茶樹對茶小綠葉蟬為害的響應及其機制研究進展
2021-04-08 04:16:46巨學陽羅智捷薛蓉蓉曾任森宋圓圓
福建農林大學學報(自然科學版) 2021年2期
關鍵詞:植物
王 杰, 童 璐, 巨學陽, 羅智捷, 薛蓉蓉, 高 州, 曾任森, 宋圓圓
(1.福建農林大學農學院,作物遺傳育種與綜合利用教育部重點實驗室;2.福建農林大學閩臺作物有害生物生態防控國家重點實驗室,福建 福州 350002)
茶樹(Camelliasinensis),隸屬山茶科山茶屬,廣泛分布在熱帶和亞熱帶溫暖濕潤的地區.目前,茶是世界上最受歡迎的三大飲品之一[1].茶葉的市場價格主要由它的口感、香氣決定,這主要與茶樹的代謝產物密切相關[2].茶樹的葉片和葉梢是茶葉加工的原料,然而,茶樹的葉片和嫩梢常會被許多害蟲取食為害,進而影響茶葉品質.
茶小綠葉蟬(Empoascaonukii)是茶樹上的重要害蟲,廣泛分布于我國各主要茶區[3].茶小綠葉蟬世代交替重疊、個體小、繁殖能力強、為害歷期長,防治難度相對較大[4].雌蟲將蟲卵產于茶樹嫩梢韌皮部,阻礙茶樹體內營養物質的運輸,若蟲和成蟲均能從茶樹葉片吸食汁液,破壞茶樹葉肉組織.茶樹受害嚴重時葉緣、葉尖呈紅褐色焦枯狀,茶葉產量和質量受到影響[5-6].茶小綠葉蟬的防治主要依賴化學農藥,但化學農藥的不合理施用使得茶小綠葉蟬對吡蟲啉、聯苯菊酯和啶蟲脒等農藥產生了抗性[7],同時導致茶葉化學農藥殘留超標[8-9].值得注意的是,茶樹適度地被取食可以讓成品茶釋放特殊風味[10],如著名的“東方美人茶”就是用茶小綠葉蟬取食過的茶梢加工而成[11].因此,茶樹與茶小綠葉蟬之間的互作是非常獨特而復雜的[12].茶樹的代謝物質在其抵御病蟲害方面具有十分重要的作用,強化該方面的研究,可以為減少化學農藥的使用提供依據,進而實現茶葉的綠色安全生產[13].本文重點圍繞調控茶樹防御反應的茶小綠葉蟬分泌物、茶樹對茶小綠葉蟬為害的分子應答機制以及茶樹代謝物在茶小綠葉蟬與茶樹互作中的作用等方面的研究成果進行分析,以期為深入了解茶樹與茶小綠葉蟬間復雜的互作關系提供參考,并為研發茶樹害蟲防治新技術和提高茶葉品質提供依據.
1 調控茶樹防御反應的茶小綠葉蟬分泌物
植物與昆蟲在協同進化的過程中需要準確識別一般的機械損傷和昆蟲的取食為害,進而在必要時通過提高相關代謝物質的含量來防御害蟲的為害[14].植物對機械損傷和昆蟲取食為害所做出的防御反應存在很大差異,主要是因為昆蟲除了對植物造成一定程度的機械損傷外,還在取食或產卵過程中分泌某些化學物質,這些與植食性昆蟲相關且可被植物感知的信號化合物被稱為昆蟲相關分子模式(herbivore associated molecular patterns, HAMPs)[15-16].HAMPs主要是蛋白質、多肽、酰胺、脂肪酸等,植物能夠識別這些物質,從而激活有別于一般的機械損傷介導的防御反應[16].已有研究者利用同位素示蹤法,證明刺吸式昆蟲如葉蟬、飛虱等主要通過口針吸食植物細胞汁液,并在取食時將一些可溶性的成分(大部分是酶類物質)分泌到植物組織中[17-18].茶小綠葉蟬體內的一些酶類物質(如香葉醇合成酶、海藻糖酶和蛋白水解酶等)可能是其參與調控茶樹與茶小綠葉蟬之間互作的重要HAMPs.
