基于酵母表面展示技術(shù)的耐熱木聚糖酶全細胞催化劑的構(gòu)建及酶學(xué)性質(zhì)研究
2021-05-09 06:02:10鄧香連劉清怡熊海容
湖北農(nóng)業(yè)科學(xué) 2021年7期
關(guān)鍵詞:催化劑
鄧香連,劉清怡,熊海容,張 莉
(中南民族大學(xué)生命科學(xué)學(xué)院,武漢430074)
木聚糖酶(Xylanase)是可將木聚糖降解為低聚木糖和少量木糖的一類酶,主要包括內(nèi)切木聚糖酶、外切木聚糖酶和木糖苷酶,其中內(nèi)切木聚糖酶能水解木聚糖主鏈中的β-1,4糖苷鍵生成木二糖、木三糖等寡糖,是降解木聚糖最主要的酶[1,2]。木聚糖酶來源廣泛,可從動物、植物或微生物中獲得,目前研究較多的是來源于細菌或真菌的木聚糖酶[3]。已有多種不同來源的木聚糖酶在多個宿主中成功進行了異源表達,如大腸桿菌(Escherichia coli)、枯草芽孢桿菌(Bacillus subtilis)、畢赤酵母等,不同程度地提高了木聚糖酶的表達量[4,5]。
木聚糖酶是一種重要的工業(yè)酶制劑,在飼料、造紙、食品、紡織等領(lǐng)域具有廣闊的應(yīng)用前景[6]。但在具體應(yīng)用過程中環(huán)境條件要求較苛刻,如飼料生產(chǎn)制粒工藝中的調(diào)質(zhì)、制粒等過程需要70℃以上的高溫處理,使添加的酶活性損失很大,降低了使用效率[7];麻類紡織中苧麻脫膠、造紙中紙漿溶解和漂白等工藝需要強酸、強堿等處理,也會使酶的活性大大降低[8],因此提高酶在高溫、酸性、堿性、含有機溶劑等條件下的穩(wěn)定性受到越來越多的關(guān)注。
微生物表面展示技術(shù)近年來發(fā)展迅速且應(yīng)用廣泛,利用微生物細胞展示酶作為催化劑(也稱全細胞催化劑)不僅具有固定化酶的特性,且可省去繁瑣的分離純化操作,制備方法簡單,易于回收和再生利用[9]。多項研究顯示全細胞催化劑較游離酶表現(xiàn)出更好的穩(wěn)定性,Yuzbasheva等[10]在解脂耶氏酵母中獲得脂肪酶Lip2p的全細胞催化劑,在50℃處理5 h后殘留約83%的酶活力,而游離酶在50℃處理1 h酶活力幾乎降為零,此外將全細胞催化劑和游離酶用EDTA、SDS、DMSO等試劑處理24 h,全細胞催化劑殘余的酶活力均高于游離酶;Wang等[11]在釀酒酵母中展示表達葡萄糖氧化酶GOx,發(fā)現(xiàn)GOx全細胞催化劑較游離酶在30~60℃具有更好的熱穩(wěn)定性。
