1-磷酸鞘氨醇代謝與急性缺血性腦血管病的研究進展
2021-04-20 02:09:00璞綜述飛審校
中風與神經疾病雜志 2021年3期
關鍵詞:小鼠
劉 璞綜述,高 飛審校
腦卒中(Stroke)是全球第二大死亡原因和第三大致殘原因[1],嚴重危害人類生命健康和生活質量。其中,急性缺血性腦卒中(Acute ischemic stroke,AIS)亦稱急性腦梗死(Acute cerebral infarction,ACI),是最常見的卒中類型,占我國腦卒中的69.6%~70.8%[2]。AIS是指局部腦組織由于急性血液循環障礙,出現缺血、缺氧、壞死,引發局灶性或彌漫性神經功能缺損的疾病,其腦組織損傷的病理生理學過程較為復雜,包括能量衰竭、興奮性毒性、氧化應激和神經炎癥等一系列細胞和分子事件,而神經炎癥的特征又包括中樞神經系統免疫細胞的浸潤、神經膠質細胞(如小膠質細胞和星形膠質細胞)的激活以及多種神經毒性分子(包括促炎細胞因子)的產生,所有這些都會進一步引起中樞神經系統內皮功能障礙、血腦屏障破壞、神經細胞凋亡,從而造成神經功能缺損[3]。
1-磷酸鞘氨醇(Sphingosine 1-phosphase,S1P)是細胞膜鞘磷脂的代謝產物,在介導細胞遷移、增殖、存活和細胞分化等多種生物學效應中起著重要的作用[4]。研究表明小鼠大腦中動脈阻塞(Middle cerebral artery occlussion,MCAO)后3 h,腦組織鞘脂組學分析提示S1P的表達水平積累,而且S1P是唯一在3 h增加的下游代謝物。鞘磷脂在神經細胞中富集,其生物合成和代謝途徑是密切相關的,它們在缺血損傷后的變化可能導致鞘脂類化合物的一種新的穩態狀態。因此對于缺血性損傷后鞘脂變化的系統研究有助于闡明其在缺血性腦卒中病理生理學中的作用[5]。本文將對S1P及其主要相關代謝酶與急性缺血性腦血管病的病理生理學機制的相關性進行歸納,為未來S1P代謝在急性缺血性腦血管病的預防、治療等方面提供理論支持。
1 S1P代謝
S1P是一種來源于細胞膜的生物活性鞘脂,在淋巴細胞轉運、膠質細胞活化、血管收縮、內皮屏障功能調節和神經元死亡途徑中發揮關鍵作用[6]。鞘氨醇(Sphingosine,Sph)是合成S1P的前體底物,由神經酰胺(Ceramide,Cer)在質膜、糖磷脂和鞘磷脂的連續降解過程中水解得到[4]。而S1P的合成是由Sph在限速酶鞘氨醇激酶(Sphingosine Kinase,SphK)的作用下磷酸化而來,SphKs有兩種底物異構體,鞘氨醇激酶1(Sphingosine Kinase1,SphK1)和鞘氨醇激酶2(Sphingosine Kinase2,SphK2)。SphK1主要定位于細胞質并轉移到細胞膜進行激活,而SphK2主要定位于細胞核。在亞細胞結構中,S1P的合成主要是在內質網中啟動,而后續復雜的代謝步驟主要發生于高爾基體中,另外還有溶酶體、細胞核和線粒體的參與[7]。S1P的分解代謝有兩條途徑:①其中一種途徑是S1P被定位于內質網的兩種特異性磷酸酶(S1P-specific phosphatases1 and S1P-specific phosphatases2,SPP1和SPP2)去磷酸化后,通過神經酰胺合成并回收S1P的鞘氨苷部分;②另一種途徑是S1P被1-磷酸鞘氨醇裂解酶(Sphingosine-1-phosphate lyase,SPL)不可逆地降解為磷酸乙醇胺和順-11-十六碳醛,然后被結合到甘油酯中。另外,細胞內的少量S1P被釋放到細胞外環境,這一過程在紅細胞、血小板和內皮細胞中是高效的[4,7]。紅細胞中S1P的輸出可能依賴于ABC轉運蛋白(ATP-binding cassette transporter);血小板以刺激依賴的方式釋放S1P,凝血酶、膠原蛋白、ADP、Ca2+等均可誘導S1P的釋放;而在血管內皮細胞中,細胞內的S1P通過其特異性轉運蛋白Spns2輸出(見圖1)[8]。