1.1 香葉醇合成酶
香葉醇是牻牛兒基焦磷酸在香葉醇合成酶作用下合成的,它對茶葉的香氣有重要影響[2],如圖1所示.研究發現,茶樹葉片被茶小綠葉蟬為害后會釋放出香葉醇,但其對茶樹體內香葉醇合成酶活性沒有影響[19].這是由于香葉醇合成酶是來源于茶小綠葉蟬,其在蟲體內活性相對較高,推測它可能來源于昆蟲脂肪體或消化系統,在取食過程中分泌于取食部位,進而與葉片中香葉醇前體牻牛兒基焦磷酸反應后釋放出香葉醇[20].香葉醇是植物中的一種重要的防御性次生代謝物,它對昆蟲有趨避作用[21],以此抵抗茶小綠葉蟬的為害.
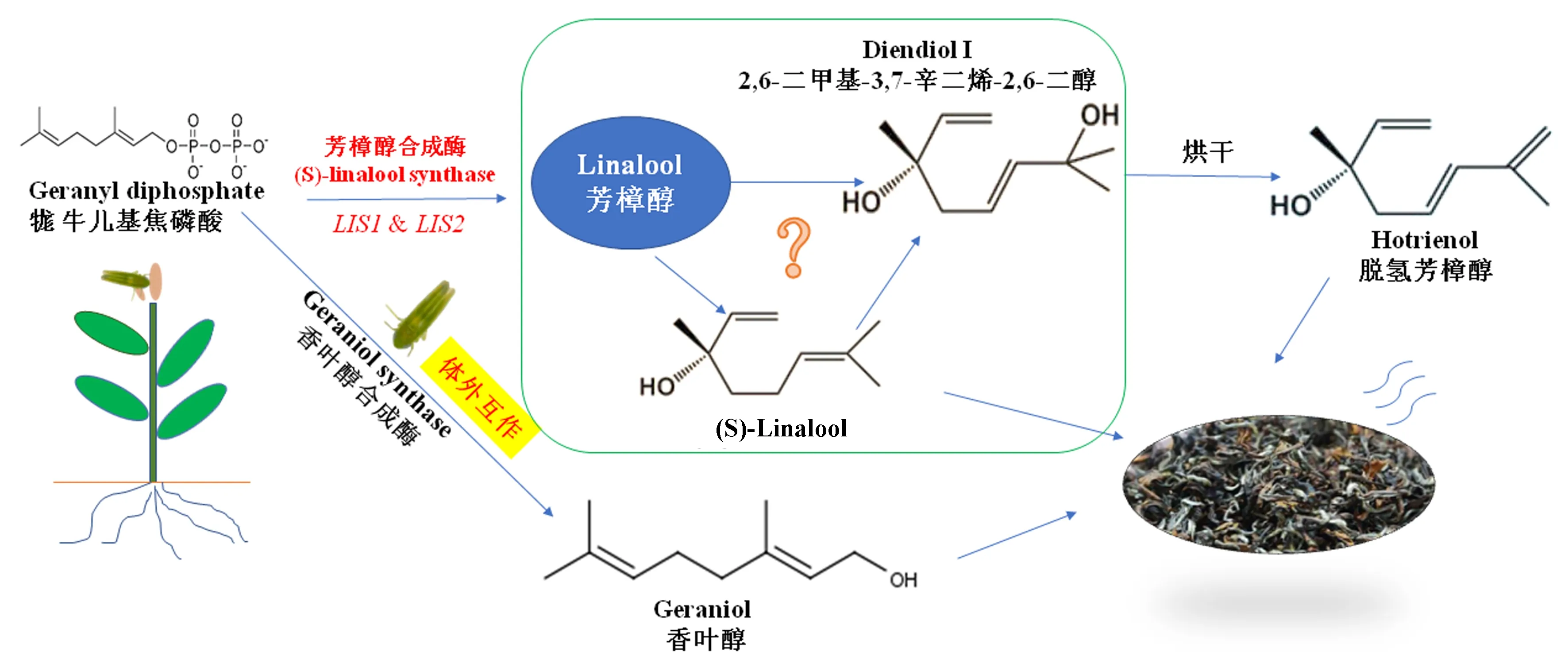
圖1 茶小綠葉蟬取食誘導茶樹生產芳樟醇和香葉醇Fig.1 The production of linalool and geraniol in tea plant after attacked by E.onukii
1.2 海藻糖酶
汪曉茜[22]成功從茶小綠葉蟬中克隆到海藻糖酶基因,并發現其對茶小綠葉蟬抵抗農藥脅迫有重要作用.海藻糖酶,又稱海藻水解酶,昆蟲利用它分解海藻糖從而為自身代謝提供重要的能量,它也是昆蟲合成幾丁質途徑中的重要酶,可以影響昆蟲體內幾丁質合成酶的表達,進而影響幾丁質的合成[23].植物在遭受昆蟲取食為害后,體內包括幾丁質酶在內的防御物質大量增多[24].海藻糖酶已經被證實參與調節植物的防御反應[25].然而,茶小綠葉蟬在為害茶樹過程中是否分泌海藻糖酶,并影響茶樹的化學防御反應或代謝過程需要進一步驗證.
