Slc7a11 基因在黑色素沉積中的研究進展
2021-04-21 06:23:46白少成靳榮帥吳信生
中國畜牧雜志 2021年4期
白少成,靳榮帥,張 琛,姚 凡,陳 陽,吳信生
(揚州大學動物科學與技術學院,江蘇揚州 225000)
黑色素分為真黑色素和褐黑色素,這兩種色素的比例影響著動物的體表和毛發(fā)顏色。一般來說,動物毛色表現(xiàn)出的黑色是通過真黑色素控制,而紅色或黃色是缺乏褐黑色素導致的[1]。溶質載體家族7 成員11(Solute carrier family7 member11,Slc7a11)位于細胞膜上,該基因編碼胱氨酸/谷氨酸xCT 轉運載體,其主要作用是將細胞外的胱氨酸轉運到細胞內,使胱氨酸在細胞內被還原為半胱氨酸[2]。褐黑色素是由多巴向半胱氨酸合成的多巴途徑產生的,而Slc7a11基因通過影響胱氨酸的轉運使細胞內不能攝取足夠的胱氨酸還原為半胱氨酸來滿足褐黑色素的合成,從而造成動物毛色的改變[3]。目前,多方學者探討了Slc7a11與動物黑色素沉積之間的關聯(lián)以及潛在機制,如何軍敏等[4]綿羊和羊駝中發(fā)現(xiàn)該基因在深色皮膚中的表達量較高;穆琳[5]在研究獺兔皮毛黑色素的沉積中證實了Slc7a11的相對功能;Wang 等[6]發(fā)現(xiàn)Slc7a11通過黑色素的生成途徑在紅羅非魚的膚色形成和分化起到重要作用。Slc7a11基因是如何調控黑色素合成的途徑使動物產生五彩斑斕的顏色,仍有待研究。本文將目前Slc7a11基因在動物體內黑色素沉積機制做一綜述,初步分析真黑色素和褐黑色素的合成以及對不同物種毛色形成原因,為探索動物黑色素沉積的分子機制提供新的思路和方法。
1 Slc7a11 基因的結構與功能
Slc7a11 屬于溶質轉運家族(Solute Carrier Family,SLC)中第二大膜蛋白家族,是溶質轉運第7 家族的第11 個成員,位于染色體4q28-q32,CDS 全長1 506 bp(人)和1 509 bp(鼠),編碼近500 個氨基酸[7]。該基因負責編碼氨基酸轉運載體xCT,目前該基因在哺乳動物的研究中多用于黑色素的調控與生成。xCT 是由Slc7a11負責編碼的氨基酸組成的轉運蛋白,作為胱氨酸/谷氨酸反轉運體xc-系統(tǒng)的輕鏈亞基,在哺乳動物體內扮演一種重要的抗氧化蛋白,主要負責特異性轉運胱氨酸或谷氨酸,從而維持細胞內谷胱甘肽(GSH)的水平[8-9]。有研究表明,Slc7a11可增強細胞的抗氧化能力,在氧化應激下通過p53 介導其抑制作用,誘導細胞發(fā)生鐵中毒[9]。Slc7a11基因通過將細胞外的胱氨酸轉運到細胞內,再將細胞內的谷氨酸以相同比例的數(shù)量運輸?shù)郊毎猓源藖砭S持GSH 在細胞內外的水平,進而維持機體內正常黑色素的水平[10]。在功能上,Slc7a11基因已被證明在細胞增殖、氧化應激反應中起重要作用,從而可以作為用于各種疾病治療中的重要手段[11-13]。在毛色方面,Slc7a11基因具有調節(jié)黑色素細胞中氨基酸的轉運功能,從而在黑色素的合成中起重要作用[14-15]。
