青藏苔草形態特征及生物量分配沿水分梯度的變化
2021-04-09 03:35:28王洪斌張煜坤張大才
草地學報 2021年3期
王洪斌, 張煜坤, 張大才
(國家林業和草原局西南地區生物多樣性保育重點實驗室, 西南林業大學林學院, 云南 昆明 650224)
濕地生態系統具有維持生物多樣性、調節氣候、涵養水源和保持區域生態平衡等多種功能,被稱為“地球之腎”[1]。中國濕地類型多樣、面積寬廣,但大部分濕地面臨過度放牧、污染和圍墾等威脅[2-4],在氣候干暖化及放牧等因素的長期影響下,沼澤濕地逐漸演替為沼澤化草甸、草甸、退化草甸甚至沙化草甸[5-6]。川西高原作為長江、黃河上游及其主要支流的源頭,分布有大面積的沼澤濕地,生態戰略地位十分重要[7-8],但濕地的退化十分嚴重,且退化過程由水分降低驅動[9-10]。因此,研究濕地植物對水分的響應對于沼澤濕地生態系統保護具有重要意義。
植物在適應異質生境時,功能器官的強弱變化與形態特征的分化能力常被稱為表型可塑性,其強弱關系著植物對逆境的適應能力以及其在植物群落中重要值的大小[11-12]。植物可塑性越強,對生境的適應能力也越強[13]。在干旱化生境中,植物的生長高度、莖葉性狀、克隆構件、根系生長及生物量都會發生相應的變化[14]。例如,華扁穗草(Blysmussinocompressus)地上、地下生物量都隨土壤水分的減少而增加,干旱條件反而有利于其生物量的積累[15];高山嵩草(Kobresiapygmaea)在適應干旱化生境時,呈現出分株和根系數量及長度增加、葉片數量和莖基直徑降低的形態變化,而矮生嵩草(K.humilis)會采取增加根系長度、減少分株數量和葉片數量同時減小莖基直徑的適應策略[16]。此外,盡管植物器官形態特征之間存在顯著的相關性,但對環境變化響應的敏感度存在差異[17]。有研究指出,旱生植物營養器官中,葉與植物生長密切相關,與外界環境進行物質能量交換最頻繁,最容易發生定向進化,是可塑性最大的營養器官[18-19]。生境干旱化過程中,根系生長量增加可以增強水分的吸收,而葉片數量減小可以減小蒸騰作用[15-16],但濕地植物以怎樣的機制響應生境干旱化以及其葉的可塑性是否也最強還尚待研究。
