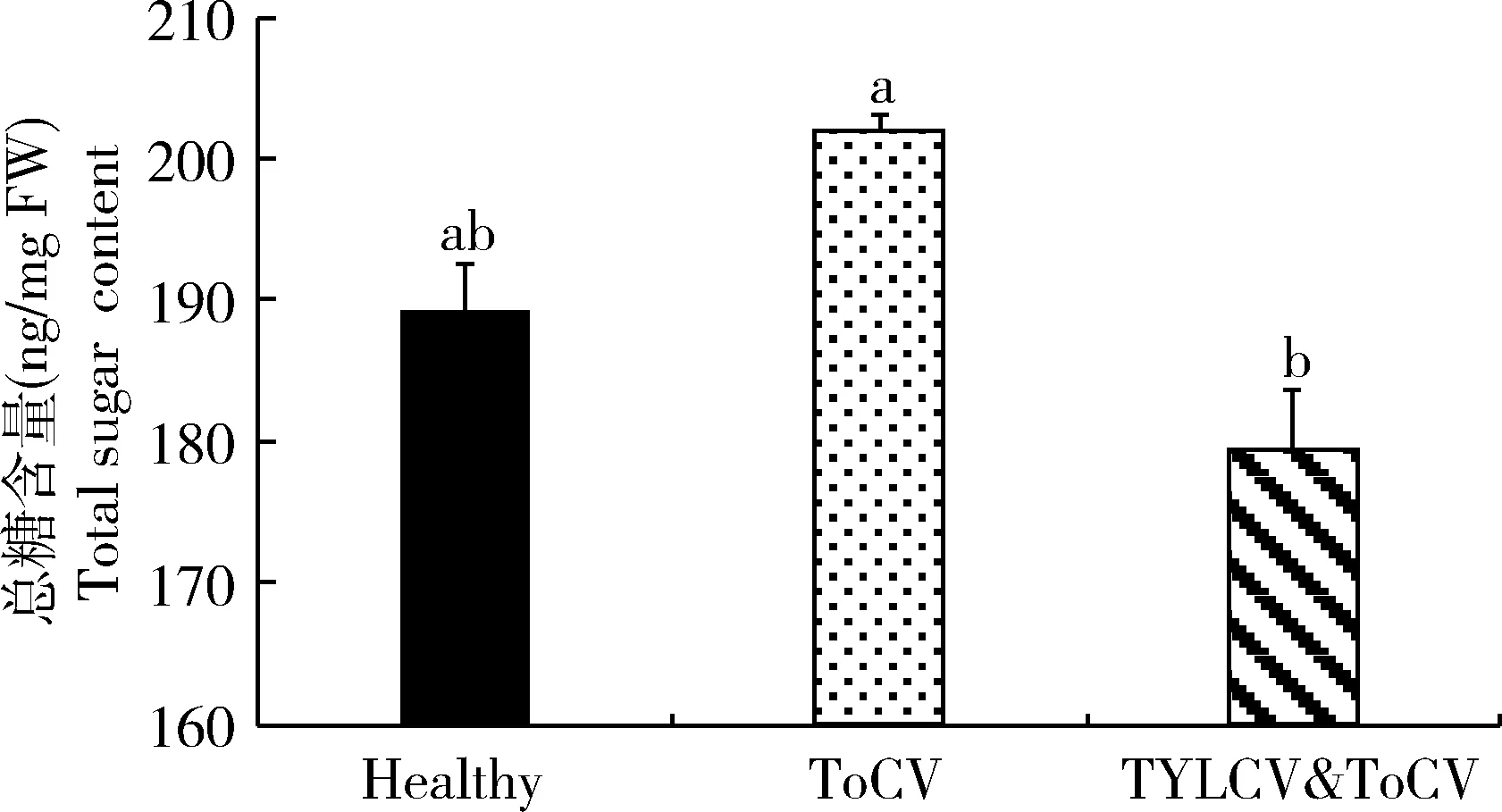
圖3 不同番茄病毒侵染對番茄植株中總糖含量的影響Fig. 3 Effects of the infection of different tomato viruses onthe total sugar content in tomato plants
2.4 病毒侵染條件下番茄植株JA信號通路關鍵基因響應模式比較
本研究選取番茄JA信號通路兩個關鍵基因FAD7和PIII,對二者在不同病毒侵染和健康番茄中的表達模式進行比較研究。相對于健康植株,ToCV單獨侵染和TYLCV&ToCV復合侵番茄植株中FAD7基因的mRNA表達水平均顯著降低(P<0.05),分別是健康番茄植株中的0.86和0.46倍(圖4: A)。TYLCV&ToCV復合侵染番茄植株中FAD7基因的表達量顯著低于ToCV單獨侵染番茄植株中該基因的表達量(P<0.05),前者是后者的0.53倍。同樣,在ToCV單獨侵染和TYLCV&ToCV復合侵染番茄植株中PIII基因的表達水平顯著低于健康番茄植株中的(P<0.05),分別是健康番茄植株中表達量的0.31和0.05倍(圖4: B)。并且TYLCV&ToCV復合侵染番茄植株中PIII基因的表達量最低,是ToCV單獨侵染番茄植株中的0.16倍,差異顯著(P<0.05)。

圖4 不同番茄病毒侵染對番茄植株中JA信號通路關鍵基因FAD7(A)和PI II(B)表達模式的影響Fig. 4 Effects of the infection of different tomato viruses on the expression profiles of the key genes FAD7 (A)and PI II (B) involved in JA signal pathway in tomato plants
2.5 病毒侵染條件下番茄植株SA信號通路關鍵基因響應模式比較
本研究選取番茄SA信號通路兩個關鍵基因NPR1和PR1,對二者在不同病毒侵染和健康番茄中的表達模式進行比較研究。結果發現,ToCV單獨侵染和TYLCV&ToCV復合侵染后番茄植株中NPR1基因的mRNA表達水平均有上升趨勢,其中TYLCV&ToCV復合侵染番茄植株中NPR1基因表達量顯著高于健康和ToCV單獨侵染番茄植株中的(P<0.05)(圖5: A)。ToCV單獨侵染和TYLCV&ToCV復合侵染番茄植株中PR1基因表達水平均高于健康番茄植株中的,但被ToCV單獨侵染后,番茄植株中PR1基因表達水平顯著高于TYLCV&ToCV復合侵染番茄植株中的(P<0.05),且前者是后者的2.78倍(圖5: B)。

圖5 不同番茄病毒侵染對番茄植株SA途徑關鍵基因NPR1 (A)和PR1 (B)表達模式的影響Fig. 5 Effects of the infection of different viruses on the expression profiles of the keygenes NPR1 (A) and PR1 (B) involved in SA signal pathway in tomato plants
3 討論
