轉betA基因小黑楊生長及適應性1)
2021-04-27 09:40:24葛夢妍顧宸瑞陳坤王偉王楚劉桂豐
東北林業大學學報 2021年4期
關鍵詞:生長
葛夢妍 顧宸瑞 陳坤 王偉 王楚 劉桂豐
(林木遺傳育種國家重點實驗室(東北林業大學),哈爾濱,150040)
汪廣宇 姜靜
(黑龍江嘉懋園林建設股份有限公司) (林木遺傳育種國家重點實驗室(東北林業大學))
高等植物在受到滲透脅迫后,其體內的甜菜堿醛脫氫酶活性升高,并在細胞質中積累甜菜堿,進而提高了植物對干旱及鹽脅迫的適應性[1-4]。植物的甜菜堿是以膽堿為底物合成,經膽堿加單氧酶(CMO)和甜菜堿醛脫氫酶(BADH)兩步催化完成[5-6]。而來自大腸桿菌的betA基因編碼的膽堿脫氫酶(CDH)具有膽堿加單氧酶(CMO)與甜菜堿醛脫氫酶(BADH)兩個酶的功能,能夠催化甜菜堿合成的兩個連續反應,既能以膽堿為底物,也能以甜菜堿醛為底物進行甜菜堿的合成[7]。因此,編碼膽堿脫氫酶基因(betA)被認為是植物耐鹽基因工程中的有效基因,已經廣泛用于植物的耐鹽轉基因研究[8-17]。筆者所在的團隊以東北地區廣泛推廣的小黑楊(Populussimonii×P.nigra)為轉基因受體,開展膽堿脫氫酶基因(betA)的轉化研究。獲得的轉betA基因小黑楊,苗期NaCl脅迫試驗顯示,轉基因株系的甜菜堿含量均顯著高于對照株系,根據NaCl脅迫后苗木的抗氧化酶活性、丙二醛(MDA)質量摩爾濃度、鹽害指數等指標,選出TB1、TB2、T4、T5、T6株系為耐鹽優良株系[18-21]。在輕度鹽堿地營建4 a的轉betA小黑楊試驗林分析顯示,T01株系其樹高、胸徑和材積生長表現最好,分別較對照株系高14.74%、18.81%和47.50%,根據隸屬函數值選出T1、T6和T8轉基因株系為耐鹽優良株系[22]。由于林木具有生長周期長、受自然因素影響大、可控制性差的特點,部分性狀表現早、晚期相關性弱,對轉基因小黑楊耐鹽性的早期評價尚需進一步驗證,為此,試驗針對13年生的轉基因小黑楊林分再次進行調查,檢測目標基因的穩定性,評價轉基因株系的生長適應性,研究結果為環境釋放及生產性試驗提供指導。
1 試驗地概況
試驗點設置在黑龍江省大慶市林原鎮新華六隊,屬于半干旱碳酸鹽堿地的代表。年平均氣溫4.2 ℃,極端最低氣溫-39.2 ℃,最熱月平均氣溫23.3 ℃,極端最高氣溫39.8 ℃,年均無霜期143 d。年均降水量427.5 mm,年均蒸發量1 635 mm。土壤pH=10.8,有機質質量分數1.98%。