青藏苔草(Carexmoorcroftii)是川西高原高寒草甸和沼澤濕地的優勢種,海拔分布范圍為3 400~5 700 m[20],在水土保持、維持生態平衡方面起著重要作用[21-22]。然而,隨著沼澤地水位的下降和高寒草甸生境干旱化的加劇,青藏苔草在植物群落中的重要值和生態位寬度逐漸下降[23]。沼澤與草甸的退化過程大多是土壤含水率降低的生境干旱化過程[24-25],目前,對于青藏苔草的相關研究主要在于遺傳多樣性[20]、生態位特征[21]、地理分布[26]及葉的形態特征隨海拔梯度的變化[27]等方面,而少有研究涉及其根、莖、葉形態特征及生物量對土壤水分梯度的響應。本文以青藏苔草為研究對象,通過研究其根、莖、葉形態特征變化和生物量分配,以揭示青藏苔草營養器官形態特征沿土壤水分梯度的變化與營養器官形態特征可塑性的強弱排序,以及生境干旱化過程中植株個體生物量的分配策略。
1 材料與方法
1.1 研究地概況
研究地點位于四川省甘孜州新龍縣銀多鄉古冰帽沼澤濕地,地理坐標為31°24′25″ N,99°49′48″ E,海拔4 300~4 450 m。據中國天氣網(www.weather.com.cn)2009—2018年的資料,該區域屬于青藏高原亞濕潤氣候區,年均降水量631 mm,年均氣溫7.9℃,年均最高氣溫18.3℃,年均最低氣溫1℃,全年60%以上降水量集中在6—8月。沼澤化草甸以青藏苔草為單優勢種,高寒草甸以矮生嵩草、大花嵩草(K.macrantha)為共優種,伴生有青藏苔草、黑褐穗苔草(Carexatrofusca‘Minor’)、高山早熟禾(Poaalpina)、極地早熟禾(P.arctica)、絲穎針茅(Stipacapillacea)等。在放牧干擾下,高寒草甸退化嚴重,藍玉簪龍膽(Gentianaveitchiorum)、大萼藍鐘花(Cyananthusmacrocalyx)、川西小黃菊(Tanacetumtatsienense)、星狀風毛菊(Saussureastella)、高山唐松草(Thalictrumalpinum)、高山大戟(Euphorbiastracheyi)、委陵菜(Potentillachinensis‘Chinensis’)、珠芽蓼(Polygonumviviparum)、圓穗蓼(P.macrophyllum)等植物在干旱生境中常見,并在部分區域形成優勢種。
1.2 樣地設置與采樣
樣地位于兩山之間的平緩地段,低凹區域為大面積沼澤水域,淺水區域分布有青藏苔草植物群落,高寒草甸分布于平緩坡面上(圖1)。從水域開始,沿著距水域距離增加的方向,根據TSC-IW型土壤水分速測儀測量的土壤含水率,并結合青藏苔草的分布范圍共設置5個4 m×4 m樣地,其中2個位于有積水的沼澤化草甸,3個位于高寒草甸。青藏苔草在5個樣地中均有分布,但各樣地中個體的空間分布格局不盡相同:1號樣地為青藏苔草沼澤化草甸,群落蓋度90%以上,幾乎沒有其它植物分布,青藏苔草植株呈均勻分布;2~5號樣地,青藏苔草植株空間分布格局逐漸從集群分布過渡到隨機的單株分布,5號樣地為青藏苔草分布的邊緣,向樣地外延伸幾乎無青藏苔草植株分布(表1)。
