巴西橡膠樹赤霉素信號轉導途徑HbRGA1結構和功能分析
2021-04-19 08:36:16陳華峰樊松樂王立豐
植物研究 2021年4期
陳華峰 樊松樂 王立豐*
(1. 東北林業大學化學化工與資源利用學院,森林植物生態學教育部重點實驗室,哈爾濱 150040;2. 農業農村部橡膠樹生物學與遺傳資源利用重點實驗室,省部共建國家重點實驗室培育基地—海南省熱帶作物栽培生理學重點實驗室,農業農村部儋州熱帶作物科學觀測實驗站,中國熱帶農業科學院橡膠研究所,海口 571101)
赤霉素信號轉導途徑是赤霉素和它的可溶性受 體 蛋 白GID1 結 合 時,GID1 與DELLA 蛋 白 的DELLA 和VHYNP 結合后,將阻遏蛋白DELLA 在26S 蛋白酶體水解[1]。DELLAs 是陸生植物特有的,屬于GRAS 轉錄調控子家族[2]。它是赤霉素信號的負調控因子。缺失該基因的擬南芥和水稻突變體會產生促進生長發育的表型。DELLA 功能與其結構、翻譯后修飾、下游轉錄調控靶基因和蛋白互作相關[3]。在模式植物擬南芥中已經鑒定了GAI、RGA、RGL1、RGL2 和RGL3 五個DELLA[4]。其N 端和C 端分別含有DELLA 和GRAS 超級家族結構域。在其功能研究領域,已證明DELLA 通過蛋白互作方式調控超過300 以上的轉錄因子[2]。擬南芥DELLA 蛋白與MYB21 和MYB24 結合調控花絲伸長[5]。FKF1 蛋白負調控DELLA 蛋白穩定性促進開花[6]。油菜BnaA6.RGA 與ABA 信號轉錄因子ABF綜和調控干旱抗性[7]。鑒于DELLA 蛋白的重要性,研究人員相繼從海棠[8],蘋果[9]等果樹,大戟科(Euphorbiaceae)植物珍珠黃楊[10]和蓖麻[11]中克隆并鑒定了DELLA蛋白基因并驗證其功能。
巴西橡膠樹(Hevea brasiliensis)起源于南美亞馬遜流域,是重要工業原料天然橡膠的主要來源[12]。橡膠樹生長、發育和產量與植膠區環境和植物激素信號密切相關[13]。隨著分子生物學在橡膠樹研究領域的不斷拓展,橡膠樹中植物激素信號的調控作用機制不斷更新。如揭示橡膠樹ABA信號途徑的PP2A 家族[14],bZIP[15]。生長素信號HbJAR1[16]和乙烯信號AP2/ERF[17]等。在橡膠樹赤霉素信號研究領域,已經鑒定了HbGAI 基因,并證明其受割膠、茉莉酸甲酯和乙烯利差異調控表達。在RGL1 的研究領域,發現RGL1 與WRKY45互作激發葉片衰老[18]。鑒于DELLA 蛋白植物生長發育和激素信號交互中的作用,揭示并證明橡膠樹中DELLA 蛋白的結構與功能將為研究生長發育和產量形成提供堅實的理論基礎。據此,本研究克隆并鑒定HbRGA1 的cDNA 全長序列,利用生物信息學分析預測該基因及其推導的氨基酸序列的結構和特性,并利用熒光定量PCR 技術分析該基因的表達模式,為闡明HbRGA1在橡膠樹的生物學功能打下基礎。
1 材料與方法
1.1 實驗材料
采用中國熱帶農業科學院橡膠研究所儋州基地國家種質資源圃種植巴西橡膠樹主推品種Rey?an73397 正常割膠的10 年樹齡的莖、葉、膠乳、樹皮為材料,用于HbRGA1 的克隆、不同組織的表達分析。白粉病、機械傷害、赤霉素、干旱、脫落酸、水楊酸、茉莉酸甲酯、生長素和乙烯利處理參照文獻方法[19~20]。
1.2 實驗方法
1.2.1 橡膠樹葉片提取總RNA和反轉錄成cDNA
參照TIANGEN 植物總RNA 提取試劑盒的方法,提取巴西橡膠樹不同組織和不同處理樣品的總RNA,RNA 的濃度與純度通過Thermo Fisher NanoDrop 2000 超微量核酸蛋白分析儀(Gene Company Limited,上海)檢測,后利用TaKaRa 反轉錄試劑盒發轉錄獲得cDNA,以cDNA 為模板利用PCR 擴 增 內 參 基 因 HbACTIN(Genbank:HQ260674.1)進行驗證。