2 材料與方法
試驗材料為小黑楊轉betA基因株系。2004年春,將小黑楊轉基因(betA)株系與對照株系1年生扦插苗去掉地上部分,僅用苗根在大慶進行定植試驗。試驗林均按照完全隨機區組設計,4次重復。參試株系14個,其中轉基因株系13個(分別命名為T01、T02、T04、T05、T06、T07、T08、T09、T10、T11、T12、T13、T14),對照株系(CK)1個,采用8株小區4行排列,株行距2 m×3 m,試驗地周圍設置3行保護行。造林后按照常規林分管理,包括定桿、除草、松土、病蟲害防治等。
2.1 目標基因的分子檢測
PCR檢測引物序列:根據betA序列,用DNASTAR的Primer Select軟件,設計引物betA-F:5′-TACATCATTATTGGTGCCGGCTCA-3′;betA-R:5′-CATCCCATTTGCCACAAAATATCCC-3′。
PCR擴增檢測:采用植物總DNA提取試劑盒提取參試株系葉片總DNA,以betA載體質粒DNA為陽性對照,以水為陰性對照,用上述引物分別進行PCR擴增。PCR反應體系為:在25 μL反應體積中含DNA模板1 μL,上、下游引物各1 μL(10 μmol/L),1.1×T3 Super PCR Mix 12.5 μL,加ddH2O補齊至25 μL。PCR擴增程序為:96 ℃預變性3 min;進行PCR循環;96 ℃變性30 s,60 ℃退火30 s,72 ℃延伸40 s,34個循環后繼續于72 ℃延伸5 min,16 ℃保溫。PCR產物用1.0%的瓊脂糖凝膠檢測。
RT-PCR擴增檢測:采用離心柱型總RNA快速提取試劑盒提取參試株系葉片總RNA,用ReverTra Ace qPCR RT Master Mix試劑盒反轉錄為cDNA,將cDNA稀釋10倍后用于RT-PCR模板。RT-PCR引物、反應體系及程序同上,PCR產物用1.0%的瓊脂糖凝膠檢測。
2.2 試驗林性狀調查及數據分析
2017年秋,調查了小黑楊轉基因株系試驗林的所有單株,采用超聲波測高測距儀測量了樹高,精確至0.1 m;采用胸徑尺測量了胸徑,精確至0.1 cm;依據各試驗點內的保存株數計算各株系保存率;根據小黑楊的二元材積表[23]計算單株材積:
V=0.000 730 584 099-0.000 040 168 020 5D2+
0.000 034 384 964 4D2H+0.000 337 503 207H+
0.000 002 052 761 11DH2。