為保證根系取樣的完整性,采用挖掘法采樣,對草地根系的研究表明,80%的根系分布于0~10 cm土層[28-29],因此挖取土塊時,深度均為20 cm以上。樣地中具有花、果的青藏苔草植株很少,部分樣地甚至沒有,處于花果期的植株莖會高出葉一些,因此選擇莖比葉片矮(即花芽還未抽出)的植株做為研究對象。在1~2號樣地,青藏苔草分布密集,蓋度極高且生長狀況均一,設置2個1 m×1 m樣方挖取一定體積的土塊,即可采集到30株以上青藏苔草;3~5號樣地青藏苔草散生分布,蓋度很低,各植株間距離較遠且生長狀況各不相同,設置3~6個1 m×1 m樣方隨機采集青藏苔草30株以上。將樣品裝入塑料袋,帶回室內進行分株并測定地上、地下指標。采樣時間為2018年8月。

圖1 樣地設置示意圖Fig.1 Sketch map of sample plots setting
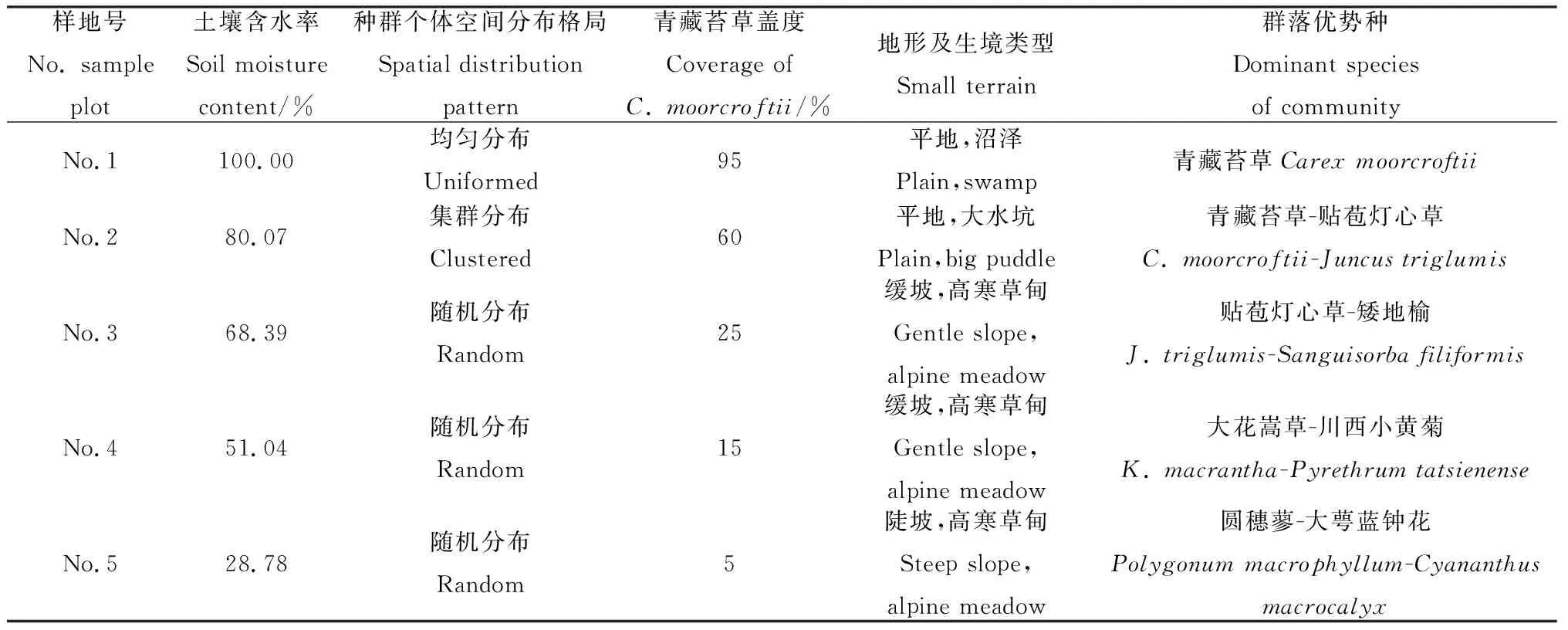
表1 樣地信息Table 1 Information of sample plot
1.3 數據測量
1.3.1土壤含水率的測定 使用TSC-IW型土壤水分速測儀測量樣地的土壤體積含水率,在干旱季節(9月底)測量,每個樣地沿樣地邊界及對角線選取10個點測量土壤體積含水率,以平均值衡量每個樣地的土壤含水率(表1)。
1.3.2植株形態特征的測量 將所有植株從土壤中完整地分離出來,每個樣地選取長勢相對均一的植株30株,分別統計葉片、殘存葉基和分蘗芽數量,用游標卡尺測量植株莖基、根狀莖和分蘗芽直徑(mm),用精度為0.1 cm的直尺測量最長三片葉的長度(取平均值代表每株葉片的長度)和根狀莖長度(cm)。完成莖、葉形態數據測量后,將根系分離出來,在尼龍網中將根系沖洗干凈,采用TTC染色法區分出活根與死根,去除死根,將分離出的活根系鋪開,使用Epson Perfection V700根系掃描儀進行掃描,并用配套的WinRHIZO根系分析軟件獲取各樣地根系的總根長(cm)、表面積(cm2)、平均直徑(cm)、體積(cm3)數據。