本研究發現,番茄植株經ToCV單獨侵染后,煙粉虱MED隱種成蟲在其上的存活率和產卵量均顯著低于在健康番茄植株上的(圖1),與Li等(2018)結合兩性生命表研究的研究結果是一致的,該研究以ToCV單獨侵染番茄植株作為寄主的煙粉虱MED隱種成蟲的繁殖力、壽命等均顯著降低。本研究還發現取食TYLCV&ToCV復合侵染番茄植株的煙粉虱MED隱種成蟲的存活率和產卵量也明顯降低,說明不論是ToCV單獨侵染,還是TYLCV&ToCV復合侵染,均能夠對其媒介昆蟲煙粉虱的寄主適應性帶來不利的影響。前人研究發現相對于ToCV侵染番茄植株,攜帶ToCV煙粉虱MED隱種則更傾向于選擇健康番茄植株(Shietal., 2018; Weietal., 2019);本研究發現ToCV侵染植株不利于煙粉虱生存,可能與攜毒煙粉虱寄主選擇性相關。
植物病毒能夠引起植物營養狀況(氨基酸、糖分含量等)的改變,從而影響媒介昆蟲的寄主適應性(Bluaetal., 1994; Colvinetal., 2006)。例如取食TYLCV侵染植株的煙粉虱體重、產卵量和存活率均顯著升高,可能與TYLCV侵染番茄韌皮部汁液中氨基酸和糖分含量升高相關(Suetal., 2015)。二斑葉螨Tetranychusurticae對TSWV侵染番茄植株具有明顯的寄主偏好性,并且其產卵量顯著升高,可能與TSWV侵染番茄植株中氨基酸含量升高相關(Nachappaetal., 2013)。而本研究結果顯示,TYLCV&ToCV復合侵染番茄植株總氨基酸含量明顯低于健康番茄植株,其中3種昆蟲必需氨基酸[纈氨酸(Val)、亮氨酸(Leu)和苯丙氨酸(Phe)]的比例顯著降低(圖2: B);TYLCV&ToCV復合侵染番茄植株總氨基酸含量和14種水解氨基酸的比例均顯著低于ToCV單獨侵染植株(圖2)。此外,TYLCV&ToCV復合侵染番茄植株中總糖含量低于ToCV單獨侵染和健康植株(圖3)。這些結果也表明TYLCV&ToCV復合侵染比ToCV單獨侵染對番茄植株營養狀況更具不利影響。
植物病毒亦能夠影響寄主植物體內的防御響應,導致昆蟲寄主適應性的改變(Zhangetal., 2012)。為進一步探究媒介昆蟲煙粉虱在ToCV單獨侵染或TYLCV&ToCV復合侵染番茄植株上生物學特性的差異,本研究中運用RT-qPCR技術對不同病毒侵染條件下番茄JA和SA信號通路關鍵基因的響應模式進行了分析。結果顯示,ToCV單獨侵染和TYLCV&ToCV復合侵染后,番茄JA信號通路兩個關鍵基因FAD7和PIII的表達量均顯著下調,并且其在復合侵染番茄植株體內表達水平最低(圖4)。前人在病毒侵染植株上也發現了JA途徑被抑制的現象(Lewseyetal., 2010; Shietal., 2014),本研究同時也發現番茄SA信號通路兩個關鍵基因NPR1和PR1則在一定程度上被不同番茄病毒誘導表達,NPR1基因在TYLCV&ToCV復合侵染條件下上調幅度最大,而PR1基因的表達水平則在ToCV單獨侵染番茄中最高(圖5)。同樣,前人在煙草花葉病毒(tobacco mosaic virus, TMV)或TYLCV侵染番茄中也發現SA含量上升的現象(Thaleretal., 2010; Shietal., 2013),煙粉虱MEAM1隱種取食SA處理番茄植株后,其存活率和產卵量均顯著降低。因此,可以推斷本研究中ToCV單獨侵染和TYLCV&ToCV復合侵染誘導的SA信號通路正響應在一定程度上降低了煙粉虱MED隱種的寄主適應性。
前人研究發現,TYLCV侵染有利于煙粉虱MED隱種種群上升,可能由于TYLCV侵染可使植物營養條件朝有利于煙粉虱生存的方向發展,并且TYLCV侵染顯著抑制了植物防御反應,諸如植物過氧化物酶和超氧化物歧化酶等的合成(Suetal., 2015)。而在本研究中,TYLCV&ToCV的復合侵染植株則不利于煙粉虱MED隱種種群上升,植株營養明顯下降,這可能是由于TYLCV&ToCV復合侵染與植物的互作關系不同于病毒單獨侵染與植物的互作關系。該方面的機制尚需進一步的研究。