1.3 蛋白水解酶
基于轉錄組數據和cDNA末端快速擴增(rapid amplification of cDNA ends, RACE)技術,于永晨等[26]成功從茶小綠葉蟬中克隆到半胱氨酸蛋白酶基因.在植食性昆蟲體內,半胱氨酸蛋白酶是重要的水解蛋白酶之一,它主要參與食物消化、生長發育、免疫調節等重要生命活動[27].胰蛋白酶也是昆蟲體內一種重要的蛋白水解酶,一般在刺吸式口器昆蟲的唾液腺中含量豐富,其可以幫助昆蟲消化食物[28-29].羅智捷[30]利用RACE等技術從茶小綠葉蟬中成功克隆到一個類胰蛋白酶基因,通過體外表達成功獲得類胰蛋白,并證實該胰蛋白酶參與誘導茶樹的防御反應.昆蟲口腔分泌物中還有一種蛋白水解酶β-葡萄糖苷酶可直接影響植物揮發物的釋放,它與存儲在植物體內糖苷化的化合物反應變成揮發性物質釋放出來,它還可以誘導植物產生防御反應[31-32].本課題組在茶小綠葉蟬唾液腺中也發現了β-葡萄糖苷酶.茶尺蠖(Ectropisoblique)在取食茶樹過程中會分泌β-葡萄糖苷酶,作用于取食部位,使茶樹釋放出與健康新梢完全不同的揮發物,增強對天敵的吸引力,從而減輕茶樹的受害程度[33].因此推測茶小綠葉蟬分泌的β-葡萄糖苷酶也有類似功能.
茶樹啟動防御反應除了與茶小綠葉蟬取食或產卵造成的機械損傷程度有關之外,還可能受茶小綠葉蟬釋放的化學物質的影響.然而,有關這方面的研究仍非常缺乏.如圖1所示,茶小綠葉蟬取食為害誘導茶樹體內兩個芳樟醇合成酶基因(LIS1和LIS2)上調表達,從而刺激芳樟醇從茶樹體內釋放出來[19].目前,2,6-二甲基-3,7-辛二烯-2,6-二醇(diendiol I)在茶樹中產生的機制仍不清楚.有學者認為它是來源于芳樟醇[34-35];持續的機械損傷也能使得茶樹體內的LIS1和LIS2上調表達,但機械損傷并不能使茶樹產生diendiol I[35],因此推測茶小綠葉蟬可能分泌某種氧化酶參與由芳樟醇形成diendiol I的氧化過程.
2 茶樹對茶小綠葉蟬為害的分子應答機制
2.1 相關信號響應
植物通過細胞表面模式識別受體(pattern recognition receptor, PRRs)感受到昆蟲的取食為害,并在識別后激活激素信號網絡來調控抗蟲防御反應[15,36-37].一般而言,咀嚼式口器的昆蟲對植物組織損傷較大,主要觸發茉莉酸(jasmonic acid, JA)信號通路,而刺吸式口器昆蟲對植物組織的損傷相對較小,主要激活水楊酸(salicylic acid, SA)信號通路[35,38].除了JA和SA之外,脫落酸(abscisic acid, ABA)、乙烯(ethylene, ET)、油菜素甾醇(brassinosteroids, BR)、細胞分裂素(cytokinin, CTK)等也參與調控植物對蟲害的防御反應[39-40].其中,JA與SA之間存在拮抗關系,這是植物為了適應眾多不同脅迫的一種調控策略[41];同時,JA與SA也存在協同作用,如當植物被蜘蛛螨(Tetranychusurticae)為害后,JA和SA信號通路協同參與到植物對蜘蛛螨的防御反應中[42].相比于吸食莖稈韌皮部汁液的刺吸式昆蟲而言,茶小綠葉蟬取食對茶樹葉片的損傷更大,這可能造成茶樹防御茶小綠葉蟬為害的反應與植物對一般刺吸式昆蟲的防御反應存在差別.
