過量表達紫花苜蓿MsHB7基因對擬南芥耐旱性的影響
2021-04-23 02:14:24候怡謠李霄龍瑞才楊青川康俊梅郭長虹
草業學報 2021年4期
關鍵詞:植物
候怡謠,李霄,龍瑞才,楊青川,康俊梅,郭長虹
(1.哈爾濱師范大學生命科學與技術學院,黑龍江省分子細胞與遺傳育種重點實驗室,黑龍江哈爾濱150000;2.中國農業科學院北京畜牧獸醫研究所,北京100193)
植物在生長發育過程中會遭遇到各種非生物脅迫,包括鹽、干旱、低溫等。在這些脅迫下,植物會產生一系列的生理和分子變化[1]。轉錄因子在這個過程中發揮著重要調控作用,它能特異性地結合下游靶基因啟動子上的順勢作用元件,從而調控下游基因的表達并調節植物的耐受性[2-3]。植物中存在許多與非生物脅迫應答有關的轉錄因子,如:MYB、WRKY、bZIP、NAC、DREB等[4]。
含有同源異型域(homeodomain,HD)的轉錄因子與多種發育過程相關,HD由同源框(homeobox,HB)基因編碼,HB基因最初是在果蠅中發現的,因其同源異型效應而命名,隨著1991年在玉米(Zea mays)中首次發現HB基因Knotted-1之后,許多植物中的HB基因被分離出來[5-6]。同源異型域-亮氨酸拉鏈(HD-Zip)蛋白是HD轉錄因子超家族中的一類轉錄因子[5],HD結構域和亮氨酸拉鏈(leucine zipper,LZ)元件緊密相連,HD負責與DNA的特異性結合,LZ使HD-Zip蛋白二聚,并且這兩個結構域之間的聯系是植物所特有的[7]。HD-Zip轉錄因子家族根據其成員的結構和功能被分為了HD-Zip I、HD-ZipⅡ、HD-ZipⅢ和HD-ZipⅣ4個亞家族[5]。其中HDZip I蛋白主要參與植物對非生物脅迫的響應[8]。例如在水分缺乏、滲透脅迫或外源ABA處理下,擬南芥(Arabidopsis thaliana)ATHB6基因的表達顯著上調[9],ATHB7和ATHB12在干旱和脫落酸(abscisic acid,ABA)條件下上調表達,并且可作為莖伸長的負調節因子,當在擬南芥中過表達ATHB7和ATHB12時,會出現與缺水條件下野生型擬南芥莖伸長降低一樣的表型[10-12]。HD-ZipⅡ類基因的表達一般受光照條件的調控,參與植物的生長發育以及避陰反應[5],如ATHB2/HAT4、HAT1、HAT2、HAT3和ATHB4受紅光/遠紅光比值變化的調節,從而誘導大多數被子植物避陰[13]。HD-ZipⅢ類蛋白參與胚胎發生、分生組織形成、側生器官發育、葉極性、維管系統發育和生長素的運輸[5],如REV、PHB、PHV、CNA和ATHB8基因調節胚后分生組織的形成[14]。HD-ZipⅣ類轉錄因子與表皮細胞分化、花青素積累、根發育以及毛狀體的形成有關[5],如GLABRA2(GL2)基因參與擬南芥毛狀體的形態發育和成熟[15]。
紫花苜蓿(Medicago sativa)是世界上廣泛種植的豆科植物之一,因其產量高、品質好、適應性強而被譽為“牧草之王”[16]。干旱是影響植物生長發育的重要因素,嚴重影響了苜蓿的產量。因此,研究與干旱脅迫相關的基因在紫花苜蓿抗逆品種的培育中具有重要意義。本研究通過在擬南芥中過量表達MsHB7基因,并對轉基因植株進行干旱處理,進而分析該基因在植物干旱脅迫過程中的功能,為將來紫花苜蓿抗旱育種提供理論支撐。
1 材料與方法
1.1 材料
實驗所用的紫花苜蓿中苜一號(M.sativacv.Zhongmu-1)和擬南芥(Col-0,WT)均由實驗室保存。植物材料種植于人工氣候箱,培養條件為24/22℃(晝/夜),60%濕度,16 h光照/8 h黑暗。農桿菌GV3101購于華越洋生物公司。
1.2 方法