1.3.3生物量的測定 將每個樣地中青藏苔草植株的地下、地上部分分裝于不同的信封,在105℃下殺青30分鐘后,于65℃下烘干48 h至恒重,用電子天平稱量每個樣地植株的地下(Belowground biomass,BB)、地上(Aboveground biomass,AB)生物量(g),并計算BB/AB值。
1.4 數據分析
獲取的數據使用Excel軟件記錄并整理后,使用SPSS 25.0軟件分析根、莖、葉形態特征及生物量與土壤含水率之間的關系。使用Origin 2019b軟件進行繪圖。
使用Canoco 4.5軟件分析青藏苔草形態特征與土壤含水率之間的排序關系,根據去趨勢對應分析(DCA)結果中“Length of gradient”判別排序模型的選擇[30]。本研究中,“Length of gradient”最大值為0.26,選擇RDA線性排序模型,將排序圖第一軸,即橫軸(—1,1)均等劃分為3段,作為劃分可塑性強弱等級的依據。第1段為(—0.333,0.333),可塑性弱;第2段為(—0.666,—0.333)&(0.333,0.666),可塑性中等;第3段為(—1,—0.666)&(0.666,1),可塑強。
2 結果與分析
2.1 青藏苔草形態特征與生物量對水分的響應
青藏苔草的分蘗芽直徑、根系平均直徑、總根體積與葉長、根系表面積、地下生物量、地上生物量分別在生境較濕潤的1號和2號樣地中達到最大值,殘存葉基數量、總根長、根狀莖長度與葉片數量、莖基直徑、分蘗芽數量分別在生境較干旱的4號和5號樣地中達到最大值,而根狀莖直徑則在3號樣地中達到最大值(表2)。青藏苔草根、莖、葉形態特征在不同水分梯度中存在顯著差異(P< 0.05,表2)。
2.2 形態特征與土壤含水率之間的關系
隨著生境干旱化,青藏苔草根系表面積、根系平均直徑、總根體積均呈減小趨勢(圖2A,B,C),而總根長呈增大趨勢(圖2D)。與最濕潤生境樣地(No. 1)相比,最干旱生境樣地(No. 5)總根長增加了55.62%,根系表面積、根系平均直徑和總根體積分別減小了25.49%,54.71%和63.37%。在生境干旱條件下,根系變細變長有利于青藏苔草吸收、利用深層土壤水分。
青藏苔草葉長隨土壤含水率降低而減小(圖3A),葉片數量、殘存葉基數量、莖基直徑均隨土壤含水率降低而增大(圖3B,C,D)。葉片縮短、數量增多是青藏苔草在干旱化生境中維持光合產出的適應策略。
青藏苔草分蘗芽數量、根狀莖長度隨土壤含水率降低呈增大趨勢(圖4A,B),分蘗芽和根狀莖直徑均隨土壤含水率降低而減小(圖4C,D)。克隆構件營養物質的分配存在權衡關系,隨著土壤含水率降低,根狀莖形態趨向更細、更長,分蘗芽趨向數量更多而形態更小,朝著占據更多生存空間的方向發展。