1.2.2 HbRGA1 cDNA全長序列的克隆
根據橡膠樹數據庫的序列設計引物(見表1)以熱研73397 的cDNA 為模板,利用PCR 擴增HbRGA1 的全長序列,使用OMEGA 膠回收試劑盒并參考使用說明對PCR 產物進行回收,連接pMD-18T 載體后轉化大腸桿菌DH5α 感受態細胞中,在含氨芐抗性培養基上培養,檢測后挑取陽性克隆送去賽默飛世爾公司測序。

表1 HbRGA1全長擴增和熒光定量引物序列Table 1 Primer sequences of full-length cloning and qRTPCR of HbRGA1
1.2.3 HbRGA1生物信息學分析及進化分析
利用在線工具SignalP-5.0 Server(http://www.cbs.dtu.dk/services/SignalP/)分析信號肽,TMHMM Server v. 2.0(http://www. cbs. dtu. dk/services/TMHMM/)預測跨膜結構,DeepLoc-1.0(http://www.cbs.dtu.dk/services/DeepLoc/)預測亞細胞定位,PSIPRED V4.09 (https://github. com/psipred/psipred)預 測 二 級 結 構,SWISS-MODEL(https://swissmodel.expasy.org/)預測三級結構,SMART:Main page(http://smart. embl-heidelberg. de/smart/set_mode.cgi?NORMAL=1)分析保守結構域,Prot?Param(https://web.expasy.org/protparam/)工具對其理化性質進行生物信息學預測分析。MEME(http://meme-suite.org/tools/meme)在線分析軟件對與HbRGA1同源性較高的序列經行motif預測分析。使用DNAMAN9 軟件經行多序列對比及同源相似性。使用MEGAX 軟件并使用鄰接法(neigh?bor-joining)經行系統發育分析(bootstrap 值設為1 000)。
1.2.4 不同處理下HbRGA1表達分析
根據HbRGA1 編碼蛋白序列設計熒光定量引物(見表1),選用橡膠樹HbACTIN 基因為內參,采用SYBR Green 法在伯樂熒光定量PCR 儀進行定量PCR 檢測,熒光定量PCR 反應體系和程序參考前人方法[21]。
1.2.5 數據統計分析
Excel 進行數據整理分析,IBM SPSS Statistics進行單因素ANOVA 檢驗分析差異顯著性,Origin Pro2018(Origin Lab Corporation,Massachusetts,USA)軟件進行作圖。
2 結果與分析
2.1 HbRGA1的克隆與生物信息學分析
以橡膠樹葉片cDNA 為模板,通過PCR 克隆得到HbRGA1 cDNA 全長序列,測序驗證正確無誤后,并將cDNA 序列命名為HbRGA1 并提交NCBI(GenBank:KM086713)。其長度2 136 bp,包含1 839 bp的ORF。HbRGA1基因編碼區的核苷酸和推導的氨基酸序列,共編碼613 個氨基酸殘基,在49-115是特征性的DELLA蛋白結構域,在254-610處是GRAS 結構域(見圖1A)。通過在線分析工具SignalP-5.0 Server(http://www.cbs.dtu.dk/services/SignalP/)分析預測HbRGA1 存在信號肽的概率是0.002(見圖1B),說明其不含信號肽。