1.2.1 紫花苜蓿MsHB7的克隆 以擬南芥ATHB7蛋白序列為參考,在NCBI(https://www.ncbi.nlm.nih.gov/)數據庫中進行序列比對,在蒺藜苜蓿(Medicago truncatula)中找到一個同源基因(XM_003602273),根據該序列設計特異性引物MsHB7-F/R(表1),以中苜一號cDNA為模板,進行PCR擴增獲得此基因的編碼區序列,之后將其連接到pEASY-T5載體上并轉化到大腸桿菌Trans1-T1感受態中,之后經菌液PCR驗證后測序。
1.2.2 生物信息學分析 通過ExPASy網站(http://web.expasy.org/protparam/)預測蛋白質的等電點及分子量,NCBI(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)預測蛋白的保守結構域,利用DNAMAN軟件與其他植物中的HD-Zip同源蛋白進行序列比對分析,用MEGA 7.0.14軟件將目的蛋白與擬南芥中HD-Zip類蛋白進行進化分析(鄰 接 法,neighbor-joining method)[17]。利 用Predict-Protein(https://www.predictprotein.org/)網站進行亞細胞定位的預測。
1.2.3 表達分析 使用5%聚乙二醇(polyethylene glycol,PEG)對在1/2 Hoagland營養液中生長4周的紫花苜蓿中苜一號進行干旱處理,并分別在處理0,1,2,4,8,12,24和48 h之后對紫花苜蓿的地上部分和根進行取材,提取總RNA,之后以Msactin2為內參基因,qMsactin2F/R和qMsHB7F/R分別為Msactin2和MsHB7的引物(表1),對MsHB7的相對表達量進行分析。
1.2.4 超表達載體pBI121-MsHB7的構建以及對擬南芥的轉化 以pEASY-T5-MsHB7質粒為模板,含有XbaI和BamHI酶切位點的pMsHB7F/R(表1)為引物,進行PCR擴增,將擴增產物連接到pBI121表達載體上,獲得pBI121-MsHB7重組質粒,并轉化到大腸桿菌Trans1-T1感受態中,測序之后提取質粒,轉化到農桿菌GV3101中,利用花序侵染法[18]轉化野生型擬南芥。
1.2.5 轉基因擬南芥的篩選及轉基因植株的檢測 將收獲的擬南芥T0代種子播種于含有卡那霉素(50 mg·L-1)抗性的1/2 MS固體培養基上,篩選出轉基因擬南芥株系并使用引物MsHB7-F1/R1對這些株系進行PCR鑒定(表1),將陽性轉基因植株進行保種并繁殖,直至獲得T3代轉基因植株,用于后續實驗。

表1 引物列表Table 1 Primer list
1.2.6 轉基因擬南芥的干旱處理及耐旱性測定 將消毒之后的轉基因擬南芥和野生型擬南芥種子種在1/2 MS培養基上,4℃春化3 d,于光照培養箱中培養14 d后,移至營養土和蛭石體積比為1∶1的營養基質中,一周后將植株分為兩組,一組正常澆水作為對照,一組不澆水進行干旱處理,18 d后觀察其表型變化并測定對照組和處理組植株的相對含水量,首先稱取葉片0.2 g,記為Wf,之后將葉片浸入水中8 h,稱其飽和鮮重Wt,再于105℃烘箱中殺青15 min,75℃條件下烘干至恒重,稱干重,記為Wd,按照公式計算相對含水量:相對含水量=(Wf-Wd)/(Wt-Wd)×100%。另外再分別取材,液氮速凍后于-80℃冰箱中保存,用于脯氨酸(proline,Pro)、丙二醛(malondialdehyde,MDA)含量的測定和抗逆相關指示基因的表達分析。