式中:V為小黑楊的材積(m3);D為測得的小黑楊胸徑值(cm);H為測得的小黑楊樹高值(m)。
經過13 a的造林試驗,試驗點中保留株系共計11個,包括10個轉基因株系(T01、T02、T04、T05、T06、T08、T09、T12、T13、T14)和1個對照株系(CK),但由于T02、T04、T09、T14保存率較低,未納入統計分析,樹高、胸徑、材積及保存率僅統計分析了7個株系,即T01、T05、T06、T08、T12、T13和CK株系。
數據分析方法:利用SPSS19.0軟件對樹高、胸徑、材積和保存率4個性狀進行方差分析及多重比較,并通過隸屬函數法選擇出在試驗點適應性較好的株系。
式中:u(Xi)為第i個性狀的隸屬函數值,Xi為第i個性狀的數值,Xmin為第i個性狀的最小值,Xmax為第i個性狀的最大值。
計算材積和保存率2個性狀的隸屬函數值并給予權重,用加權單因子法計算出各株系的隸屬函數均值。
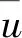
重復力計算公式:
式中:F為方差分析中株系間F值。
遺傳增益計算公式:
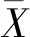
變異系數計算公式:
式中:σ為標準差;μ為平均值。
3 結果與分析
3.1 betA基因在小黑楊基因組中整合和表達
以待測株系葉片總DNA為模板,以betA質粒為陽性對照,以水為陰性對照,對外源betA基因進行PCR擴增,結果如圖1A,轉基因株系均在1 500 bp呈現特異性擴增條帶,與預期的betA基因序列(1 470 bp)大小吻合,而對照株系及水對照未見擴增譜帶,說明13年生時,外源betA基因仍穩定整合于轉基因株系基因組中,尚未發生丟失。
以上述株系的cDNA為模板,以betA質粒為陽性對照,進行RT-PCR檢測,結果如圖1B,轉基因小黑楊在1 500 bp處均呈現特異性擴增譜帶,與預期的betA基因序列(1 470 bp)大小吻合,而陰性對照均未見擴增譜帶,說明轉基因小黑楊中導入的外源betA基因能夠在mRNA水平上正常表達。
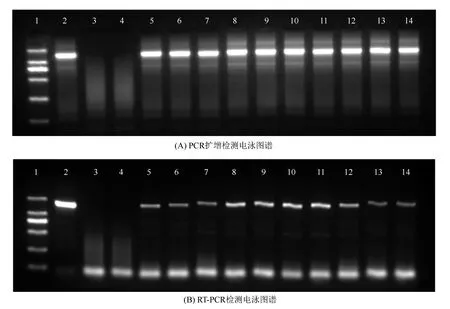
1.Marker;2.陽性質粒;3.陰性水對照;4.對照株系;5~14.轉betA株系T01、T02、T04、T05、T06、T08、T09、T12、T13、T14。
3.2 轉基因株系生長性狀差異顯著性