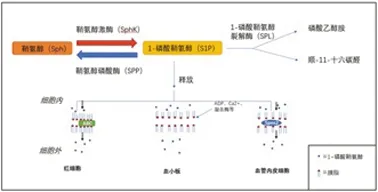
圖1 1-磷酸鞘氨醇代謝示意圖
2 S1P代謝與急性缺血性腦血管病
2.1 S1P與急性缺血性腦血管 血漿中的S1P濃度約為0.9μmol/L,大部分與高密度脂蛋白(約60%)和白蛋白(約35%)結合,少數與其他脂蛋白結合(主要包括低密度脂蛋白和極低密度脂蛋白)[6]。腦組織中的S1P由星形膠質細胞或小腦顆粒細胞分泌,因此S1P可能在中樞神經系統中以自分泌和/或旁分泌的方式發揮作用[9]。在缺血性腦損傷后,S1P的濃度隨時間動態變化。在小鼠MCAO后3 h,腦組織S1P顯著升高,在24 h水平略有下降,而S1P的前體鞘氨醇水平下降。因此,鞘氨醇可能被用于S1P的合成[5]。S1P通過減輕缺血應激下神經母細胞瘤細胞線粒體功能障礙和細胞死亡而增強細胞活力,有可能是通過S1P誘導PKC-ε的線粒體移位,減輕線粒體鈣超載、線粒體通透性轉換孔開放、線粒體內膜電位去極化和腫脹,從而在腦缺血期間提供神經保護[10]。在缺血再灌注(Ischemic-reperfusion,IR)條件下,S1P在腦毛細血管屏障完整性中起著至關重要的作用。IR增強S1P的產生,通過影響緊密連接(Tight junctions,TJs)蛋白的表達增加血管通透性,抑制S1P進入細胞外可改善IR引起的血腦屏障(Blood-brain barrier,BBB)功能障礙[11]。此外,S1P還能有效地調節內皮細胞對損傷的反應以及免疫功能。例如,S1P通過作用于S1PR1激活Gi-PI3K-Akt-eNOS通路維持內皮細胞的完整性、抑制內皮炎癥;相反,S1P作用于S1PR2激活核轉錄因子(Nuclear factor kappa-B,NF-κB)和應激激活蛋白激酶p38途徑,誘導內皮通透性和炎癥[12,13]。S1P通過其相應的受體有效地調節血管內皮的通透性,而內皮通透性改變是炎癥和內皮損傷反應中的關鍵事件。鑒于S1P在調節內皮完整性和內皮炎癥中的重要作用,微血管功能障礙是缺血性腦卒中后重要的一種病理生理狀態,以S1PR為靶點可能成為急性缺血性腦梗死血管保護的新療法[12]。我國已經進行了兩項針對缺血性腦卒中患者的臨床試驗,結果表明單獨或聯合組織纖溶酶原激活物(Tissue plasminogen activator,tPA)使用FTY720(S1P受體激動劑)可降低微血管內皮細胞通透性或保護血腦屏障的內皮完整性,從而保護腦血管并改善缺血性腦卒中預后[14,15]。此外,FTY720還通過阻止淋巴細胞的增殖、激活星形膠質細胞、調節內皮細胞屏障、抑制自噬等多種機制改善腦損傷,是缺血性腦卒中治療的有力候選藥物[16]。研究證實,apoM作為高密度脂蛋白中S1P的載體,介導了HDL-S1P對內皮細胞的保護作用。載脂蛋白apoM和高密度脂蛋白結合的S1P是高密度脂蛋白血管保護作用的主要因素,與apoM結合的S1P為S1PR1提供了一種強直刺激,這對于血管屏障功能至關重要的內皮細胞粘附連接的形成很重要,從而起到血管保護作用[17]。綜上所述,S1P主要通過影響神經細胞線粒體功能障礙減少細胞凋亡、調節血管內皮細胞通透性保護血腦屏障、抑制內皮炎癥,從而在急性缺血性腦血管病腦缺血期間提供神經保護。