本研究利用酵母表面展示技術(shù)將實驗室前期篩選獲得的耐熱木聚糖酶DSB[12]展示表達在畢赤酵母細胞表面獲得DSB全細胞催化劑,并對全細胞催化劑和游離酶的酶學(xué)性質(zhì)進行比較分析,為DSB全細胞催化劑的工業(yè)應(yīng)用提供一定的理論依據(jù),也為獲得穩(wěn)定性更好的酶制劑提供一定的理論基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 菌種與質(zhì)粒含木聚糖酶DSB基因的重組質(zhì)粒pPICZαA/DSB由實驗室構(gòu)建并保藏。克隆宿主大腸桿菌Top10、表達宿主巴斯德畢赤酵母X33均由實驗室保藏。
1.1.2 主要試劑PrimeSTAR Max DNA聚合酶、限制性內(nèi)切酶、T4 DNA連接酶、DNA Marker等購于大連寶生物工程有限公司;DNA凝膠回收試劑盒、PCR清潔試劑盒、質(zhì)粒小量提取試劑盒等購于上海Axy?Prep公司;底物木聚糖為實驗室自提樺木木聚糖[13];Tryptone和Yeast Extract購于Oxoid公司;山梨醇、瓊脂、無氨基酵母氮源YNB、生物素均購于Biosharp公司;DSB游離酶由實驗室制備;其他試劑均為國產(chǎn)分析純。
1.1.3 培養(yǎng)基大腸桿菌培養(yǎng)基LB、大腸桿菌抗性篩選平板低鹽LB(含抗生素Zeocin,終濃度25μg/mL)、畢赤酵母培養(yǎng)基YPD、畢赤酵母抗性篩選平板YPDSZ(含抗生素Zeocin,終濃度100μg/mL)、畢赤酵母生長/誘導(dǎo)培養(yǎng)基BMGY/BMMY等配方見Invitrogen公司畢赤酵母操作手冊。
1.2 方法
1.2.1 展示表達DSB重組質(zhì)粒的構(gòu)建以畢赤酵母細胞壁蛋白Gcw61(Accession in NCBI XP_002 494332)[14]為錨定蛋白,去除自身信號肽后設(shè)計引物(表1),并在5'端和3'端分別引入KpnⅠ和NotⅠ位點(下劃線標記)。以畢赤酵母X33基因組為模板進行PCR擴增,對獲得的GCW61基因片段與質(zhì)粒pPICZαA/DSB同時進行雙酶切,然后進行連接、轉(zhuǎn)化和篩選后獲得展示表達DSB重組質(zhì)粒pPICZαA/DSB-GCW61,并送至武漢擎科生物技術(shù)有限公司進行測序驗證。
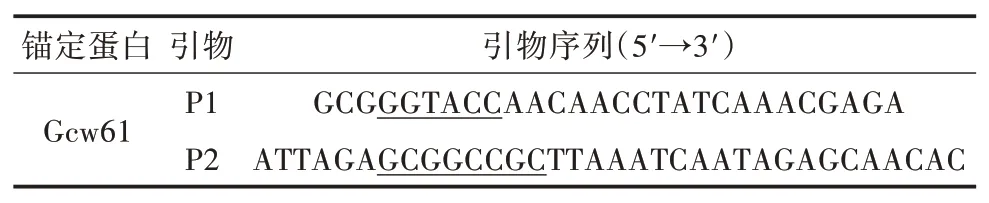
表1 PCR擴增引物序列