在鹽堿地開展造林試驗的轉基因株系,生長量及保存率是評定參試株系抗逆性的重要指標。因此,對參試的7個株系生長性狀及保存率方差分析顯示(表1),株系間各生長性狀及保存率的差異均達到了極顯著水平(P<0.01);株系間材積和保存率的變異系數均較大,分別為36.61%和28.20%,進一步對各性狀方差分量分析發現,株系間樹高、胸徑、材積及保存率的方差分量比例分別為43.42%、76.00%、63.12%和35.69%,說明株系間的差異對各株系的生長表現有較大影響。基于上述轉基因株系間的生長性狀存在顯著性差異,開展優良株系選擇很有必要。
3.3 優良株系的選擇
材積可以代表樹木的生長量,因此試驗利用材積、保存率2個性狀采用模糊數學隸屬函數法對7個株系進行綜合評價(表2),綜合評價中對材積、保存率分別給予0.5的權重值,求算出隸屬函數均值。隸屬函數均值越高,其綜合評價越好,若將隸屬函數均值在0.8以上的評為優良株系,則入選株系為T01、T06和T08,其中T01名列前茅,為最優株系,樹高、胸徑、材積均值分別為11.03 m、11.76 cm、0.055 m3,遺傳增益分別為12.1%、11.2%和34.5%;入選的T06、T08株系材積均值分別高于最差株系(T05)的228.64%、286.13%,材積遺傳增益分別為5.8%、22.8%。本次入選的株系與2008年對5年生的該試驗林優良株系選擇結果一致(表2),在2008年的選擇分析中生長表現最差的T04、T07、T10、T11、T14等5個轉基因株系,本次調查發現上述5個株系保存率已經降至10%以下,故此未納入本次統計分析。由表2還可見,經過13 a的造林對比試驗,入選的T01、T06、T08優良株系的保存率幾乎沒有發生變化,說明轉betA小黑楊5年生時的選擇結果準確可靠,外源betA基因在小黑楊基因組適宜位點的插入確實可以顯著提高其耐鹽性。
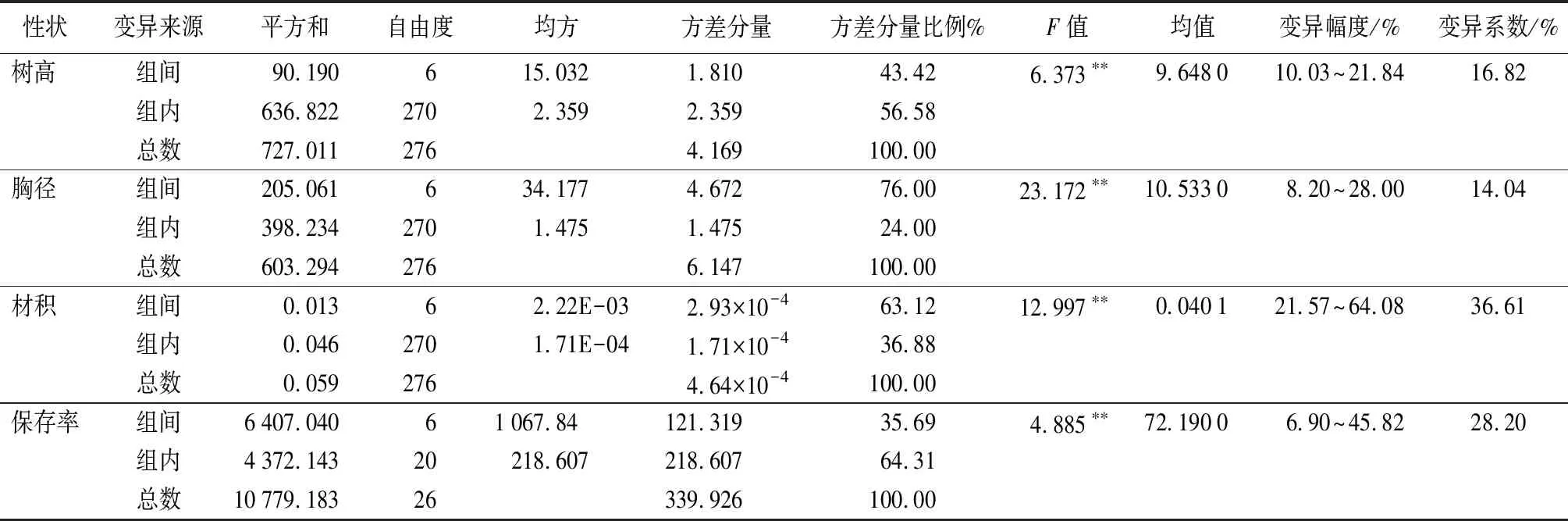
表1 參試株系各性狀方差分析
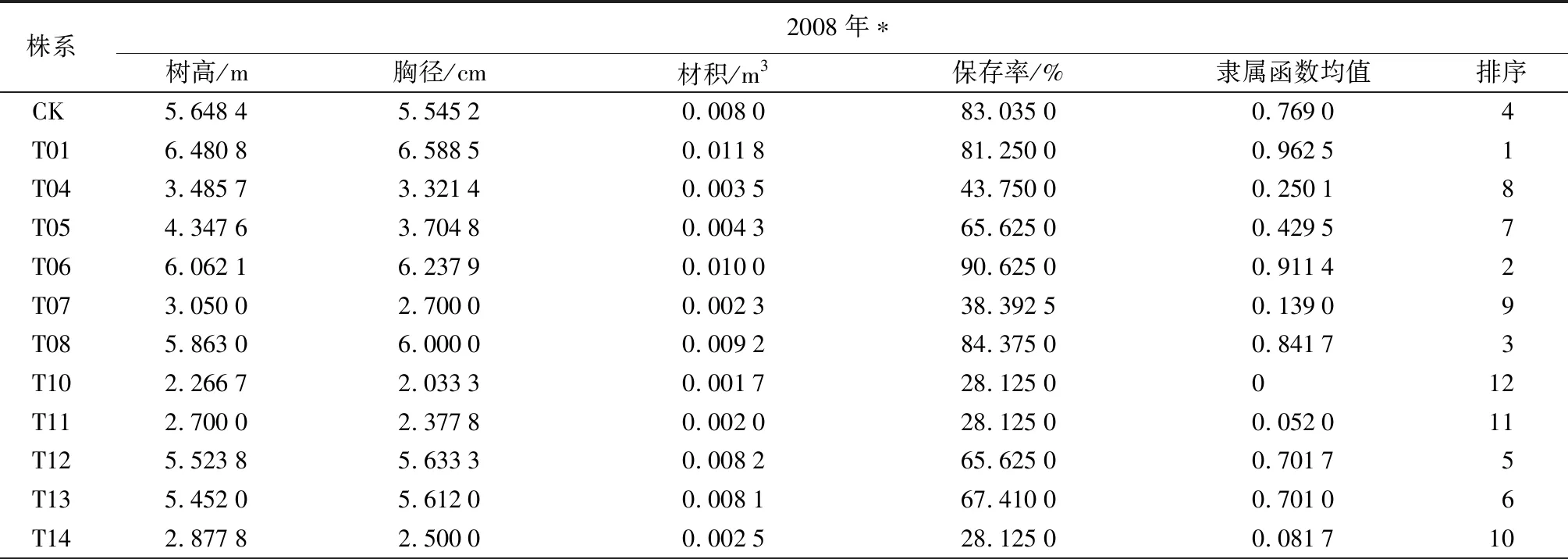
表2 兩次調查參試株系各性狀平均值比較
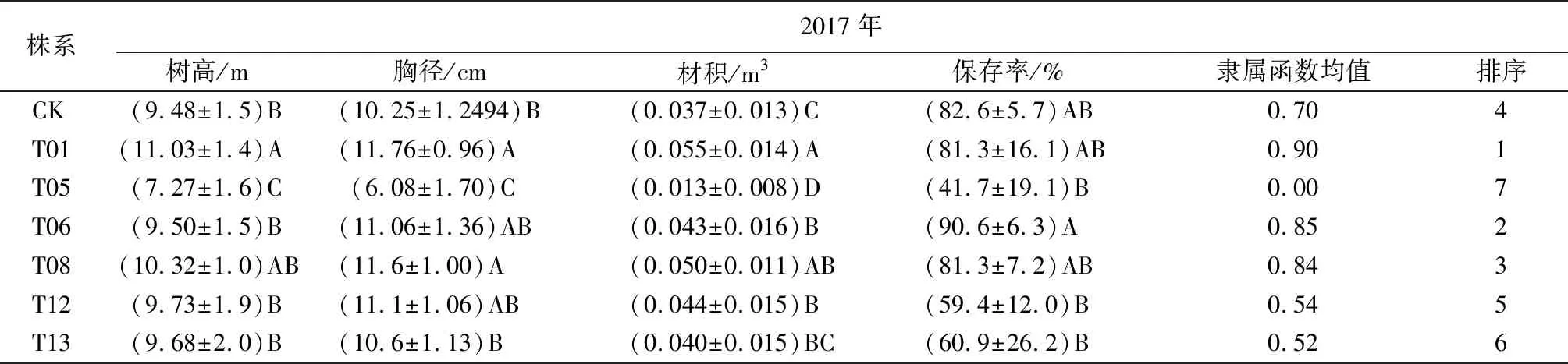
株系2017年樹高/m胸徑/cm材積/m3保存率/%隸屬函數均值排序CK(9.48±1.5)B(10.25±1.2494)B(0.037±0.013)C(82.6±5.7)AB0.704T01(11.03±1.4)A(11.76±0.96)A(0.055±0.014)A(81.3±16.1)AB0.901T05(7.27±1.6)C(