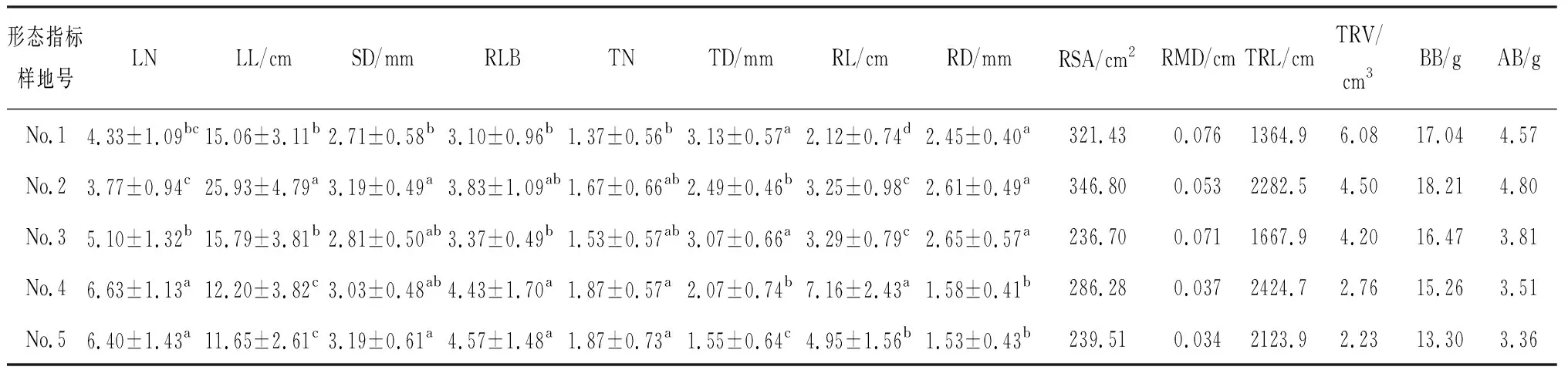
表2 青藏苔草形態特征及生物量Table 2 Morphological characteristics and biomass of Carex moorcroftii
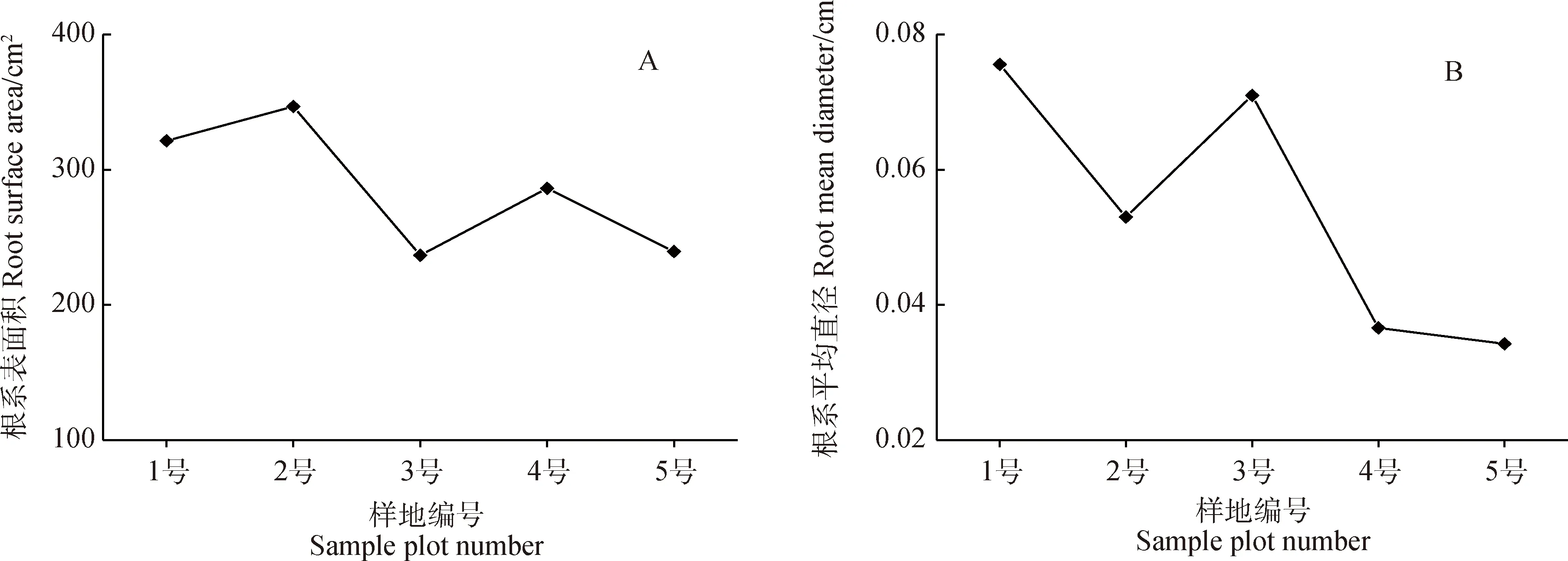
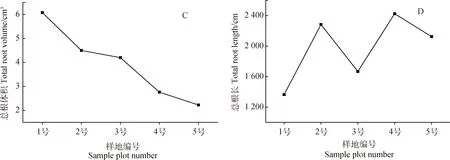
圖2 根系形態特征的變化Fig.2 Variation of root morphological characteristics
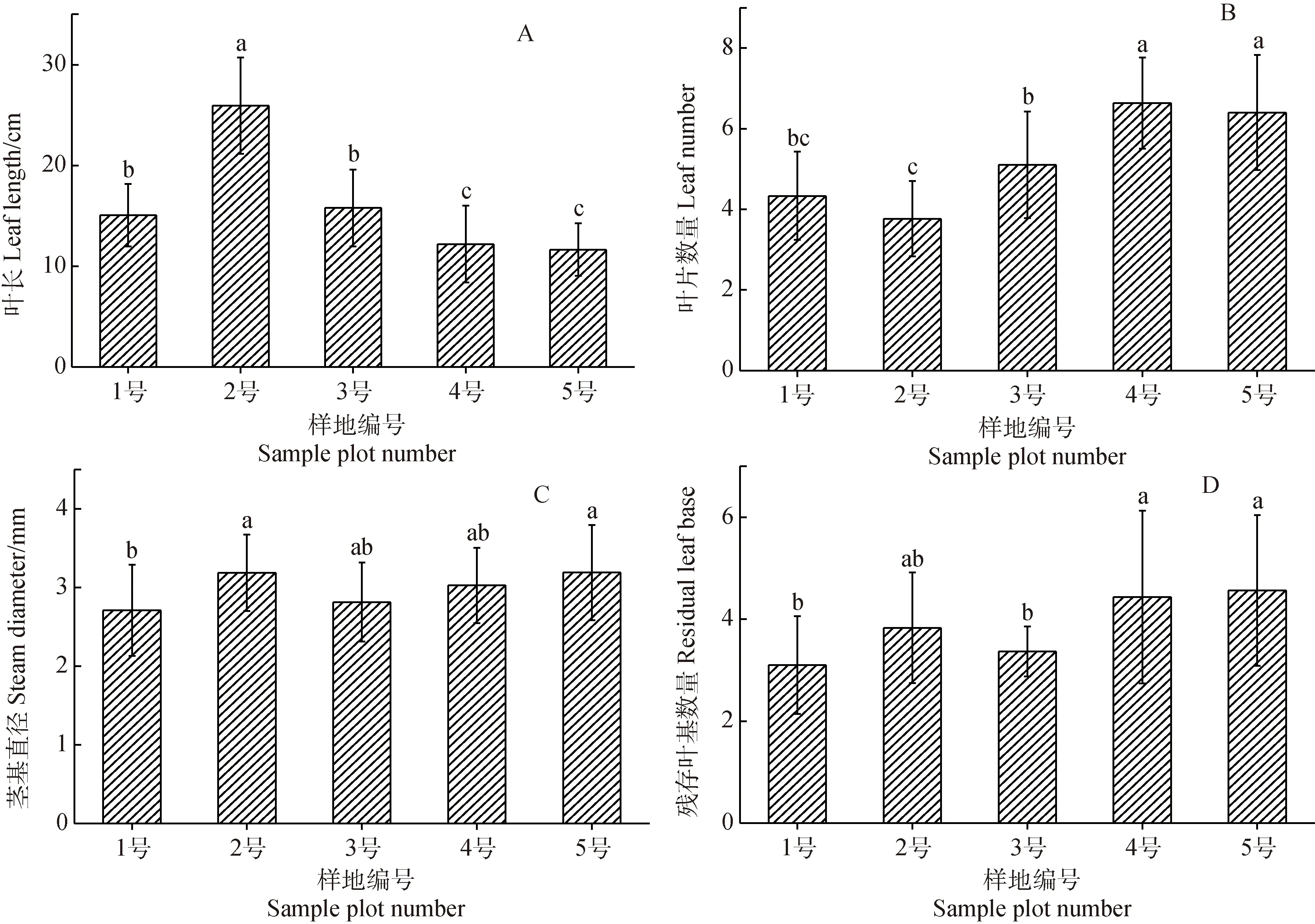
圖3 莖、葉形態特征的變化Fig.3 Variation of stem and leaf morphological characteristics
2.3 水分梯度下生物量的變化與分配
青藏苔草地下、地上生物量均隨土壤含水率的降低而減小,BB/AB值先減小后增大(圖5)。2號樣地地下、地上生物量最大,分別為4.8 g,18.21 g;5號樣地地下、地上生物量最小,分別為3.36 g,13.30 g;4號樣地的BB/AB值最小,為0.23,1號樣地的BB/AB值最大,為0.27。
2.4 營養器官形態可塑性排序
根據RDA排序圖,青藏苔草形態特征可塑性對土壤水分梯度的響應可以分為以下3類:(1)可塑性強形態特征,包括總根體積、根系平均直徑、根狀莖長度;(2)可塑性中等形態特征,包括葉長、分蘗芽直徑、根狀莖直徑、根系表面積、葉片數量、殘存葉基數量、總根長;(3)可塑性弱形態特征,包括地上生物量、地下生物量、莖基直徑、分蘗芽數量。各個形態特征的可塑性強弱不同,因此隨土壤含水率變化的程度也不同,存在非對稱響應機制。綜合營養器官形態特征可塑性可得出,根的可塑性最強,莖的可塑性中等,葉的可塑性最弱。

圖4 克隆構件形態特征的變化Fig.4 Variation of morphological characteristics in clonal modular
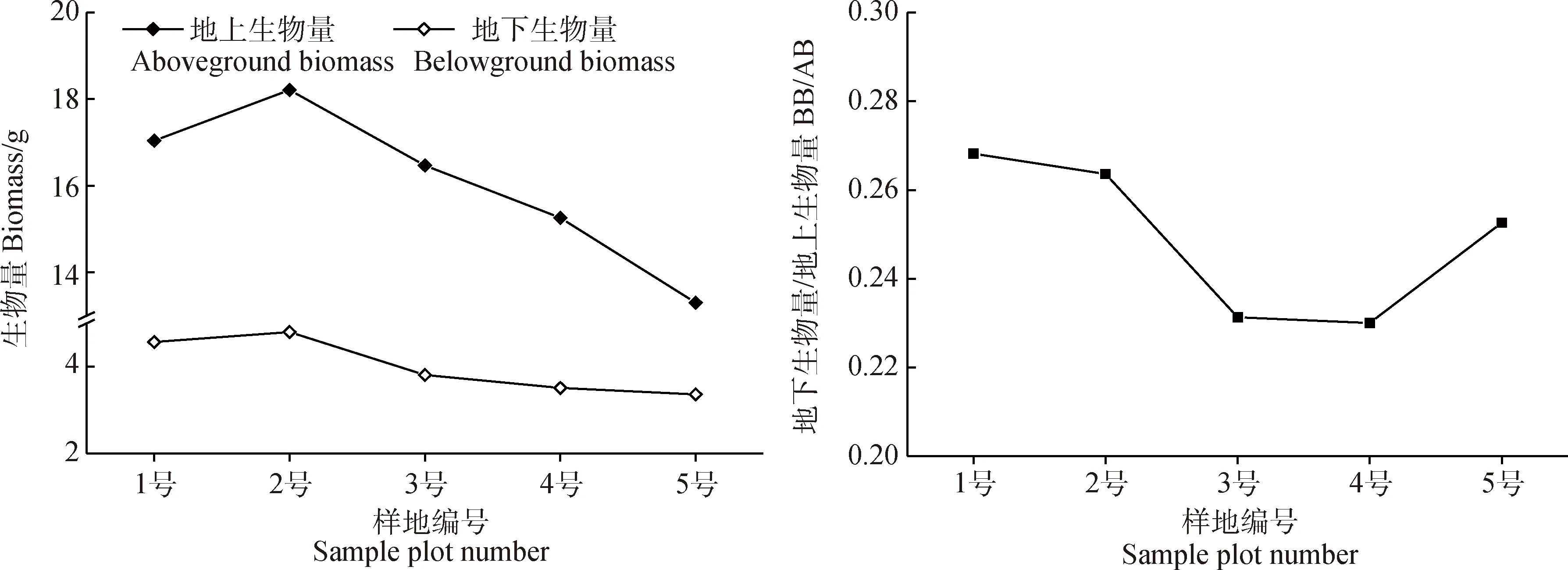
圖5 生物量與BB/AB值的變化Fig.5 Variation of biomass and BB/AB ratio