茶小綠葉蟬為害的茶樹(金萱)葉片中JA、SA和ABA含量均有所上升,然而持續通過機械損傷模擬茶小綠葉蟬取食只誘導JA和ABA含量上升,對SA沒有顯著影響[43].最新的研究同樣證實茶樹(金萱)JA和SA激素信號會受到茶小綠葉蟬取食的誘導,其中SA信號似乎在茶樹防御茶小綠葉蟬為害的過程中起主導作用[44].茶樹經水楊酸甲酯(methyl salicylic acid, MeSA)或茶小綠葉蟬處理后,兩者的茶梢揮發物組成相似,其中,乙酸乙酯、乙醇、反-2-己烯醛、羅勒烯、反式-芳樟醇氧化物、順式-芳樟醇氧化物、芳樟醇、法呢烯、MeSA、丁酸順-3-己烯酯、香葉醇、苯甲醇、苯甲醛和 3,7,11-三甲基-1,6,10-十二烷三烯-3-醇等組分含量上升,順-3-己烯-1-醇等組分含量明顯下降[45].茶園中的茶樹經MeSA處理后對茶小綠葉蟬產生了比較明顯的趨避作用,且茶梢的防御相關酶活性和基因表達情況與茶小綠葉蟬為害誘導的結果相似[46].
有研究認為,茶小綠葉蟬刺激茶樹產生抗性主要與JA信號通路有關.根據轉錄組測序結果,茶樹(鳥王種)被茶小綠葉蟬為害后的差異表達基因主要與植物激素信號轉導和亞麻酸代謝途徑有關[47],而JA主要通過亞麻酸的十八烷代謝途徑進行下游信號的傳導來調控植物的防御反應[48].不同茶樹品種的抗性水平存在差異[49-50],可能造成茶樹啟動抗蟲相關激素信號的不同.例如:經茶尺蠖取食后,JA在茶樹抗性品種(上梅州)中更容易被誘導;而SA水平在敏感品種(平陽特早)中始終維持在較高的水平,JA含量并未因害蟲取食而發生顯著改變[51].因此,有必要深入探索茶樹抗性品種和敏感品種應對茶小綠葉蟬為害時體內SA和JA的作用機制,為培育茶樹抗蟲品種提供更多的依據.
2.2 相關基因表達
隨著基因測序技術的不斷發展,茶樹的基因組測序工作已初步完成,為茶樹功能基因的挖掘以及生長發育和品質的調控及抗性機理等方面的深入研究奠定了基礎[52-53].
消減抑制雜交(suppression subtractive hybridization, SSH)是一種以mRNA 差別顯示為基礎建立起來的篩選差異表達基因的技術[45].利用SSH對茶樹(鳧早2號)被茶小綠葉蟬取食為害后的差異表達基因進行篩選分析,在所分離的21個差異表達基因中包括了病程相關蛋白(pathogenesis-related proteins, PRs)基因、核糖體非活性蛋白(ribosome-inactivating protein, RIP)基因、鈣離子感受蛋白-絲氨酸蛋白激酶(calcium sensor protein-serine protein kinases, CBL-CIPK)信號相關基因、細胞壁強化基因、次生代謝產物合成基因、水轉運基因以及活性氧相關基因[54].此外,通過建立SSH數據庫發現茶小綠葉蟬取食可以特異性激發大吉嶺茶磷酸海藻糖合成酶基因和乙醇脫氫酶的表達[55].
利用轉錄組測序(RNA-seq)研究都勻毛尖茶(鳥王種)對茶小綠葉蟬取食為害的響應,結果發現與對照相比,茶小綠葉蟬為害12 h有555個差異表達基因,其中上調表達基因有399個,下調表達基因有186個.通過KEGG功能顯著性富集分析得出,茶小綠葉蟬取食為害所引起差異表達的基因主要參與植物激素信號轉導、亞麻酸代謝、苯丙素的生物合成以及萜類化合物合成,其中參與植物激素信號轉導的差異基因最多,推測其在茶樹抗蟲防御過程中起著非常重要的作用[47].