采用酸性茚三酮法測定脯氨酸含量,具體操作步驟為:稱取約0.1 g組織,研磨后加入1 mL 3%磺基水楊酸,之后沸水浴提取10 min,12000 r·min-1離心10 min后取上清,冷卻,同時將脯氨酸用蒸餾水稀釋為20、15、10、8、6、4、2、1、0μg·mL-1,取0.25 mL提取液或脯氨酸溶液,加入0.25 mL冰乙酸與0.25 mL 2.5%酸性茚三酮,沸水浴30 min,冷卻后,加入0.5 mL甲苯,振蕩30 s,靜置片刻,然后吸取上層溶液于520 nm波長處比色,根據脯氨酸溶液建立標準曲線,以此計算樣本脯氨酸含量,計算公式為脯氨酸含量(μg·g-1FW)=(C×V1)/(V2×W),其中,C為脯氨酸含量(μg);V1為提取液總體積(mL);V2為測定時所用提取液體積(mL);W為樣品鮮重(g)。丙二醛的測定步驟為:稱取約0.1 g組織,研磨后加入1 mL 10%三氯乙酸(TCA)溶液,12000 r·min-1離心10 min,取0.25 mL上清液加入0.25 mL 0.6%硫代巴比妥酸(TBA)溶液,沸水浴15 min后冷卻、離心,分別在450、532、600 nm波長下測定上清液的吸光值,計算公式:丙二醛含量(nmol·g-1FW)=[6.452×(A532-A600)-0.559×A450]×V1/(W×V2)[V1:提取液總體積(mL);V2:測定時所用提取液體積(mL);W:樣品鮮重(g)]。
1.2.7 轉基因擬南芥抗逆相關基因的表達分析 提取上述所取樣品的總RNA并反轉錄為cDNA。通過qRT-PCR技術,以擬南芥ATactin7為內參基因,qATactin7F/R為內參基因引物(表1),檢測轉基因和野生型擬南芥中MsHB7基因及幾個抗逆相關基因的表達量。
1.3 統計分析
利用Excel 2019對數據進行統計分析,采用單因素方差對熒光定量和生理指標進行差異顯著性分析,采用GraphPad Prism 6作圖。
2 結果與分析
2.1 紫花苜蓿MsHB7的生物信息學分析
對獲得的MsHB7基因序列(GenBank:MT508841)進行生物信息學分析,該基因包含一個738 bp的開放閱讀框,可編碼245個氨基酸,蛋白質的分子量為29020.17 Da,理論等電點5.33,含有兩個結構域:同源異型域(41~91)和亮氨酸拉鏈結構域(93~131)。進化樹分析結果表明,MsHB7與擬南芥第I類同源異型域-亮氨酸拉鏈蛋白聚為一類,并與擬南芥中的ATHB7和ATHB12親緣關系較近(圖1)。多重序列比對表明,MsHB7與其他植物中的同源蛋白都具有保守的同源異型框結構域序列和亮氨酸拉鏈結構域序列(圖2)。亞細胞定位預測表明MsHB7定位于細胞核上。
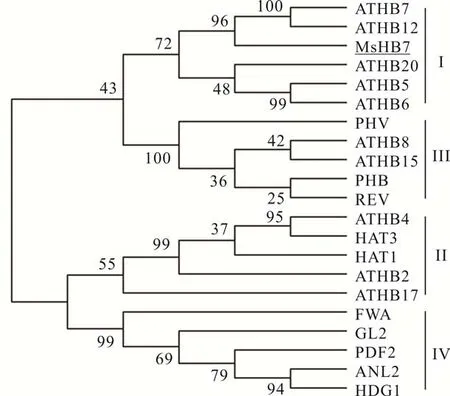
圖1 MsHB7與擬南芥中一些同源異型域-亮氨酸拉鏈蛋白進化樹分析Fig.1 Phylogenetic tree analysis of MsHB7 and some homeodomain-leucine zipper proteins from A.thalianaATHB7:NP_182191.1;ATHB12:NP_191748.1;ATHB20:NP_186771.1;ATHB5:NP_201334.1;ATHB6:NP_565536.1;PHV:NP_174337.1;ATHB8:NP_195014.1;ATHB15:NP_849795.1;PHB:NP_181018.1;REV:NP_200877.1;ATHB4:NP_182018.1;HAT3:NP_191598.1;HAT1:NP_193476.1;ATHB2:NP_193411.1;ATHB17:NP_178252.2;FWA:NP_567722.1;GL2:NP_001185443.1;PDF2:NP_567274.1;ANL2:NP_567183.2;HDG1:NP_191674.1.