1.2.2 展示表達DSB重組菌的構(gòu)建及誘導(dǎo)表達對pPICZαA/DSB-GCW61使用SacI進行線性化,然后采用電轉(zhuǎn)化法轉(zhuǎn)化宿主菌畢赤酵母X33感受態(tài)細胞,涂布于YPDSZ抗性篩選平板上,于28℃培養(yǎng)2~3 d,使用AOX1通用引物對轉(zhuǎn)化子進行菌落PCR鑒定獲得展示表達DSB重組菌X33/DSB-GCW61。將該重組菌接種于BMGY培養(yǎng)基中,28℃、250 r/min過夜培養(yǎng)至OD600達2~6,取適量菌體重懸于BMMY培養(yǎng)基中,控制起始OD600約為1,28℃、250 r/min連續(xù)發(fā)酵168 h,每隔24 h取樣,并補加1%的甲醇。以畢赤酵母X33作為陰性對照,所有試驗均重復(fù)3次。
1.2.3 重組菌的免疫熒光顯微鏡分析發(fā)酵結(jié)束后取1 mL菌懸液,離心去除上清,用磷酸鹽緩沖溶液(PBS,pH 7.4)洗滌菌體3次,重懸至含10 mg/mL BSA的PBS中,調(diào)整樣品OD600值約為5。取200μL樣品加入鼠源抗6×His標簽單抗(1∶200稀釋),室溫下低速振蕩孵育2 h。孵育結(jié)束后用PBS洗3次,重懸至含10 mg/mL BSA的PBS中,加入Alexa Flour 488標記羊抗鼠IgG抗體(1∶200稀釋)于避光處室溫低速振蕩孵育1 h。孵育結(jié)束后用PBS洗3次,使用激光共聚焦顯微鏡觀察,激發(fā)波長為488 nm,以畢赤酵母X33細胞作為陰性對照。
1.2.4 重組菌展示表達DSB酶活力的測定對X33/DSB-GCW61進行誘導(dǎo)表達,每隔24 h取樣后離心去除上清,用50 mmol/L pH 6.0磷酸氫二鈉-檸檬酸緩沖溶液洗滌菌體3次,然后用等量緩沖溶液重懸后獲得DSB全細胞催化劑樣品。以0.5%木聚糖為底物,采用DNS法[15]測定酶活力,同時測定菌懸液的OD600值,根據(jù)1OD600=0.245 g/L[9]計算出最終酶活力。酶活力單位定義為在最適溫度和pH條件下,1 min內(nèi)水解0.5%木聚糖底物產(chǎn)生1μmol木糖所需要的全細胞催化劑的量。以畢赤酵母X33細胞作為對照,所有試驗均重復(fù)3次。
1.2.5 DSB全細胞催化劑和游離酶的酶學(xué)性質(zhì)比較分析
1)溫度對全細胞催化劑和游離酶酶活力的影響。在pH 6.5條件下,分別在40、50、60、65、70、75、80℃測定全細胞催化劑和游離酶的酶活力,設(shè)2種酶的最高酶活力為100%,計算不同溫度條件下2種酶的相對酶活力。取用pH 6.5檸檬酸緩沖液適當稀釋后的2種酶置于70℃水浴處理2.5 h,每隔30 min測定2種酶的酶活力,設(shè)未處理的2種酶的酶活力為100%,計算在70℃條件下處理不同時間2種酶的殘余酶活力。