6.08±1.70)C(0.013±0.008)D(41.7±19.1)B0.007T06(9.50±1.5)B(11.06±1.36)AB(0.043±0.016)B(90.6±6.3)A0.852T08(10.32±1.0)AB(11.6±1.00)A(0.050±0.011)AB(81.3±7.2)AB0.843T12(9.73±1.9)B(11.1±1.06)AB(0.044±0.015)B(59.4±12.0)B0.545T13(9.68±2.0)B(10.6±1.13)B(0.040±0.015)BC(60.9±26.2)B0.526
4 結論與討論
4.1 外源betA基因在轉基因小黑楊中的穩定表達
快速發展的生物技術下轉基因作物新品種的不斷培育為全球糧食安全保障帶來了新的機遇,同時,轉基因作物的環境釋放和商品化生產也引起了全世界對轉基因作物生物安全問題的極大關注[24]。1983年世界首例轉基因植物培育成功,為植物基因工程帶來了廣闊前景,但轉基因植物的應用也越來越受到爭議。目前對由轉基因逃逸可能導致的生態安全性等方面問題還沒有結論性的結果,但已引起政府、社會和科學界的廣泛關注[25]。筆者所在研究團隊也對轉基因小黑楊進行了生物安全性評價,采集了轉基因株系根際土壤及周邊雜草,分別對提取的DNA樣品進行選擇劑及目標基因的PCR檢測,結果表明,目標基因既沒有逃逸也沒有釋放到環境中,而是穩定存在于轉基因小黑楊的基因組中。
本研究通過對轉betA小黑楊的分子檢測表明,betA基因在轉基因株系的基因組中能夠長期穩定整合并能夠正常表達。13年生的轉基因小黑楊株系均已到達可以用材的樹齡,本研究的跟蹤調查周期足夠長,選擇結果更加準確可靠。
4.2 轉betA基因能夠提高植物的耐鹽性
積累滲透保護物質是植物在滲透脅迫條件下一種重要而普遍的適應機制,對于禾本科和藜科植物,最有效的物質當屬季胺類化合物中的甘氨酸甜菜堿。它作為滲透保護物質一方面能夠使細胞保持適當的滲透勢而防止脫水;另一方面能夠對生物大分子的結構及功能起穩定、保護作用。但煙草(NicotianatabacumL.)、番茄(LycopersiconesculentumMill.)、馬鈴薯(SolanumtuberosumL.)、水稻(OryzasativaL.)等作物卻沒有合成甘氨酸甜菜堿的能力[8]。