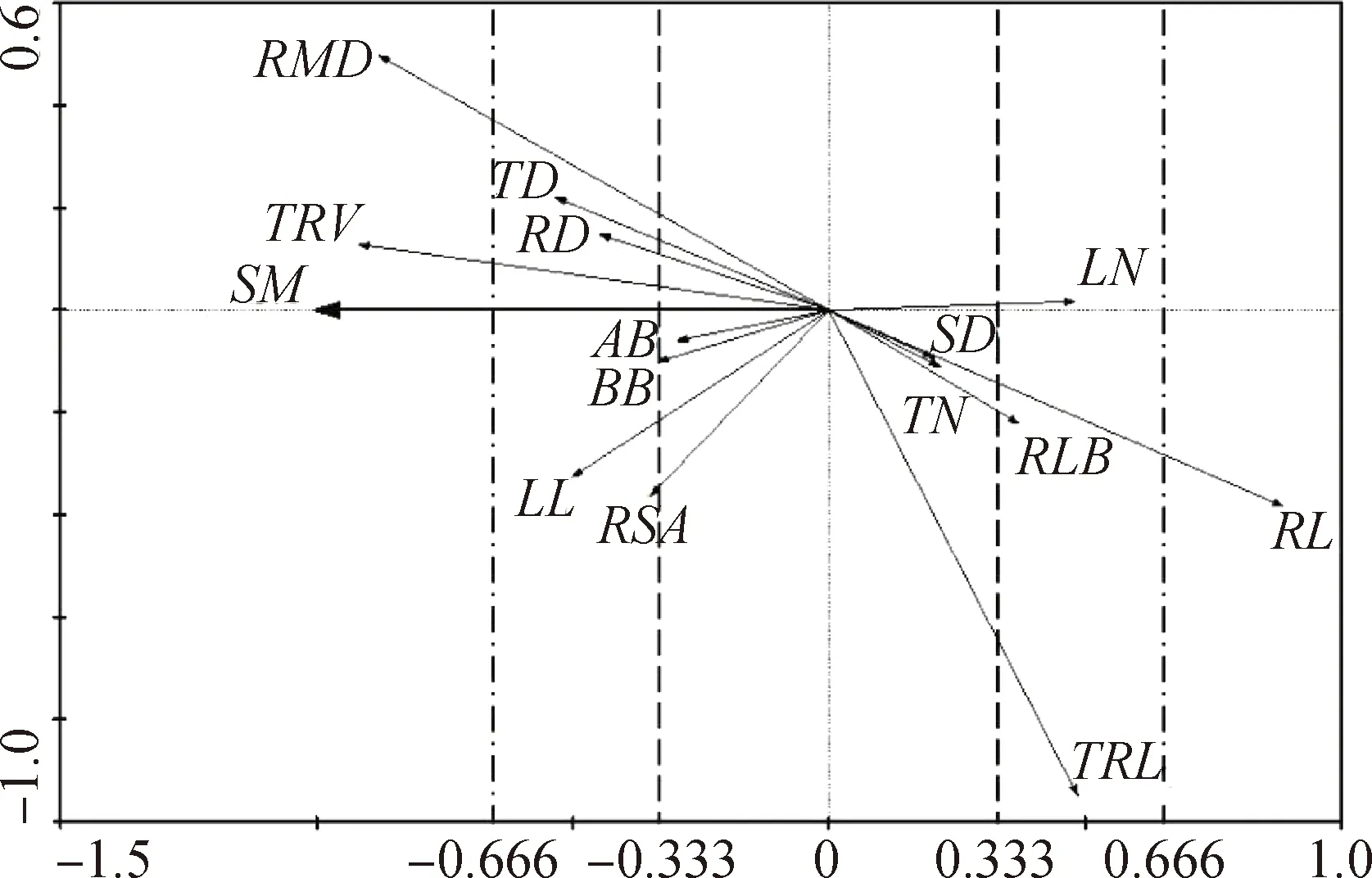
圖6 形態特征與土壤含水率RDA排序圖Fig.6 RDA ordination diagram of morphological characters and soil moisture content注:SM,土壤含水率Note:SM,soil moisture
3 討論
3.1 形態特征的非對稱響應
植物在受到干旱脅迫時,將調整形態特征之間的關系以適應生境變化,這將導致形態特征對水分響應敏感度的分化[31],即在生境梯度下,植物不同性狀在變化的同時,變化的程度卻存在較大差異,這種非對稱響應機制(Asymmetric responses)在生態系統中普遍存在,已有學者進行了相關報導[32]。RDA排序分析將青藏苔草形態特征對水分響應的敏感度分為強、中、弱3類(圖5),表明青藏苔草形態特征沿土壤水分梯度的變化存在非對稱響應機制。隨著生境干旱化,青藏苔草總根體積、根系平均直徑可塑性大,地上生物量、葉片數量可塑性小,這與黃櫨[33](Cotinuscoggygria)的研究結果一致,說明土壤水分的降低大大刺激了植株的根系發育,根在青藏苔草適應干旱生境的過程中起著十分重要的作用。隨著生境干旱化,生物量分配朝著根莖分配、葉片分配減小的方向發展,這與根、莖、葉形態特征的非對稱響應有關,根的直徑、表面積、體積減小的程度大于根長增加的程度,分蘗芽、根狀莖直徑減小的程度大于分蘗芽數量、根狀莖長度增加的程度,故其地下生物量下降;葉長、莖基直徑減小的程度大于葉片數量增加的程度,故其地上生物量下降。青藏苔草對各種性狀進行權衡,根、根狀莖與分蘗芽偏向細長化,葉偏向數量化,以達到在干旱化生境中的最優生活對策。
3.2 營養器官形態特征對水分梯度響應的差異
根是植物吸收水分和養分的主要營養器官,當水分成為限制植物生長的條件時,植物會增加根長,使根系吸收利用深層土壤水分,從而適應干旱生境[34-35]。對青藏苔草的研究發現,生境干旱化使得青藏苔草根系形態特征發生了改變,且各個指標的變化趨勢存在差異。根系平均直徑、根系表面積、總根體積均隨生境干旱化程度的增加呈減小趨勢,而總根長隨生境干旱化程度的增加呈增大趨勢(圖2)。高山嵩草和矮生嵩草在適應干旱化生境時,同樣呈現出根系長度增加的形態變化[16],這說明青藏苔草和高山嵩草、矮生嵩草在干旱條件下的生存策略相似,都會通過調節根直徑、表面積、體積與根長的關系來利用極為有限的資源。在干旱生境中,青藏苔草得不到充足的水資源,光合作用被抑制,僅能獲得有限的光合產物,故其根系直徑減小,將有限的資源用于根系的伸長生長,以利用深層土壤水源。