2)pH對全細胞催化劑和游離酶酶活力的影響。分別配制pH 4.0~8.0 50 mmol/L的Na2HPO4-檸檬酸緩沖液體系,以上述緩沖液分別配制0.5%的木聚糖底物,且以對應(yīng)pH緩沖液稀釋2種酶,分別使用上述底物測定2種酶的酶活力,反應(yīng)溫度均為65℃,設(shè)2種酶的最高酶活力為100%,計算不同pH條件下2種酶的相對酶活力。分別用pH 3.0和pH 6.0的緩沖液適當稀釋2種酶后,放置40℃水浴處理2.5 h,每隔30 min分別測定2種酶的酶活力,設(shè)未處理的2種酶的酶活力為100%,計算不同pH條件下處理不同時間2種酶的殘余酶活力。
3)高濃度鹽對全細胞催化劑和游離酶酶活力的影響。在2種酶的酶活反應(yīng)體系中分別加入1%、3%、5%、10%、15%的NaCl或KCl,然后在65℃、pH 6.5條件下測定2種酶的酶活力,以不加鹽的2種酶的酶活力為100%,計算不同鹽濃度條件下2種酶的相對酶活力。
4)有機溶劑對DSB全細胞催化劑和游離酶酶活力的影響。分別配制0~30 %的甲醇、乙醇、正丁醇、丙三醇溶液,用上述不同濃度的有機試劑稀釋2種酶并在室溫孵育30 min后,在65℃、pH 6.5條件下測定2種酶的酶活力,設(shè)未用有機試劑處理的2種酶的酶活力為100%,計算不同濃度有機試劑處理后2種酶的相對酶活力。
5)金屬離子和化學(xué)試劑對DSB全細胞催化劑和游離酶酶活力的影響。在2種酶的酶活反應(yīng)體系中加入適量不同金屬離子和化學(xué)試劑溶液,使其終濃度為1.0 mmol/L,然后在65℃、pH 6.5條件下測定2種酶的酶活力,設(shè)未做處理的2種酶的酶活力為100%,計算不同金屬離子和化學(xué)試劑處理后2種酶的相對酶活力。以上所有試驗均重復(fù)3次。
2 結(jié)果與分析
2.1 展示表達DSB重組菌的構(gòu)建
以畢赤酵母X33基因組為模板,用引物P1/P2進行PCR擴增獲得錨定蛋白Gcw61的基因片段(144 bp),將其與質(zhì)粒pPICZαA/DSB同時使用KpnⅠ和NotⅠ雙酶切,然后進行連接、轉(zhuǎn)化和篩選后獲得展示表達DSB的重組質(zhì)粒pPICZαA/DSB-GCW61,經(jīng)武漢擎科生物技術(shù)有限公司測序驗證載體構(gòu)建成功(圖1)。隨后將該重組質(zhì)粒轉(zhuǎn)入畢赤酵母X33,篩選獲得展示表達DSB的重組菌X33/DSB-GCW61。
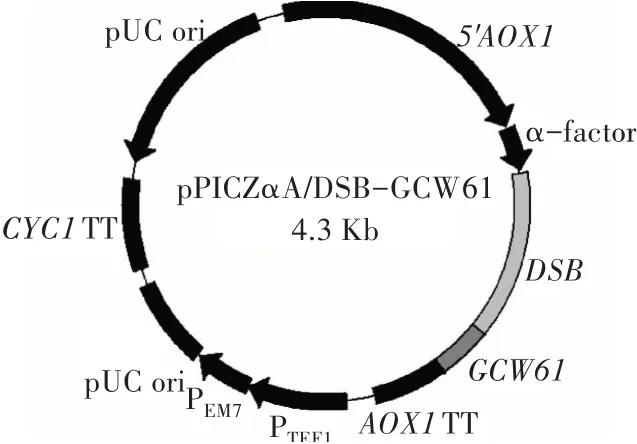
圖1 重組質(zhì)粒pPICZαA/DSB-GCW61