Slc7a11基因在調控氨基酸的轉運過程中發(fā)揮重要作用,且在大多數(shù)物種中較為保守。何軍敏等[4]采集4 種不同被毛顏色的伊犁哈薩克羊皮膚作為樣品,運用PCR-SSCP 以及測序技術對Slc7a11基因進行多態(tài)性分析,結果在該群體中未發(fā)現(xiàn)多態(tài)性;Tian 等[16]通過克隆羊駝Slc7a11基因的CDS 序列全長并進行序列比對和系統(tǒng)發(fā)育分析顯示,羊駝Slc7a11基因CDS 有1 512 bp,轉錄蛋白包含12 個跨膜區(qū)域的高度保守的氨基酸通透酶,并且確定了羊駝xCT 和野駱駝同一性最高并共享相同的分支都是高度保守。Anello 等[17]同樣在美洲駝中證實Slc7a11編碼具有氨基酸通透酶結構域的高度保守蛋白,并且在其DNA 序列中檢測到7 個SNP,其中6 個為同義,1 個非同義,其中非同義SNP 與顏色的表達無關。此外,陳秋燃等[18]運用SNP 技術檢測不同被毛顏色獺兔Slc7a11基因的12 個外顯子,均沒有發(fā)現(xiàn)多態(tài)性的存在。目前,Slc7a11基因的研究多集中于毛皮動物,其在不同物種間的功能也具有相似性,這進一步說明該基因的編碼序列較為保守。
2 Slc7a11 基因調控褐黑素合成的途徑
在哺乳動物中,動物被毛顏色的改變受黑色素種類和含量不同的影響。黑色素在哺乳動物體內大致分為兩類:一是不含硫原子且主要表現(xiàn)為黑色和棕色的真黑色素,另一種是含硫原子且大多表現(xiàn)為黃/紅色的褐黑色素,兩者在動物體內的比例和分布特征決定了它們的顏色[19-20]。對于褐黑素的合成途徑(圖1),是多巴醌因酪氨酸酶(TYR)活性降低而導致其濃度降低,并與大量的GSH 或半胱氨酸結合形成谷胱甘肽多巴,在谷氨酰轉肽酶的催化下生成半胱氨酸多巴,從而形成褐黑色素[21]。Slc7a11通過調控胱氨酸/谷氨酸的轉運來影響動物毛發(fā)中黑色素的沉積,當該基因發(fā)生大片段缺失后,其編碼的轉運蛋白發(fā)生改變,使其編碼的氨基酸失去轉運功能,導致無法平衡細胞內外的GSH 水平,進而直接作用于多巴醌到半胱氨酸的反應階段來調控褐黑素的合成,最終造成動物毛色的改變[22-24]。
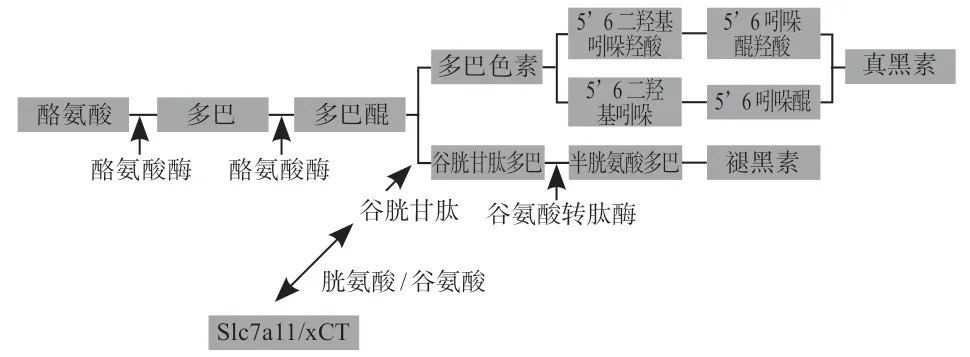
圖1 Slc7a11 基因在褐黑素生成過程中的調控機制
3 Slc7a11 在動物體內黑色素沉積過程中的作用機制
Slc7a11基因在調控氨基酸的轉運過程中發(fā)揮重要作用,其通過為細胞提供半胱氨酸來幫助合成抗氧化劑GSH,從而促進細胞的抗氧化能力[25]。因為GSH 是細胞內主要的抗氧化劑,而半胱氨酸是組成GSH 的3 種氨基酸之一,所以Slc7a11的表達量影響著GSH 的合成和細胞的抗氧化能力[26]。黑色素生成實際上就是一個依賴氧化的過程,在黑色素細胞中,細胞內的半胱氨酸可以進入黑色素小體,并與黑色素合成途徑結合,從而降低半胱氨酸對谷胱甘肽合成的有效性[27]。半胱氨酸作用于黑色素的合成途徑也會影響生物體的色素沉著,因為在哺乳動物的毛發(fā)或禽類的羽毛等表皮結構中沉積時,褐黑素會表現(xiàn)為橙色和淺黃色[28]。黑色素細胞中的褐黑素合成需要消耗GSH,如果黑色素細胞中的半胱氨酸大量用于合成褐黑素,可能會導致黑色素的顏色強度隨著Slc7a11的表達而增加[29]。
3.1Slc7a11基因在黑色素沉積過程中的表達規(guī)律 目前,關于Slc7a11基因調控哺乳動物毛發(fā)顏色的研究多集中于表達規(guī)律的分析。李洪濤等[30]采集2 周齡白色、棕色和黑色哈薩羔羊左肩胛部的皮膚組織,檢測到Slc7a11基因在棕色皮膚的表達量最高,其次是黑色皮膚,且這2 種顏色的羔羊與白色羔羊的Slc7a11基因表達量差異極顯著,同時檢測到xCT 蛋白在哈薩克羊皮膚中的表達量同樣在棕色皮膚中最高,該研究表明Slc7a11基因與棕色哈薩克羊的毛色表型顯著相關;He 等[31]為評估Slc7a11基因在調節(jié)綿羊毛色中的作用,克隆了正常綿羊皮膚成纖維細胞中該基因cDNA 的全長,通過使用睪丸注射轉基因方法獲得了sxCT-轉基因綿羊,并且該綿羊被毛呈現(xiàn)出褐色和黃色的斑點,這表明sxCT可以選擇型表達以增加羊毛中褐黑素的產生,從而驗證了睪丸注射轉基因技術可用于遺傳修飾綿羊毛色的猜想。