隨著生境干旱化,青藏苔草呈現出葉長縮短,莖基直徑增大,葉片與殘存葉基數量增多的形態變化特征(圖3)。在生境干旱化條件下,青藏苔草葉長縮短的形態變化與西藏嵩草(K.tibetica)的葉片變化相同[36];莖基直徑的形態變化與其它高原濕地植物受到干旱脅迫后的變化相反[37];葉片數量的變化與灰化苔草(C.cinerascens)、高山嵩草和矮生嵩草在生境干旱化過程中通過減少葉片數量降低蒸騰作用,減少水分散失的變化不同[16,38],說明青藏苔草與其他植物在干旱條件下的生存策略有所不同。葉片是植物形成光合產物的重要器官,葉長縮短雖然極大的降低了青藏苔草與其他植物的競爭力[39],但水分與養分的運輸距離縮短,大大提升了運輸效率[40],并節省了運輸的能量消耗。因此,在葉長縮短的情況下,增加葉片數量對維持植株的光合葉面積具有重要意義,這是青藏苔草在干旱脅迫生境中的最優適應策略。莖基部是多年生草本植物葉片的著生區,有著貯藏營養物質的作用,是葉片數量增加的基礎,莖基直徑越大,貯藏的營養物質越多,能供給葉片生長的營養就越多[16],殘存葉基數量則隨葉片數量的增加而增多。綜上,在生境干旱化的影響下,青藏苔草縮短葉長提升運輸效率,同時增加莖基直徑及葉片數量以維持光合作用,殘存葉基數量也隨葉片數量的增多而增多,4個形態特征之間表現出很強的相關性。
克隆植物的生殖主要靠克隆構件形成新的無性系分株來完成,無性繁殖力強,無性系存活的時間也較長[41-42]。隨著生境干旱化,青藏苔草的根狀莖直徑減小、長度增加,分蘗芽直徑減小、數量增多(圖4)。分蘗芽數量的變化與生境干旱化條件下高山嵩草的響應機制相同[16],說明青藏苔草與高山嵩草在干旱化生境中的無性繁殖能力都增強。隨著干旱化程度增加,青藏苔草的資源積累受到限制,導致其資源進行轉移,具體表現為根狀莖與分蘗芽的直徑減小,而根狀莖長度與分蘗芽數量增加,這種資源的分配轉移有利于青藏苔草占據土壤空間,增加與其他植物的競爭力。已有研究表明,苔草屬植物存在生理整合現象(Physiological Integration),即在由地下根莖相連的各分株之間,營養物質和水分可以通過根莖進行運輸和轉移,實現整個無性系的資源共享,從而提高種群對脅迫環境的適應性[43]。青藏苔草作為典型的克隆植物,在土壤水分虧缺的情況下,既存在一般植物常有的適應生境干旱化的機制,又發揮了克隆植物的優勢,占據更多生存空間,以提高對干旱環境的適應能力。