早期研究表明,將betA基因轉入煙草中并將轉基因株系置于高濃度氯化鈉溶液中培養,轉基因煙草生長速度更快且能夠積累甜菜堿,耐鹽性有顯著提升[26-27];將betA基因轉入番茄中,在200 mmol/L的NaCl處理下,轉基因番茄與對照番茄相比葉片質膜透性降低23%,耐鹽性增加[8];將betA基因轉入棉花(Gossypiumspp.)中,在鹽脅迫和低溫脅迫下,轉基因株系葉片中甘氨酸甜菜堿的含量均顯著高于對照株系,且轉基因棉花的種子萌發率較對照株系高近一倍,轉基因棉花株系也表現出生物量增加,葉片失水慢的性狀[28];將betA基因轉入小麥(TriticumaestivumL.),經鹽脅迫和干旱脅迫后,轉基因小麥葉片中甘氨酸甜菜堿含量較對照株系高約2倍,在鹽脅迫下的種子萌發率為對照株系的1.45倍,同時轉基因小麥也表現出了生物量的增加和光合作用的改善,說明betA基因的轉入顯著提高了小麥的抗逆性[29-30]。2001年,筆者所在的林木遺傳育種國家重點實驗室首次將betA基因轉入小黑楊中[31],隨后在小黑楊生長發育的不同階段都進行了耐鹽性試驗并證實了betA基因確實能夠提高小黑楊的耐鹽性。本研究中,經過在鹽堿地上的造林試驗,面對長達13年的鹽脅迫,轉betA小黑楊株系依然表現出了良好的生長特性。
4.3 不同轉基因株系間生長性狀的差異
外源基因導入小黑楊基因組時,有時會對固有基因的轉錄表達產生影響,進而影響了轉基因株系的生長發育,這是造成不同轉基因株系間性狀差異的主要因素,農桿菌介導的T-DNA插入不僅破壞了插入位點的基因功能,而且因為T-DNA序列已知,使得被破壞的基因序列的分離相對簡單和快捷,因此該技術被廣泛地用于功能基因的研究[32]。在本研究中,最初在造林地點對12個小黑楊株系進行了造林對比試驗,經過4 a造林試驗后,雖然全部存活,但幾個耐鹽性差的轉基因株系(T04、T07、T10、T11、T14)的生長量和保存率明顯小于其他優良耐鹽株系和對照株系;13 a后,上述耐鹽性差的5個轉基因株系陸續死亡,說明外源基因的插入非但沒有提高這些株系的抗鹽堿能力,反而影響了該株系的正常生長發育。T01、T06和T08這3個株系在鹽堿地具有生長快、保存率高的優良表現,說明外源基因在這3個株系株系的基因組中插入到適宜的位點,既不影響轉基因小黑楊的正常生長,又使植株獲得了比較好的耐鹽堿能力。
通過對兩次優良株系的選擇結果進行對比可以發現,2017年和2008年所選擇出的優良株系一致,且兩次調查中,各株系的生長表現排序完全相同。說明小黑楊作為速生樹種,在造林試驗的初期就可以獲得準確可靠的選擇結果。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