2.2 SphK與急性缺血性腦血管病
2.2.1 SphK 鞘氨醇激酶(Sphingosine Kinase,SphK)將鞘氨醇轉化為1-磷酸鞘氨醇(Sphingosine 1-phosphase,S1P),是內源性生成S1P過程中的限速酶。目前已經克隆和鑒定了兩種SphK亞型(SphK1和SphK2),這兩種酶的編碼基因分別定位于17號染色體和19號染色體上,以往的研究表明,這兩種激酶具有重疊的重要功能,單獨敲除SphK1或SphK2的小鼠模型發育正常,但這兩種亞型基因的同時缺失由于血管發生改變和嚴重出血將導致胎鼠死亡[6,18]。盡管這兩種亞型催化同一生化反應,卻表現出不同的亞細胞定位、組織分布和生理功能,SphK1主要存在于細胞質中,且在肺、脾、腎、心臟、腎近端小管和心肌細胞中含量豐富,具有促生存功能;而SphK2主要定位于細胞膜(包括線粒體、內質網和細胞核的細胞膜),且主要分布于大腦,具有抑制生長、促凋亡功能[6,16]。研究發現,以生長因子和存活因子為主的一些生物刺激可激活SphK,從而增加細胞內的S1P濃度,這些刺激包括血小板源性生長因子、神經生長因子、維生素D3、表皮生長因子、TNF-a交聯的免疫球蛋白受體、乙酰膽堿以及G蛋白偶聯受體的配體。然而,這些不同刺激的效果取決于表達各自受體的細胞類型,這可能不僅反映了主要的信號通路,還反映了每個細胞中存在的鞘氨激酶亞型[19]。S1P產生的細胞位置決定了SphK1和SphK2這兩種緊密相關的酶如何利用相同的底物產生相同的產物,從而對細胞生存產生相反的影響,并在調節神經磷脂代謝水平方面發揮相反的作用[9]。
2.2.2 SphK1與急性缺血性腦血管病 激活小膠質細胞中的SphK1對于腦缺血再灌注(Cerebral ischemia-reperfusion,CIR)損傷的神經炎癥具有重要意義。研究發現在小鼠MCAO模型中,1 h后的CIR導致明顯的梗死損傷。與非缺血、對側皮質或假手術小鼠皮質的SphK1 mRNA相比,1 h后的MCAO再灌注半暗帶皮質的SphK1 mRNA顯著增加,并在再灌注6 h開始增加,并持續至再灌注后96 h。有趣的是,1 h后MCAO再灌注皮質半暗區SphK2的mRNA水平并沒有改變。免疫組化結果顯示,SphK1在小鼠MCAO再灌注24 h后的表達,主要與小膠質細胞標記物和皮質半暗帶神經元標記物共定位,提示小膠質細胞和神經元是腦缺血后SphK1誘導的主要細胞來源[20]。Manhua等[21]研究發現SphK1在CIR損傷的小膠質細胞中表達增高,并通過調節激活的小膠質細胞中IL-17A的表達,誘導CIR過程中神經元的凋亡。而抑制SphK1的表達和敲除SphK1基因可以抑制小膠質細胞介導的缺血狀態下的神經炎癥從而減輕CIR[20]。另一項研究表明過氧化氫誘導的CIR損傷后SphK1的轉錄和表達水平升高。增加的SphK1通過激活NF-κB信號通路,促進內質網應激并誘導炎癥反應,而過度內質網應激和炎癥反應進一步觸發神經元死亡,從而加重CIR損傷。相反地,SphK1的缺乏減輕CIR損傷的內質網應激和炎癥反應[22]。也有研究表明在MCAO后再灌注24 h,SphK1在梗死區皮質的表達明顯減少,但在梗死區周圍皮質保留,且部分神經元共同表達[23]。一般認為,缺血損傷核心的大部分細胞在MCAO后再灌注24 h內死亡,MCAO后再灌注24 h SphK1表達的降低可能反映了死亡細胞中細胞蛋白的普遍降解。因此SphK1在皮質半暗區表達的誘導,可能是一個由潛在的可存活的和可逆損傷的組織組成的區域,被認為是中風治療的一個目標區域[20,24]。綜上所述,SphK1主要通過激活小膠質細胞誘導神經元凋亡,從而在急性缺血性腦血管病腦缺血期間介導神經炎癥反應。