轉錄組數據顯示,與正常的處理相比,茶小綠葉蟬為害顯著誘導了茶樹體內調控JA和SA通路的相關轉錄因子上調.具體來說,茶樹(金萱)被茶小綠葉蟬取食誘導上調的轉錄因子家族包括WRKY(24)、AR2/ERF(24)、bHLH(18)、NAC(10)和GRAS(10).其中,WRKY是SA信號通路的主要調節因子,AR2/ERF是JA信號通路ERF分支的調節因子;JA信號通路主要調控因子bHLH家族有18個基因顯著上調,其中包括4個編碼MYC2的基因;此外,NAC家族中的NAC90和NAC2,以及GRAS家族中的SCL21和SCL33,均可以作為調控茶樹對茶小綠葉蟬抗性的潛在因子[44].除了在植物激素信號通路上發現較多上調的基因外,茶小綠葉蟬取食誘導的抗性基因在苯丙氨酸類黃酮生物合成和蠟質合成通路上也有顯著富集[44].
3 茶樹代謝物在茶小綠葉蟬與茶樹互作中的作用
植物被昆蟲取食為害后,通過激活植物激素介導的信號轉導途徑,調控與其相關的代謝通路基因表達,最終引起植物代謝組分發生變化[16].植物代謝物主要包含初生代謝物質和次生代謝物質.其中,初生代謝物質主要包括丙氨酸、天冬氨酸、谷氨酸等各種游離氨基酸,葡萄糖、果糖等糖類,有機酸(蘋果酸、甘油酸)等;次生代謝物質主要包括酚類、萜類、含氮化合物等,植物受到昆蟲取食或機械損傷后這些物質會產生顯著的變化[56].茶樹被茶小綠葉蟬為害后,其代謝物質也會受到嚴重影響.茶樹代謝物分為非揮發性和揮發性兩種,這些物質除了在茶樹抵御蟲害過程中起重要作用外,還在一定程度上影響茶的品質.
3.1 非揮發性代謝物
3.1.1 游離氨基酸 茶樹體內游離的氨基酸對茶葉風味的影響很大,茶葉中最主要的游離氨基酸是谷氨酸和茶氨酸,大約占茶葉干重的1%~2%[57-59].通過對茶樹(金萱)葉片中20種氨基酸進行定量分析,發現茶小綠葉蟬取食3周后茶樹葉片中谷氨酸和茶氨酸等主要的游離氨基酸含量與受到機械損傷處理的茶樹相比呈現下降趨勢[11].在短時間(10 d)處理下,茶樹葉片中游離氨基酸含量隨著茶小綠葉蟬取食為害程度的加強而呈現降低的趨勢[60].氨基酸是昆蟲生長的必需營養物質,茶小綠葉蟬取食后的茶樹葉片中游離氨基酸(如茶氨酸和谷氨酸)含量下降可能是茶樹對茶小綠葉蟬的一種防御策略.
3.1.2 多酚類物質 外界脅迫往往會引起植物多酚物質的積累[61].茶葉中茶多酚的最主要成分是黃酮類化合物兒茶素,約占茶葉干重的30%[62].黃酮等多酚類物質在很多植物中被發現與植物抗病性和抗蟲性密切相關[63].隨著茶小綠葉蟬取食為害時間的延長,茶樹(金萱、福云六號)葉片中兒茶素、黃酮含量呈現先快速增多后減少的趨勢[60].茶樹(金觀音)損傷程度隨著茶小綠葉蟬蟲口數量和為害時間的增加而增大,且茶樹損傷程度(0、5、10和20頭)與其受到蟲害后產生的多酚含量呈現非線性的關系,在中度損傷(10頭)時多酚的含量最高[64].研究發現,植食性害蟲取食能激活植物體內JA介導的信號轉導網絡,通過誘導苯丙氨酸酶(phenylalanine ammonia lyase, PAL)和多酚氧化酶(polyphenol oxidase, PPO)活性,使得植物體內多酚含量增加,從而增強對害蟲的抵抗能力[65-66].此外,兒茶素等多酚物質是形成茶紅素和其他聚合物的前體化學物質,它會影響紅茶和烏龍茶的湯色和口感[67-68].這些研究說明多酚物質在茶樹響應茶小綠葉蟬脅迫和成品茶品質方面起著重要作用.