2.2 干旱條件下MsHB7表達模式分析
對生長4周的紫花苜蓿中苜一號使用5%PEG模擬干旱處理,分別對處理0,1,2,4,8,12,24和48 h之后紫花苜蓿的地上部分和根的總RNA進行qRTPCR分析。結果顯示:MsHB7基因在地上部分中隨著處理時間的增加呈先上調然后下調的循環表達趨勢,和地上部分的表達模式不同,在根中MsHB7基因的表達量在1和2 h下調表達,而后在4 h顯著上調表達(圖3)。
2.3 轉MsHB7基因擬南芥的鑒定
利用花序侵染法轉化野生型擬南芥植株,通過篩選之后獲得6個轉基因株系,對其中3個轉基因株系(L1,L2和L6)進行PCR檢測的結果顯示,轉基因株系均擴增出目的條帶,而野生型擬南芥則未能擴出目的條帶(圖4)。利用qRT-PCR檢測MsHB7基因在野生型和轉基因擬南芥中的表達量,結果表明,MsHB7基因在轉基因植株中成功表達,而在野生型擬南芥中沒有檢測到其表達(圖5)。

圖2 MsHB7與其他植物中同源異型域-亮氨酸拉鏈蛋白的氨基酸序列比對分析Fig.2 The amino acid sequence alignment analysis between MsHB7 and homeodomain-leucine zipper proteins from other plantsA:同源異型框結構域序列比對Sequence alignment of homeobox domain;B:亮氨酸拉鏈結構域序列比對,6個保守的亮氨酸殘基用“L”表示Sequence alignment of leucine zipper domain,the six conserved leucine residues are represented by“L”.XP_003602321.2:蒺藜苜蓿M.truncatula ATHB-12;XP_004502719.1:鷹嘴豆Cicer arietinum ATHB-12-like;XP_017420762.1:小豆Vigna angularis ATHB-12-like;XP_016695089.1:陸地棉Gossypium hirsutum ATHB-12-like;XP_018842791.1:核桃Juglans regia ATHB-12-like;XP_002320889.1:毛果楊Populus trichocarpa ATHB-7;XP_018729630.1:巨桉Eucalyptus grandis ATHB-12-like;XP_007218213.1:桃樹Prunus persica ATHB-12;XP_012088620.1:麻瘋樹Jatropha curcas ATHB-12;NP_001281285.1:蘋 果Malus domestica ATHB-12-like;XP_016538252.1:辣 椒Capsicum annuum ATHB-12-like;NP_001311821.1 ATHB-12-like:煙草Nicotiana tabacum ATHB-12-like;XP_006491341.1:橙Citrus sinensis ATHB-7;XP_003548207.2:大豆Glycine max ATHB-12;XP_004230017.1:番茄Solanum lycopersicum ATHB-12;NP_191748.1:擬南芥A.thaliana ATHB-12.
2.4 干旱脅迫下轉MsHB7基因擬南芥表型分析
將正常生長的擬南芥幼苗干旱處理18 d,3個轉基因株系(L1,L2和L6)與WT相比,萎蔫得更為嚴重,而對照組并沒有明顯差異,說明紫花苜蓿MsHB7基因的異源表達降低了擬南芥對干旱的耐受性(圖6)。
2.5 干旱脅迫下轉MsHB7基因擬南芥生理指標分析
為了進一步研究轉基因和野生型擬南芥的耐旱性,測定了干旱處理后轉基因和野生型擬南芥的相對含水量、脯氨酸(Pro)和丙二醛(MDA)含量。結果顯示,干旱處理后,野生型擬南芥的相對含水量顯著高于轉基因型擬南芥,而脯氨酸和丙二醛含量顯著低于轉基因型擬南芥(圖7)。
2.6 干旱脅迫下抗性相關基因的表達分析
為了研究MsHB7基因對其他抗逆調控相關基因的影響,本研究分析了ATCAT1、ATDREB2A、ATLEA3、ATRD29A這4個基因的表達水平。結果顯示處理之后ATCAT1、ATDREB2A、ATRD29A在轉基因中的表達水平明顯高于野生型,而ATLEA3的表達水平明顯低于野生型(圖8)。

圖3 紫花苜蓿在PEG處理下MsHB7基因的相對表達量分析Fig.3 Analysis of relative expression of MsHB7 gene in alfalfa treated with PEG*和**分別表示同一組織中不同時間點與0 h相比差異顯著(P<0.05)和極顯著(P<0.01)。*and**respectively indicate that the difference between different time points in the same organization and 0 h is significant(P<0.05)and extremely significant(P<0.01).