Tian 等[16]以白色和棕色羊駝為實驗組,檢測這2 種顏色羊駝皮膚組織中mRNA 和xCT 蛋白的表達水平,發(fā)現(xiàn)棕色羊駝皮膚組織中的相對表達量顯著高于白色羊駝,這與李洪濤[30]在哈薩克羊中的研究結果一致;其次,Tian[16]通過免疫組化分析得出,xCT 轉運蛋白的信號分別在毛干、外根鞘、毛囊基質和真皮乳頭中被發(fā)現(xiàn),并且在棕色羊駝皮膚中呈現(xiàn)強陽性,在白色羊駝皮膚組織中明顯下降,該項研究證實Slc7a11基因對羊駝棕色毛發(fā)具有調控作用。除了在羊的種類上有此發(fā)現(xiàn)外,穆琳[5]在獺兔中的研究表明Slc7a11基因的表達量在蛋白青色和蛋白黃色的皮膚組織中較高。以上研究表明,Slc7a11對棕色毛發(fā)和皮膚的調控具有重要意義,進一步證實該基因在調控褐黑素生成過程中發(fā)揮的重要作用。
3.2Slc7a11基因在黑色素沉積過程中的調控機制 在Slc7a11基因調控黑色素沉積機制的研究中,Chintala 等[32]發(fā)現(xiàn)小鼠的淺灰色(sut)突變是由Slc7a11基因的缺失導致,xCT 蛋白缺失導致胱氨酸向黑色素細胞的轉運減少,進而導致細胞內GSH 水平不足以滿足合成褐黑素的需求,阻礙了褐黑素的生成途徑,但真黑素沒有發(fā)生變化,因此小鼠的毛色由黃色變?yōu)榛疑hen 等[33]通過實時PCR 技術檢測Slc7a11在不同毛色獺兔背部皮膚中的表達量,發(fā)現(xiàn)彩色獺兔皮膚中均有Slc7a11表達,其中以黃色獺兔皮膚中的表達量最高,白色最低,進一步利用定點突變技術、雙熒光素酶報告系統(tǒng)和競爭性EMSA 實驗,確認POU2F1 蛋白可結合于Slc7a11啟動子-731~703 bp 區(qū)域調控該基因的啟動子活性,進而影響Slc7a11的表達,參與獺兔的黑色素沉積。Yang 等[34]在對獺兔成纖維細胞中Slc7a11基因的過表達和干擾實驗,發(fā)現(xiàn)當Slc7a11過表達或抑制時,與同樣調控黑色素生成的MITF、MC1R、ASIP、CREB1、Slc24A5相關基因的mRNA 和蛋白表達量呈正相關,這表明Slc7a11可以影響獺兔細胞中其他參與調控黑色素沉積的基因。但于Slc7a11基因的作用機制研究還處于起步階段,作為黑色素沉積的關鍵基因,其上游調控因子及下游作用于黑色素沉積途徑等還有待進一步的挖掘與探究。
4 總結與展望
黑色素在動物體內的沉積對動物毛色的改變起著決定性的作用,所以研究者嘗試通過研究黑色素在動物體內的變化來培育出理想的毛色品系,同時也為人類黑色素沉積相關疾病提供重要靶點。Slc7a11基因作為近年來新發(fā)現(xiàn)的與黑色素沉積相關的研究熱點,它通過調節(jié)胱氨酸/ 谷氨酸的轉運來維持細胞內外正常的GSH 水平,進而影響動物體內褐黑素的形成,最終表現(xiàn)為動物被毛顏色的改變。目前,關于調控黑色素生成的基因中,Slc7a11基因多集中于毛皮動物毛色的研究,對于其中具體的調控機理還有待發(fā)掘。此外,調控黑色素合成的基因之間存在一定的聯(lián)系,但其中具體的作用機制較為空白,深入解析其具體的作用機制有助于進一步探尋動物體內黑色素沉積的規(guī)律及揭示動物毛色形成機理。