3.1.3 含氮次生代謝物 茶樹中含氮次生代謝物主要包括生物堿、非蛋白氨基酸、防御蛋白等.茶樹中的生物堿主要有咖啡因、可可堿以及茶堿等,這些物質與揮發物以及多酚化合物的不同之處在于它們幾乎很少受到制茶工藝的影響,其含量在成茶和鮮葉中相差不大,它們的含量主要與茶樹品種和外界脅迫有關[69].
鄒武等[70]對福建4個茶樹品種(福云、鐵觀音、毛蟹、黃棪)與茶小綠葉蟬種群的關系進行調查分析,發現在生化指標方面,茶小綠葉蟬的種群數量與茶樹葉片中咖啡堿含量呈一定負相關性.金珊等[71]研究表明,大部分茶樹抗蟲品種中非蛋白氨基酸類物質γ-氨基丁酸含量高于感蟲品種,且隨著人工飼料中γ-氨基丁酸濃度的升高茶小綠葉蟬的死亡率也增大.
潘杰[72]成功從茶樹中克隆了半胱氨酸蛋白酶抑制劑基因和核糖體失活蛋白基因,通過體外表達成功獲得這兩種蛋白,且發現核糖體失活蛋白處理能降低茶小綠葉蟬的存活率.植物體內的可溶性蛋白含量變化可以反映植物受脅迫程度.調查發現,茶小綠葉蟬蟲口密度與茶樹中可溶性蛋白的含量呈現負相關性,可溶性蛋白含量高的新梢顏色較深,不易招引昆蟲,且可溶性蛋白含量高的葉片皮層組織厚,不易取食[73-74].利用2D-PAGE和LC/MS/MS分別對遭受不同程度茶小綠葉蟬為害的烏龍茶鮮葉進行蛋白組學分析,發現其中有89個差異蛋白,這些蛋白主要參與生物合成、碳水化合物代謝與轉運、脅迫響應以及氨基酸代謝過程,所以這些蛋白質會影響茶樹抗性和代謝活動[75].
3.2 蟲害誘導的揮發性物質(herbivore-induced plant volatiles, HIPVs)
3.2.1 綠葉性揮發物 綠葉性揮發物(green leaf volatiles, GLVs)是由植物體內的亞油酸以及α-亞麻酸經脂氧合酶與脂肪酸過氧化氫裂解酶等的一系列酶促反應而形成.一旦植物細胞受損傷該類物質就會立即釋放,主要是一系列小分子質量的脂肪酸衍生物,包括6個碳的醛、醇和脂類,這些物質也被稱為C6揮發物[76].茶樹被茶小綠葉蟬為害后產生的GLVs主要有順-3-己烯醛、順-3-乙酸葉醇酯、順-3-己烯醇、反-3-己烯醛、苯乙醇等[6],其中,順-3-己烯醇對茶小綠葉蟬有顯著的引誘效果[77],并且外源施加該物質可以誘導茶樹體內JA的含量增多,增強茶樹對害蟲的抗性[78].
3.2.2 萜類揮發物 與被咀嚼式口器害蟲為害相比,茶樹被茶小綠葉蟬為害后所釋放的GLVs的量明顯減少,而萜烯類物質和水楊酸甲酯的含量相對增多[79].揮發性單萜或倍半萜類物質如羅勒烯、法尼烯和月桂烯等是由萜烯類合成酶等經過一系列酶促反應而產生[79-80].茶樹(龍井43)被較多茶小綠葉蟬(200頭·株-1)取食28 h后大約釋放31種揮發物,其中25種也出現在被少量茶小綠葉蟬(100頭·株-1)取食的茶樹中;羅勒烯、法尼烯和(E)-4.8-二甲基-1,3,7-壬三烯占據兩種蟲害處理產生的混合揮發物總量的90%,并且蟲害嚴重時這3種物質釋放的量更大[81].不同茶樹抗性品種(高抗鐵觀音和低抗肉桂)被茶小綠葉蟬取食后產生的揮發物,主要有β-月桂烯、β-羅勒烯、反式-β-羅勒烯、α-法尼烯、芳樟醇、(Z)-丁酸-3-己烯酯、癸醛、吲哚、己酸-3-己烯酯和苯乙醇,其中,β-月桂烯、β-羅勒烯、反式-β-羅勒烯、α-法尼烯、芳樟醇屬于組成型揮發物,且在高抗品種鐵觀音上的含量遠遠大于低抗品種肉桂[82].由此可見,茶樹受到茶小綠葉蟬取食為害后所釋放的萜類物質與茶小綠葉蟬為害程度和茶樹品種密切相關.