2.2.3 SphK2與急性缺血性腦血管病 缺氧預處理(Hypoxic Preconditioning,HPC)可顯著而短暫地提高腦微血管SphK2蛋白的表達和活性,Wacker等研究發現SphK2調節血腦屏障中的連接蛋白,并參與缺氧預處理誘導的缺血耐受。在小鼠短暫MCAO后,SphK2基因的缺失增加了腦缺血梗死面積,并導致神經功能惡化,這表明SphK2參與了腦缺血損傷后的神經保護[25]。Pfeilschifter等[26]在小鼠瞬時大腦中動脈閉塞(Transient middle cerebral artery occlusion,tMCAO)模型的研究中顯示,SphK2基因敲除的小鼠在再灌注24 h后出現的缺血性病變要比野生型小鼠大25%,且同時伴有更嚴重的神經功能缺損。此外,tMCAO后鞘氨醇模擬物FTY720的神經保護作用被SphK2基因的缺失所消除,認為SphK2特異性磷酸化的鞘氨醇模擬物FTY720在嚙齒類動物缺血性卒中模型中以依賴Sphk2的方式發揮保護作用[27]。在異氟醚預處理(Isoflurane preconditioning,IsoPC)的小鼠中,預處理小鼠皮質SphK2 mRNA和SphK2蛋白水平迅速上調,且隨時間呈動態變化。在小鼠MCAO后再灌注24 h,上調的SphK2表達仍然是對照組的2.2倍,而腦SphK1 mRNA和蛋白水平在IsoPC后不同時間點的表達無變化。相反地,SphK2抑制劑ABC294640預處理阻斷了IsoPC的保護作用,即阻斷了IsoPC對小鼠腦梗死體積的減少和神經功能評分的改善,這可能是由于SphK的抑制抑制了原代神經元中異氟醚誘導的缺血耐受,神經元SphK2的上調和激活在腦預處理中至關重要,通過減少神經元細胞凋亡保護大腦免受缺血性損傷[28]。研究表明預處理刺激通過SphK2誘導小鼠皮質神經元自噬,而自噬反過來又有助于IsoPC和HPC誘導的皮質神經元的神經保護作用[29],其機制認為是預處理相關的SphK2上調激活自噬,通過其BH3結構域與Bcl-2相互作用,從而將其與Beclin-1解離,保護神經元免受缺血性損傷[30]。此外,SphK2在巨噬細胞的自噬體和溶酶體的脂質分解代謝中起關鍵作用,并在動脈粥樣硬化形成中起保護作用。SphK2通過促進自噬脂質降解防止巨噬細胞膽固醇積聚和動脈粥樣硬化,而SphK2缺乏引起的鞘脂代謝紊亂會損害巨噬細胞的自噬體-溶酶體功能,導致巨噬細胞中脂質的過度積累和動脈粥樣硬化的加重。因此,SphK2可能是治療動脈粥樣硬化的新靶點[31]。綜上所述,SphK2主要通過調節血腦屏障中的連接蛋白、減少神經元細胞凋亡,從而在急性缺血性腦血管病腦缺血期間提供神經保護。
3 結 語
綜上所述,S1P及SphK與急性缺血性腦血管病的神經功能缺損程度密切相關,近年來,S1P及SphK在神經系統疾病,尤其腦血管疾病的發生、發展的相關正在全世界范圍內深入開展,其在細胞學中的生物學效應已經得到了初步證實,主要涉及促進神經元細胞增殖、誘導神經細胞凋亡、調節內皮細胞通透性、促進或抑制神經炎癥反應的作用。因此,通過對S1P代謝與急性缺血性腦血管病病理生理學機制相關性的研究,可以為未來急性缺血性腦血管病的治療提供新的方向。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