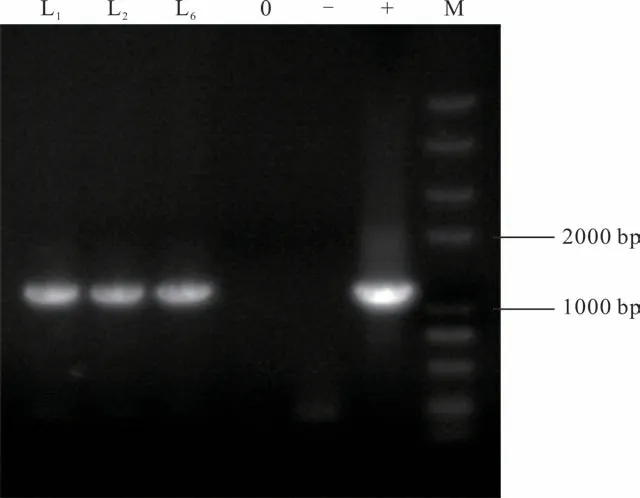
圖4 MsHB7轉基因擬南芥的PCR檢測Fig.4 PCR identification of transgenic A.thaliana with MsHB7M:DNA分子量標準DNA marker;0:水ddH2O;-:陰性對照Negative control;+:陽性對照Positive control.
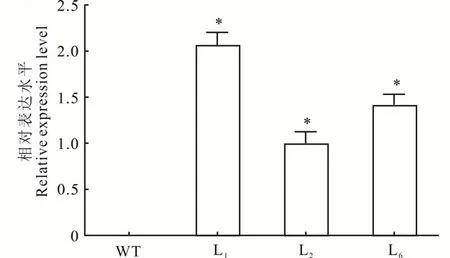
圖5 轉基因擬南芥中MsHB7的相對表達水平Fig.5 The relative expression level of MsHB7 in transgenic A.thaliana*表示轉基因擬南芥與WT相比差異顯著(P<0.05)。*indicates that the difference between transgenic A.thaliana and WT is significant(P<0.05).

圖6 干旱脅迫下擬南芥表型分析Fig.6 Phenotypic analysis of A.thaliana under drought stress

圖7 干旱脅迫下擬南芥生理指標分析Fig.7 Analysis of physiological indexes of A.thaliana under drought stress*表示轉基因擬南芥與WT相比差異顯著(P<0.05),**表示轉基因擬南芥與WT相比差異極顯著(P<0.01)。下同。*indicates that the difference between transgenic A.thaliana and WT is significant(P<0.05);**indicates that the difference between transgenic A.thaliana and WT is extremely significant(P<0.01).The same below.
3 討論
同源異型域-亮氨酸拉鏈(HD-Zip)蛋白是植物特有的一類轉錄因子,目前為止,已經從很多物種中鑒定出了HD-ZipⅠ類亞家族成員,如在擬南芥中鑒定了17個[19],在玉米中鑒定17個[8]。已有研究表明,HD-ZipⅠ亞家族成員與重要的生理過程調控相關,包括植物的生長、發育、衰老以及對非生物和生物脅迫的響應[20]。本實驗從紫花苜蓿中分離出了HD-ZipⅠ類亞家族中的一個成員MsHB7,進化樹分析和多重序列比對表明MsHB7確實為HD-ZipⅠ亞家族中的一員,且與擬南芥中ATHB7和ATHB12親緣關系較近,與其他轉錄因子一樣,MsHB7定位在細胞核上[21-23]。有報道顯示HD-ZipⅠ亞家族基因的表達受外界環境因素的調控,如干旱、極端溫度和滲透脅迫等[5],如擬南芥ATHB7基因的表達受缺水、滲透脅迫以及外源ABA的誘導[10],ATHB12的表達受干旱和ABA誘導[11],玉米干旱誘導基因Zmhdz10也受鹽和ABA的誘導,且在不同的處理時間下表達量不同[24]。在本研究中,MsHB7基因在干旱條件下上調表達,并且地上部分和根中的表達模式存在差異,不同的時間點基因的表達量也不同,推測該基因在對干旱脅迫的響應中存在時間和空間的特異性。為了更好地了解MsHB7基因的功能,本研究利用花序侵染法進行轉化,獲得了MsHB7基因過量表達轉基因擬南芥。