3.3 生物量分配策略
隨著生境干旱化,青藏苔草的地下、地上生物量都減小,這與西藏嵩草在土壤水分減少條件下地下、地上生物量也減小的研究結果一致,而與矮生嵩草、鵝絨委陵菜(Potentillaanserina)、海乳草(Glauxmaritima)和華扁穗草地下、地上生物量與土壤含水率之間的關系相反[15,36]。矮生嵩草、鵝絨委陵菜、海乳草、和華扁穗草適宜生長于土壤水分適中的平緩灘地或山地陽坡,是耐旱性較強的植物。而西藏嵩草喜歡分布在潮濕或積水的環境中,這與青藏苔草的生境喜好相同,因此,當生境干旱化程度加劇,青藏苔草的地下、地上生物量分配與喜濕植物相同,與矮生嵩草等植物存在差異。青藏苔草BB/AB值隨著生境干旱化,總體上呈現出緩慢降低的趨勢,在最濕潤生境(0.27)中BB/AB值最大,在半干旱半濕潤生境(0.23)中BB/AB值最小,而在最干旱生境(0.25)中出現回升。其原因是隨著生境干旱化,地上生物量減小的程度比地下生物量減小的程度要大的多。即青藏苔草在干旱化生境中更偏向于維持地下生物量,減小地上生物量。
生物量可以直觀地反映研究對象積累有機物的多少,關系著植物的發育及其結構組成,植物會通過改變生物量的分配以適應逆境[44]。當環境發生改變時,植物將改變對不同器官的生物量分配,最大限度地提高獲取各種資源的能力[45],隨著生境干旱化,青藏苔草的地下、地上生物量都減小,說明其在干旱化的生境中獲取維持生命和生長所需資源的能力有限,故隨著土壤含水率降低,它的個體空間分布格局從均勻分布逐漸過渡到隨機分布,個體數量及分布范圍逐漸減小。此外,青藏苔草的地下生物量減小的程度小于地上生物量,說明在干旱化生境中,地下部分對其適應干旱生境的意義更大。
4 結論
為適應干旱化生境青藏苔草會改變自身表型特征以維持自身正常生長和繁殖,根的可塑性比莖和葉強,地下部分對其適應干旱化生境的意義更大。青藏苔草適應干旱化生境的策略為:葉片長度縮短、數量增多在縮短輸導距離的同時保持植株的葉面積;發揮克隆植物的優勢,增加分蘗芽數量與根狀莖長度以增強對空間的占據能力;減小根系直徑,增加根系長度以吸收利用深層土壤水分。根、莖、葉形態特征具有一定的可塑性,但生物量的降低減弱了青藏苔草與其他植物的競爭力,限制了青藏苔草在干旱生境中的分布。研究結果有助于深入認識干旱區濕地植物對土壤水分的響應關系,對于合理利用干旱區濕地資源、保護干旱區濕地生態系統具有重要參考價值。