2.2 重組菌熒光的觀察
木聚糖酶DSB的N端帶有6×His標簽,取誘導(dǎo)表達168 h的重組菌使用抗體處理后在激光共聚焦顯微鏡下觀察(激發(fā)波長488 nm),結(jié)果如圖2所示。由圖2可見,重組菌細胞發(fā)出綠色熒光,表明錨定蛋白Gcw61成功將DSB展示表達在畢赤酵母細胞表面。
2.3 展示表達DSB重組菌的酶活分析
對重組菌X33/DSB-GCW61在搖瓶水平誘導(dǎo)表達168 h,每隔24 h取樣,檢測木聚糖酶全細胞催化劑的酶活力,結(jié)果如圖3所示。從圖3可以看出,隨著發(fā)酵時間的增加,DSB全細胞催化劑的酶活力會持續(xù)增長,且在72~144 h增長較快,144 h后酶活力趨于穩(wěn)定,168 h時酶活力最大,達13 510 U/g。
2.4 DSB全細胞催化劑與游離酶酶學(xué)性質(zhì)的比較分析
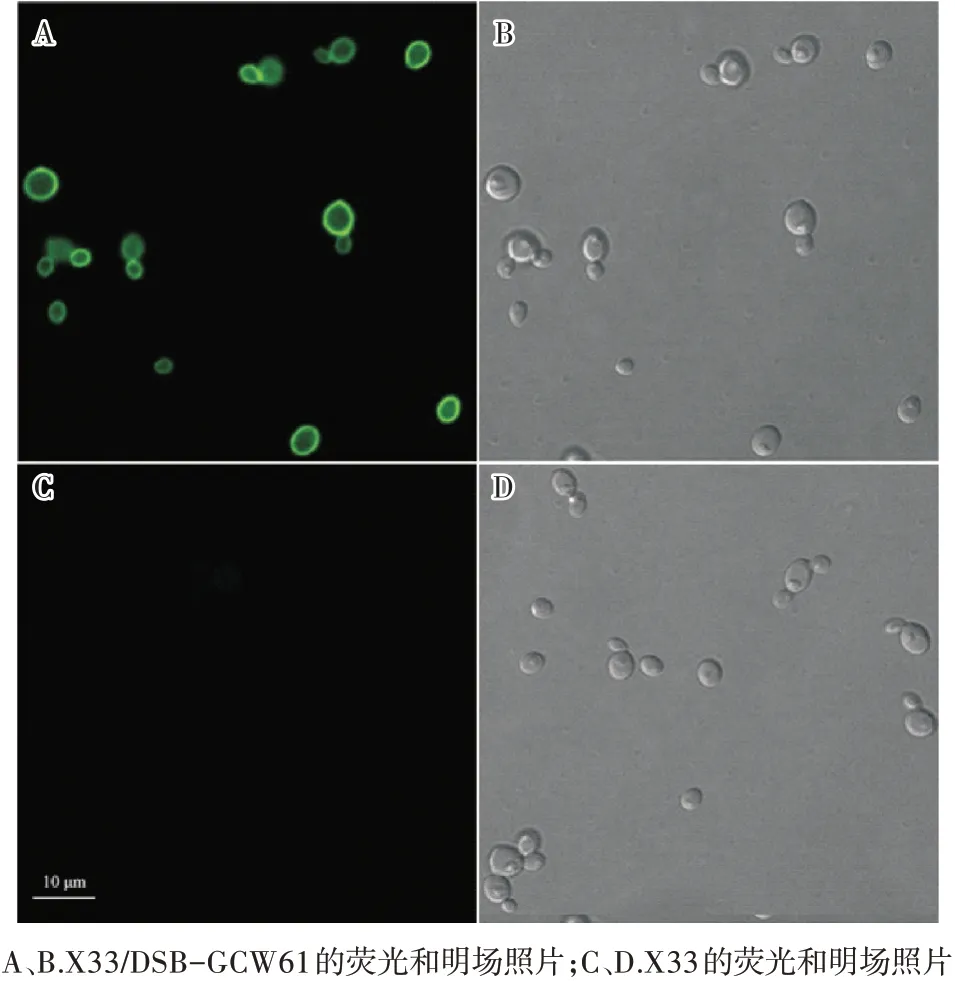
圖2 重組菌的熒光顯微鏡觀察
2.4.1 溫度對2種酶酶活力的影響測定不同溫度下DSB全細胞催化劑和DSB游離酶的酶活力,從圖4(A)可以看出,2種酶的最適反應(yīng)溫度均為65℃,屬于高溫酶,且2種酶在50~75℃都能保持50%以上的相對酶活力,但DSB全細胞催化劑的相對酶活力均高于DSB游離酶。將2種酶在70℃條件下保溫處理,監(jiān)測2種酶酶活力的變化,圖4(B)的結(jié)果顯示隨著處理時間的增加,2種酶的酶活力均逐漸降低,但DSB全細胞催化劑的相對酶活力始終顯著高于DSB游離酶,在處理120 min時,DSB全細胞催化劑的相對酶活力為50%,而DSB游離酶的相對酶活力已降至30%。工業(yè)應(yīng)用中很多工藝需高溫處理,如飼料加工制粒過程溫度會高達70℃以上,DSB全細胞催化劑比DSB游離酶具有更好的耐熱性,這一特性使其在飼料行業(yè)具有較好的應(yīng)用前景。
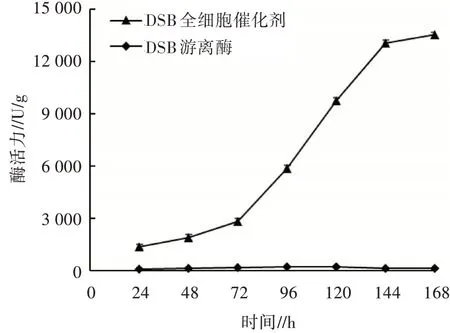
圖3 2株菌的酶活曲線
2.4.2 pH對2種酶酶活力的影響測定不用pH條件下DSB全細胞催化劑和DSB游離酶的酶活力,結(jié)果見圖5(A),2種酶的最適反應(yīng)pH均為6.5,在pH 5.0~7.5均可保持60%以上的相對酶活力,但DSB全細胞催化劑的相對酶活力略高于DSB游離酶。將2種酶在pH 3.0和pH 6.0酸性條件下處理,監(jiān)測2種酶酶活力的變化,從圖5(B)可以看出,在pH 6.0、40℃條件下處理150 min,DSB全細胞催化劑的酶活力無明顯下降,而DSB游離酶的酶活力下降約7%;而在pH 3.0、40℃條件下處理90 min,2種酶可以保持80%以上的相對酶活力,處理150 min也可保持60%以上的相對酶活力,DSB全細胞催化劑的相對酶活力均略高于DSB游離酶。