利用在線分析 工 具PSIPRED V4.0(http://bioinf.cs.ucl.ac.uk/psipred/)預測HbRGA1 蛋白的二級結構(見圖3A)發現存在許多Alpha Helix 和Random coil 結構,利用SWISS-MODEL(https://swissmodel.expasy.org/in?teractive)在線分析工具分析其三級結構與二級結構預測結果相符合(見圖1B)。利用ProtParam(https://web.expasy.org/protparam/)工具在線分析蛋白質的理化性質(見表1),證明其實疏水性蛋白。HbRGA1 與毛果楊PtPOPTR(Populus tricho?carpa,XP_002305198),蓖麻RcGAI(Ricinus com?munis,XP_002534030),胡 楊PeGAI(Populus eu?phratica,XP_011002785),蘋果MdGAI-like(Malus domestica,XP_008343058),橡膠樹HbGAI(Hevea brasiliensis)進行同源性分析,總相似度達到82.5%9(見圖2)。進化分析表明HbRGA1 與橡膠樹HbGAI 聚為一類(見圖3A)。TMHMM Server v.2.0(http://www.cbs.dtu.dk/services/TMHMM-2.0/)分析預測HbRGA1 無跨膜蛋白結構,利用在線分析工具DeepLoc-1.0(http://www.cbs.dtu.dk/services/DeepLoc/)預測HbRGA1 蛋白亞細胞定位,發現定位于細胞核中的概率為0.998(見圖3B)。利用MEME 在線分析軟件分析橡膠樹HbRGA1 與其他植物DELLA 蛋白序列有3 個共同的motif(見圖3C)且標注了其在相關序列的位置,motif序列與生物功能密切相關。
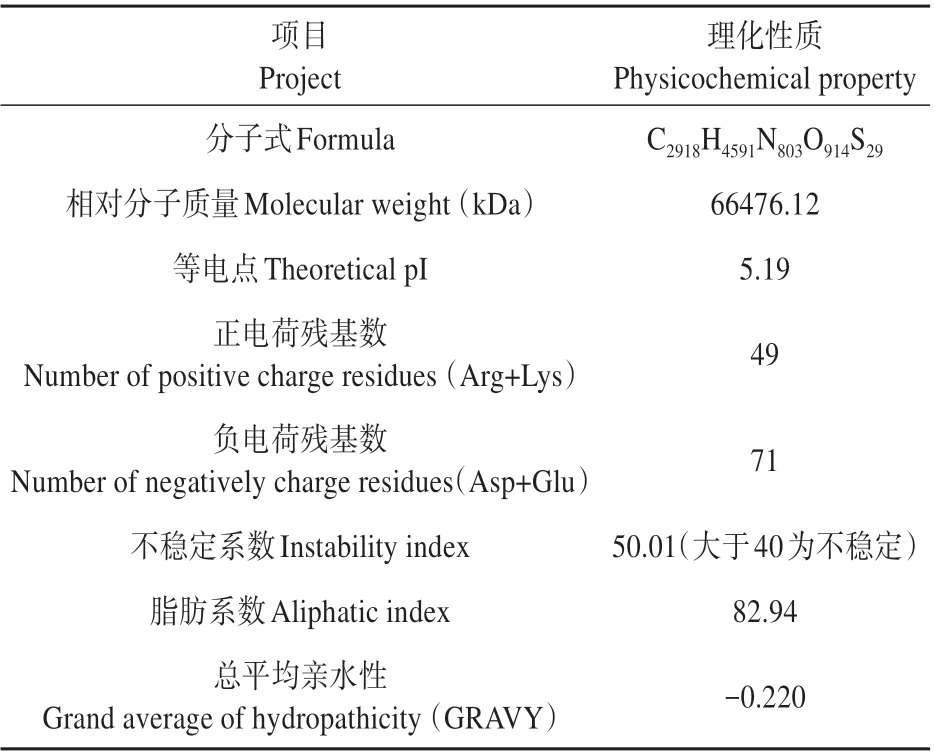
表2 橡膠樹HbRGA1蛋白質理化性質Table 2 Physical properties of HbRGA1 from H. brasil‐iensis
2.2 HbRGA1的表達分析
通過分析實時熒光定量PCR 數據發現,HbRGA1 在橡膠樹樹皮、葉片、膠乳和花中均有表達,其中在葉片和花中的表達量較高,葉片的表達量是膠乳中的8倍左右。在樹皮和膠乳中HbRGA1的表達量相對較低(見圖4)。據此,我們以橡膠樹芽接苗為材料進一步分析了在干旱和不同激素處理下的表達規律。從圖4可以看出,隨著白粉病級別的提高,HbRGA1 的表達量呈持續上升趨勢。機械傷害處理的葉片,HbRGA1 的表達量分別在1 和10 h 達到兩個峰值,分別為初始0.5 h 的4 和6 倍。赤霉素處理后,HbRGA1的表達量在0.5 h就上調接近12倍,隨后呈現下降的規律。
在干旱條件下,HbRGA1 基因的表達量在3 d顯著性上升,達到對照的6 倍,之后表達量呈下調趨勢。在ABA 作用下,HbRGA1 基因的表達量在0.5~10 h 顯著性上調且在10h 達到最高點,之后下調,但仍高于處理前的水平。水楊酸處理后,HbRGA1 的表達量在10~48 h 長時間保持峰值,隨后下調。茉莉酸甲酯處理后,HbRGA1 的表達量呈現先上升后下降的規律,在6 h 達到峰值,表達量是對照的6 倍。生長素處理后,HbRGA1 的表達量在6~48 h 呈現峰值,表達量是對照的6 倍。乙烯利噴施處理的葉片中,HbRGA1 基因的表達量在6 h呈現峰值,隨后急劇下調并趨于穩定(見圖5)。
3 討論
3.1 HbRGA1是植物DELLA蛋白家族成員