3.2.3 特殊功能的HIPVs 茶小綠葉蟬取食為害可誘導茶樹產生特異性的揮發物,其中萜類物質在茶樹-茶小綠葉蟬-天敵三級營養互作中起重要作用(圖2).茶樹被茶小綠葉蟬為害后所形成的diendiol I,對捕食性天敵白斑獵蛛(Evarchaalbaria)有明顯的引誘活性[83].diendiol I是用來判斷茶樹是否受到茶小綠葉蟬取食為害的一種標志性物質,因為茶樹被機械損傷和其他昆蟲為害后并不能產生該物質[35].此外,許多研究發現,茶小綠葉蟬為害均誘導茶樹產生β-羅勒烯和α-法尼烯[84-88].已有研究證明,β-羅勒烯和α-法尼烯可以作為茶樹與茶樹間的通訊物質,進而影響茶樹的生理代謝活動和抗蟲防御反應[89-90].茶樹(龍井43)被茶小綠葉蟬取食為害后茶梢揮發性化合物組成和相對含量發生了改變,所釋放出的反-2-己烯醛、苯甲醛、α-法尼烯和水楊酸甲酯可以引誘茶小綠葉蟬卵寄生蜂微小裂骨纓小蜂(Schizophragmaparvula)[91].受到茶小綠葉蟬為害的茶樹所產生的萜類物質橙花醇可以作為啟動植物防御反應的信號物質,促進茶樹體內 JA和ABA含量增多,誘導茶樹積累次生代謝產物,從而提高其對茶小綠葉蟬的抗性[92].HIPVs可以誘導茶樹及其鄰近的茶樹產生防御和代謝方面的變化,進而影響最終成品茶的品質,但其相關的作用機制有待深入探究.
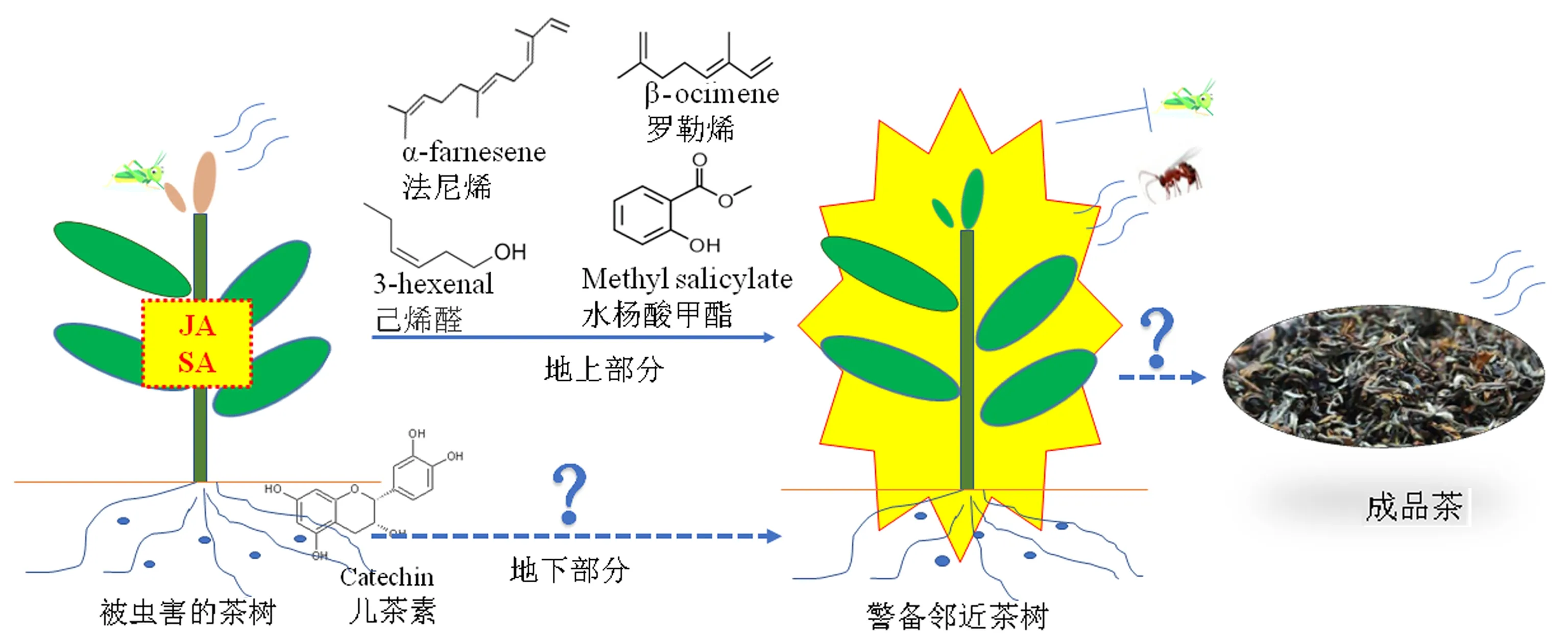
圖2 被茶小綠葉蟬為害的茶樹代謝反應對鄰近健康茶樹的影響Fig.2 Metabolic changes in E.onukii-attacked tea plant and its impact on neighboring healthy tea plant
4 展望