表明DSB全細胞催化劑比DSB游離酶在酸性環(huán)境中更穩(wěn)定,適用于工業(yè)應(yīng)用中的酸性環(huán)境。
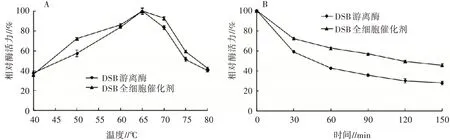
圖4 DSB全細胞催化劑和DSB游離酶的最適反應(yīng)溫度(A)及熱穩(wěn)定性(B)
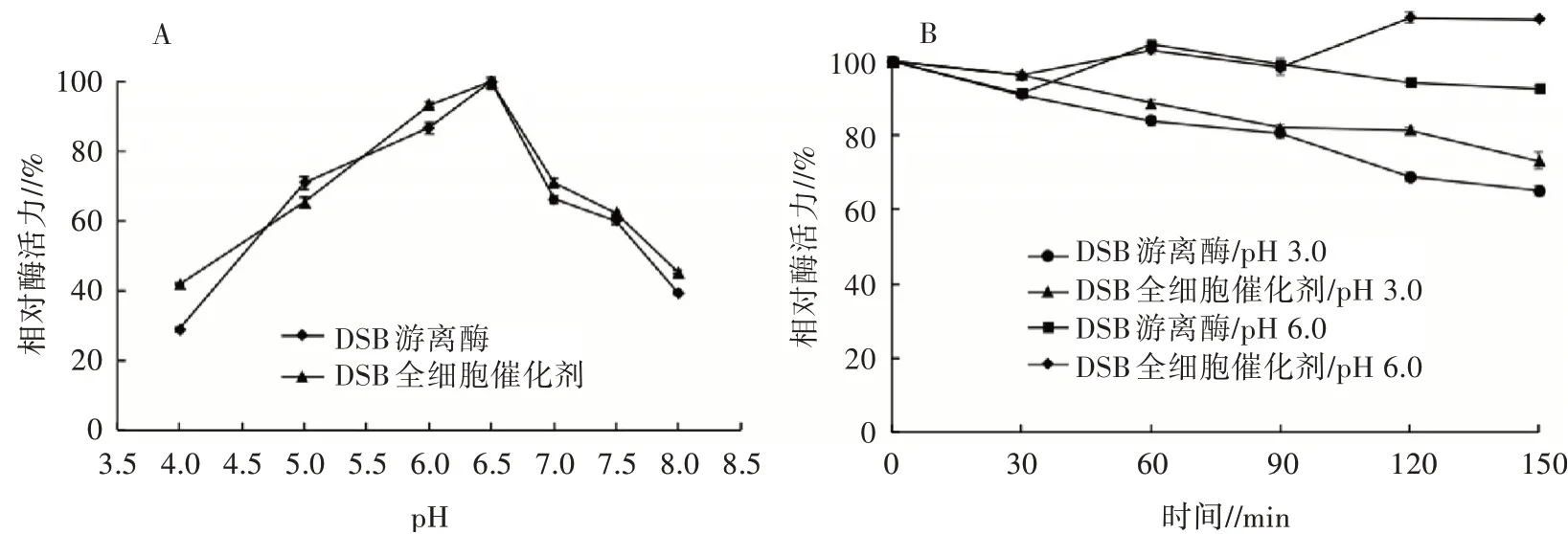
圖5 DSB全細胞催化劑和游離酶的最適pH(A)及pH穩(wěn)定性(B)
2.4.3 不同濃度NaCl和KCl對2種酶酶活力的影響在含不同濃度NaCl或KCl條件下分別測定2種酶的酶活力,從圖6(A)可以看出,當NaCl濃度在1%~5%時2種酶的酶活力均略有提高,當NaCl濃度高達10%和15%時,2種酶仍能維持80%以上的相對酶活力,但DSB全細胞催化劑的相對酶活力略高于DSB游離酶。而1%~15%的KCl對2種酶的酶活均有不同程度的促進作用,當KCl濃度為3%時DSB全細胞催化劑和DSB游離酶酶活力提高最多,分別達115%和113%。結(jié)果表明,DSB全細胞催化劑和DSB游離酶在高濃度鹽溶液中均能保持較高的酶活力,且DSB全細胞催化劑的耐鹽性更強,因此適用于工業(yè)中高鹽的環(huán)境。
2.4.4 有機溶劑對2種酶酶活力的影響4種常見有機溶劑對2種酶酶活力的影響見圖7。圖7(A)顯示5%~15%的甲醇對DSB全細胞催化劑的酶活力有顯著的促進作用,其中5%甲醇處理后DSB全細胞催化劑的相對酶活力增加幅度最大,達138%,且30%甲醇處理后DSB全細胞催化劑仍能維持94%的相對酶活力;而DSB游離酶的酶活力隨著甲醇體積分數(shù)的增加呈緩慢下降趨勢,30%甲醇處理后DSB游離酶殘余87%的相對酶活力。乙醇對2種酶酶活力的影響結(jié)果如圖7(B)所示,隨著乙醇體積分數(shù)的提高,2種酶的酶活力均緩慢降低,30%乙醇處理后2種酶均能維持75%以上的相對酶活力,但DSB全細胞催化劑的相對酶活力均略低于DSB游離酶。圖7(C)的結(jié)果顯示,5%丙三醇處理后DSB游離酶的相對酶活力增加至106%,隨后隨著丙三醇體積分數(shù)的提高其相對酶活力逐漸降低,30%丙三醇處理后殘留約60%的相對酶活力,而DSB全細胞催化劑隨著丙三醇體積分數(shù)的增加相對酶活力逐漸降低,且降低趨勢與游離酶基本一致。正丁醇對2種酶酶活力的影響見圖7(D),5%正丁醇處理后DSB全細胞催化劑和DSB游離酶的酶活力分別提高至122%和110%,隨后隨著正丁醇體積分數(shù)的提高2種酶的酶活力略微降低,30%正丁醇處理后2種酶均能保留90%以上的相對酶活力。