有研究表明HD-ZipⅠ轉錄因子的成員對非生物脅迫起著重要的調控作用,并且對植物耐受性的影響存在差異。其中,一些成員在植物抵御非生物脅迫過程中起正調控作用,如玉米Zmhdz10基因在水稻(Oryza sativa)和擬南芥中的過表達增加了植株對鹽和干旱的耐受性[24];鷹嘴豆CaHDZ12基因在煙草和鷹嘴豆中的過表達也使植株對干旱和鹽脅迫的耐受性增強[20]。另外,還有一些HD-ZipⅠ轉錄因子家族的成員在植物抵御非生物脅迫過程中起負調控的作用,例如在擬南芥中過表達麻瘋樹JcHDZ07基因使轉基因株系對鹽脅迫更敏感[25];水稻HD-ZipⅠ家族基因Oshox22的過表達使植株對干旱的耐受性降低,并且當Oshox22啟動子區域被T-DNA插入突變后水稻的耐旱性增強[26];紫花苜蓿MsHB2基因過表達擬南芥在NaCl和ABA脅迫下,生長被抑制,說明該基因在鹽等非生物脅迫下對紫花苜蓿的抗逆性起負調控作用[27]。本研究中,將野生型和轉基因擬南芥幼苗進行干旱處理18 d后發現,與野生型相比,轉基因擬南芥萎蔫程度更明顯,這表明在擬南芥幼苗中MsHB7基因可能對干旱脅迫起到了負調控的作用,從而降低了擬南芥的耐旱性,這與ATHB7和ATHB12負調控莖的生長類似[11-12]。
此外,本研究還測定了轉基因株系和野生型擬南芥在干旱脅迫后的相對含水量,脯氨酸和丙二醛含量,以研究MsHB7過表達引起的生理生化變化。相對含水量能夠反映植物組織的水分狀況[28],干旱處理后,轉基因型和野生型擬南芥的相對含水量明顯下降,而野生型擬南芥的相對含水量明顯高于轉基因型。在非生物脅迫下植物會被刺激產生活性氧(reactive oxygen species,ROS),膜脂過氧化是活性氧造成的主要影響,一般通過測定丙二醛含量來檢測膜脂過氧化[29]。本實驗對擬南芥進行干旱處理之后,轉基因型擬南芥較野生型積累了更多的丙二醛,這表明MsHB7過表達擬南芥在干旱后產生了更為嚴重的膜脂損傷。脯氨酸被認為是一種滲透保護劑,參與維持氧化還原平衡,活性氧解毒和保護蛋白質結構[30]。在干旱處理后,脯氨酸含量明顯升高,與野生型相比,轉基因擬南芥積累了更多的脯氨酸,推測這是由于干旱后MsHB7的過表達對轉基因植株造成了更為嚴重的損傷,因此需要更多的脯氨酸來維持滲透平衡。
有研究表明,轉基因植株中基因的過表達對不同的活性氧清除酶有不同的影響,例如在干旱脅迫下,ZjZFN1過表達植株中SOD,POD的表達水平下降,而APX的表達水平升高[31];在本研究中,經干旱處理后轉基因株系中ATCAT1的表達量與野生型相比顯著上升。DREB2A蛋白屬于Apetala2/乙烯響應元件結合因子(AP2/ERF)家族,具有耐鹽性和抗旱性[32],許多LEA蛋白是由低溫、滲透脅迫和在營養組織中施加外源ABA誘導的,在擬南芥中DREB1A、DREB2A-CA的過表達或AREB1的激活引起一些LEA基因表達的上調,并且所有的這些轉基因植物都提高了對干旱脅迫的耐受性,這些觀察結果表明,LEA蛋白對水分的缺乏具有耐受性[33]。ATRD29A也被認為是逆境脅迫響應基因。研究表明,ATDREB2A和ATRD29A在轉基因株系中的表達量明顯高于野生型,而晚期胚胎發生豐富蛋白基因ATLEA3在轉基因株系中的表達量則較野生型低。綜上結果表明,MsHB7基因可能直接或間接地調節這些抗逆基因的表達水平,從而調節植物的耐旱性。
4 結論
生物信息學分析表明MsHB7轉錄因子屬于第I類同源異型域-亮氨酸拉鏈蛋白且受干旱誘導。干旱處理18 d后,超表達MsHB7轉基因擬南芥植株與野生型相比萎蔫程度更明顯,野生型擬南芥的相對含水量要顯著高于轉基因株系,并且轉基因株系積累了更多的脯氨酸和丙二醛。此外,在干旱處理之后,4個抗逆相關的基因中,ATCAT1、ATDREB2A和ATRD29A在轉基因中的表達量明顯升高,ATLEA3的表達量明顯下降。干旱脅迫條件下,MsHB7轉錄因子在植物中可能起到了負調控的作用。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