DELLA 蛋白是植物赤霉素信號的轉錄遏制因子,含有特征性的DELLA 和GRAS 超級家族結構域。DELLA 結構域的序列為DELLAVLGYKVRSSDMADVAQKLEQLEMVMGTAQEDGISYLCSDT ?VHYNPSDLSGWVQSMLSELNPPMCLDASG,在 所有植物物種中均保守,DELLA 和VHYNP這兩個基序是赤霉素信號受體GID1 蛋白的結合區域。采用缺段突變技術,將RGA 基因的DELLAVL?GYKVRSSEMA 敲除后,突變體擬南芥表現出GAI一樣的表型,說明DELLA 結構域是DELLA 蛋白行駛功能的重要區域[22]。DELLA 蛋白家族的另外一個結構域是GRAS 也具有DELLA 蛋白的功能,在缺失DELLA 結構域的突變體發揮部分替代功能[23~24]。本研究發現,我們克隆的HbRGA1蛋白分別在49-115 是特征性的DELLA 蛋白結構域,在254-610 處是GRAS 結構域(見圖1A),并與其他植物已知的DELLA 蛋白相似度達82.5%以上(見圖2),并與橡膠樹HbGAI 蛋白聚在一起,說明HbRGA1是植物DELLA 蛋白家族中一員。亞細胞定位顯示其定位在細胞核,與F-box 蛋白SLY1 和GID1蛋白等互作[25]。
3.2 HbRGA1 在橡膠樹植物激素信號交互中具有重要作用
盡管DELLA 蛋白最早是在赤霉素激素信號研究中發現的,但其被證明參與多種生長發育過程和其他植物激素信號途徑[26~27]。例如,擬南芥中乙烯通過調控DELLA 抑制赤霉素生長效應,進而調控擬南芥生長發育[28]。生長素單獨調控DELLA 蛋白進而調控赤霉素水平[29]。在茉莉酸信號中,DELLA 蛋白通過與茉莉酸信號阻遏子JAZ蛋白競爭性結合參與調控茉莉酸信號[30]。在植物逆境抗性,冷害誘導因子CBF1 通過調控DELLA蛋白含量調控赤霉素代謝[31]。遠紅光通過DELLA蛋白刺激的ABA合成抑制萌發[32]。這與本研究的結果一致。在單獨逆境和激素處理下,都能上調HbRGA1 的表達量。其中,赤霉素誘導效果最早,在0.5 h 達到峰值(見圖4)。干旱、水楊酸和生長素誘導效應最久,可達2~3 天(見圖5)。這說明,HbRGA1 在橡膠樹中可能參與多種植物激素信號轉導過程的交互。這與前人的研究結果一致,如HbGAI 受茉莉酸甲酯和乙烯利誘導差異表達[33]。蓖麻RcGAI 在葉片中表達量高[34]。下一步擬采取構建過表達載體轉基因,基因編輯[35]和免疫印跡技術鑒定hbRGA1的互作蛋白和功能[36]。
3.3 HbRGA1 在橡膠樹排膠機理與調控技術研究中的應用
巴西橡膠樹膠乳來自于位于巴西橡膠樹韌皮部的乳管細胞,里面含有線粒體、質體等細胞器,具有割膠后膠乳再生的作用。不同品種具有不同的排膠特性[37]。這些排膠特性的差異是由于里面的差異基因表達決定的[37]。挖掘橡膠排膠的關鍵功能基因及相關分子標記是天然橡膠產業亟待解決的理論和技術問題。目前,已經證明乙烯通過延長排膠時間促進膠乳產量提升[39]是由于Hev b 7-like 蛋白起到的去凝集作用[40]。作為赤霉素和乙烯激素信號交互的重要調控蛋白[28],DELLA 蛋白精細的調控兩種激素介導的生長發育平衡。本研究發現,HbRGA1 在赤霉素處理前期0.5 h 高表達,隨后顯著下調表達。乙烯利處理在6 h 顯著上調HbRGA1 表達。表明激素信號可能通過時間或空間差異調控DELLA 蛋白基因表達進而調控阻遏蛋白含量,抑制赤霉素信號轉導。可采用分析不同品種、不同排膠時間的膠乳代謝組和蛋白組聯合分析鑒定HbRGA1 在橡膠樹排膠過程中的作用,為研發排膠調控技術提供指導。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
電子制作(2018年18期)2018-11-14 01:48:24
電子制作(2018年11期)2018-08-04 03:25:42
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