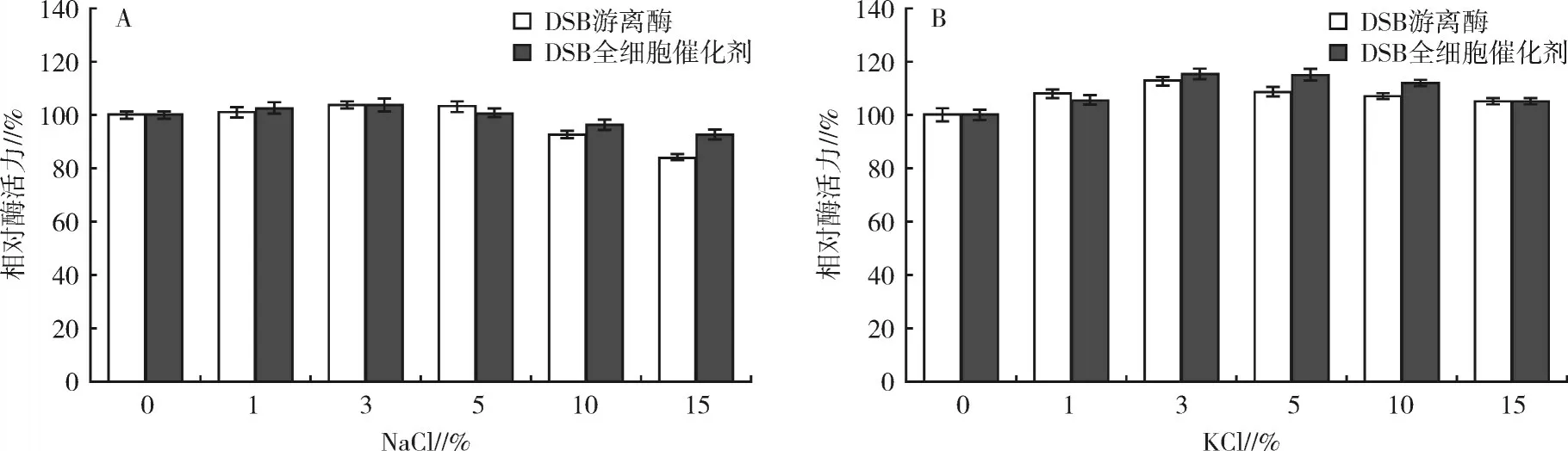
圖6 不同濃度NaCl和KCl對2種酶酶活力的影響
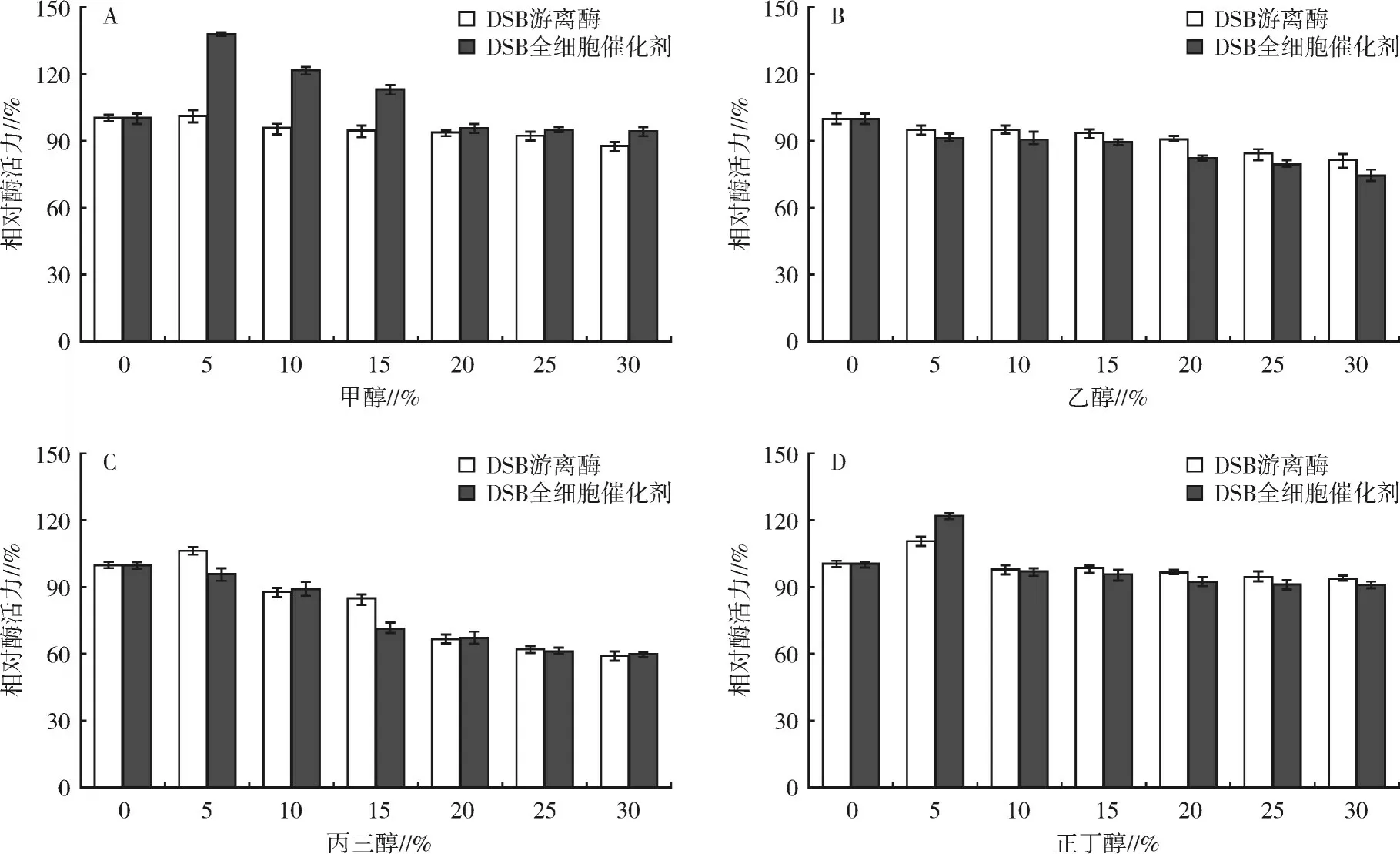
圖7 不同濃度有機溶劑對2種酶酶活力的影響
2.4.5 金屬離子和化學(xué)試劑對2種酶酶活力的影響不同金屬離子和化學(xué)試劑對2種酶酶活力的影響見表2。Na+、K+、Ca2+、Mg2+、Ni2+對2種酶酶活力均有不同程度的促進作用,且除Mg2+、K+外,其他金屬離子存在時DSB全細胞催化劑酶活力的提高幅度均高于DSB游離酶;Zn2+對DSB游離酶酶活力有輕微的抑制作用,而對DSB全細胞催化劑酶活卻有顯著的促進作用,其相對酶活力提高至120.05%;Mn2+、Pb2+、Fe3+、SDS、EDTA對2種酶酶活力均有不同程度的抑制作用,且同種金屬離子或化學(xué)試劑導(dǎo)致2種酶酶活力的下降幅度無顯著差別;Cu2+對2種酶酶活力均有顯著的抑制作用,DSB全細胞催化劑和DSB游離酶的酶活力分別下降至38.84%和52.79%;Li+、Co2+對2種酶酶活力無明顯影響。表明DSB全細胞催化劑和DSB游離酶對大部分金屬離子和化學(xué)試劑有一定耐受性,且DSB全細胞催化劑的耐受性優(yōu)于DSB游離酶。
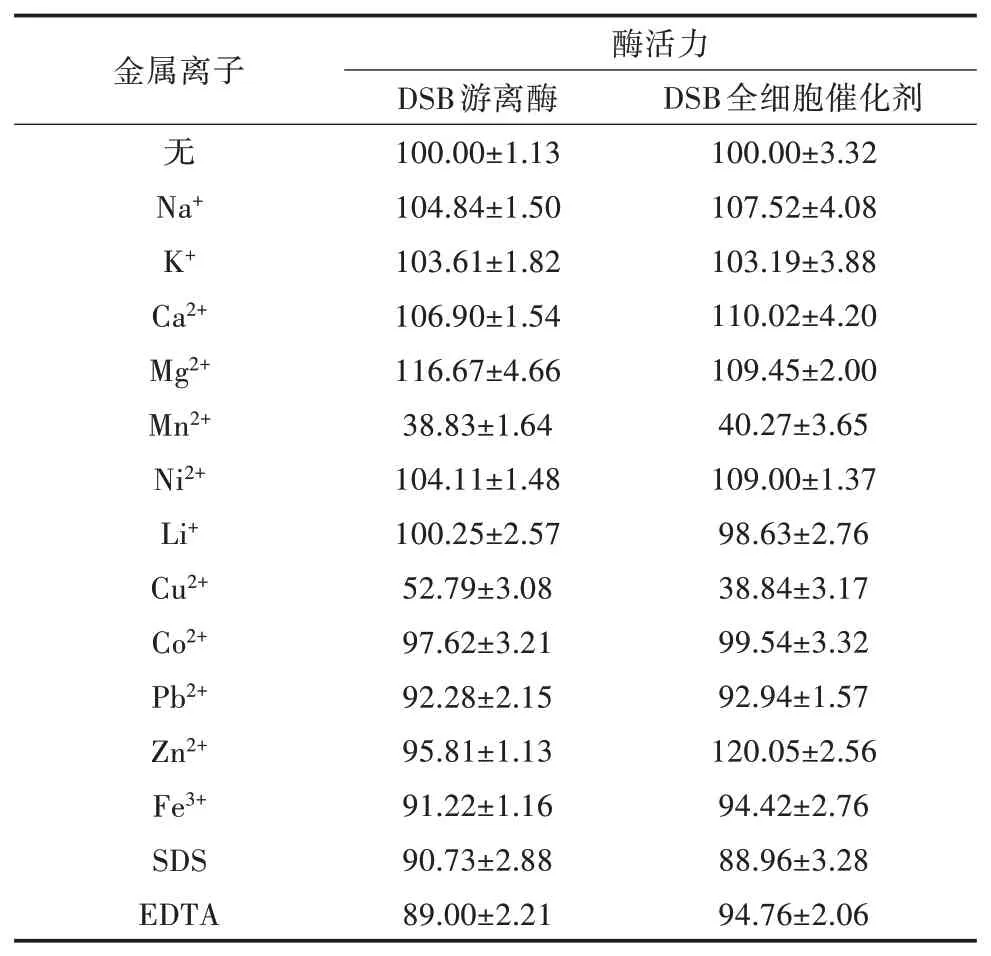
表2 不同金屬離子和化學(xué)試劑對2種酶酶活力的影響(單位:%)
3 小結(jié)與討論
目前已有多種不同來源的木聚糖酶在大腸桿菌、畢赤酵母、枯草芽孢桿菌等多個宿主中成功進行異源表達[16-18],但大部分均為分泌表達,需進行繁瑣的分離和純化后才能進行應(yīng)用。利用微生物表面展示技術(shù)將酶展示表達在微生物細胞表面獲得全細胞催化劑,可省去繁瑣的分離純化步驟,制備方法簡單[19]。本研究利用畢赤酵母表面展示技術(shù),以細胞壁蛋白Gcw61為錨定蛋白,將實驗室前期篩選獲得的耐熱木聚糖酶DSB進行展示表達,搖瓶發(fā)酵獲得的DSB全細胞催化劑的酶活力最高達13 510 U/g,高于木聚糖酶在其他宿主中展示表達的酶活力[4,20]。錨定蛋白的選擇對展示表達的酶活力有一定的影響,后續(xù)研究可選用更多的錨定蛋白進一步提高DSB全細胞催化劑的酶活力[21]。
木聚糖酶在飼料、造紙、食品、紡織等領(lǐng)域具有廣闊的應(yīng)用前景,然而在具體應(yīng)用過程中會有一些苛刻的環(huán)境條件,如高溫環(huán)境、強酸環(huán)境、高鹽環(huán)境、含有機溶劑的環(huán)境等,會對酶的活性造成很大的影響,從而降低了使用效率[22]。有研究表明在苛刻環(huán)境條件下全細胞催化劑較游離酶表現(xiàn)出更優(yōu)的穩(wěn)定性,Chen等[4]將來源于Cellulomonas fimi的木聚糖酶Cex展示表達在大腸桿菌細胞表面,Cex全細胞催化劑在40~65℃能維持70%以上的相對酶活力,而Cex游離酶在55℃時酶活力已降至43%,Cex全細胞催化劑在pH 6~8能維持約100%的相對酶活力,而Cex游離酶在pH 8時僅殘余29%相對酶活力;Yuzba?sheva等[10]在解脂耶氏酵母中構(gòu)建獲得脂肪酶Lip2p的全細胞催化劑,將全細胞催化劑和游離酶分別用EDTA、SDS、DMSO、Tween 80、Triton X-100試劑處理24 h,全細胞催化劑殘余的酶活力均高于游離酶。本研究將制備的DSB全細胞催化劑與DSB游離酶的酶學(xué)性質(zhì)進行了比較,分析發(fā)現(xiàn)在70℃高溫條件和pH 3.0的酸性條件下,DSB全細胞催化劑的穩(wěn)定性均優(yōu)于DSB游離酶;此外在高濃度NaCl、高濃度甲醇、多種金屬離子(Na+、Ca2+、Ni2+)的環(huán)境條件下,DSB全細胞催化劑均能維持較高的酶活力且穩(wěn)定性優(yōu)于DSB游離酶,為DSB全細胞催化劑的工業(yè)應(yīng)用提供一定的理論依據(jù),也為獲得穩(wěn)定性更好的酶制劑提供一定的理論基礎(chǔ)。
猜你喜歡
大自然探索(2023年7期)2023-11-14 13:08:06
石油石化綠色低碳(2019年6期)2019-02-13 09:39:01
石油石化綠色低碳(2019年6期)2019-01-14 01:16:22
智富時代(2018年3期)2018-06-11 16:10:44
浙江大學(xué)學(xué)報(工學(xué)版)(2016年11期)2016-06-05 09:21:04
Coco薇(2016年2期)2016-03-22 02:45:06
超硬材料工程(2016年1期)2016-02-28 22:20:04
中國資源綜合利用(2016年4期)2016-01-22 08:27:23
合成化學(xué)(2015年4期)2016-01-17 09:01:27
應(yīng)用化工(2014年3期)2014-08-16 13:23:50