本文從多個方面總結了茶樹對茶小綠葉蟬為害的響應及其機制.茶小綠葉蟬為害茶樹葉片除了造成機械損傷外,還會分泌相關的化學信號物質如HAMPs.茶樹可以識別這些化學信號,從而調控茶樹體內防御信號的應答,誘導產生各類代謝產物.茶樹所產生的代謝物可以影響茶小綠葉蟬的為害行為,其中揮發性物質會影響茶小綠葉蟬對寄主的初步選擇,而非揮發性物質會對茶小綠葉蟬的生長和繁殖造成影響.此外,茶樹中許多次生代謝產物是由于茶小綠葉蟬為害所誘導產生的特異性物質.因此,鑒定茶小綠葉蟬或茶樹相關的化學物質是研究茶小綠葉蟬與茶樹互作關系的關鍵,有利于實際應用.
有關茶樹對茶小綠葉蟬防御信號通路的研究,特別是茶小綠葉蟬為害激活茶樹的激素信號通路及相關基因的表達[43-44],雖然有了一定的進展,但依舊有許多問題亟待解決.茶樹受到茶小綠葉蟬為害后的防御反應是一系列復雜因子綜合作用產生的結果,可能還與效應子、昆蟲共生菌有關.目前還沒有關于茶小綠葉蟬HAMPs或效應子的深入分析,這在一定程度上限制了對茶小綠葉蟬與茶樹的化學互作機制的理解.此外,昆蟲不是獨立的個體,其體內還棲息著大量的微生物,已有大量研究證明昆蟲共生菌在調控植物免疫、降解植物次生代謝物、調節昆蟲免疫等方面有重要作用[81,93-94].毛迎新等[95]對湖北地區5個不同地理位置的茶小綠葉蟬種群的成蟲共生菌進行擴增子測序分析,發現它們共有的優勢菌為鹽單胞菌屬(Halomonas)、希瓦氏菌屬(Shewanella)和沃爾巴克氏體屬(Wolbachia).茶小綠葉蟬優勢菌Wolbachia是在節肢動物體內廣泛分布的一種共生菌,它不僅可以提高宿主昆蟲的生理活動,還能調節昆蟲與植物的相互作用,如Wolbachia侵染玉米根葉甲(Diabroticavirgifera)后可以抑制玉米根部防御相關基因的表達[96].Zhou et al[20]從茶小綠葉蟬上分離到了假單胞菌屬(Pseudomonas)和不動桿菌屬(Acinetobacter)細菌,這兩種菌在其他昆蟲中可以降解植物次生代謝物[97-98].因此,需要進一步加強對茶樹與茶小綠葉蟬互作中茶小綠葉蟬共生菌的研究.
目前,大部分研究主要關注茶樹被茶小綠葉蟬為害后地上部分所產生的代謝物質的變化及其在生態層面的功能[19,35,90],而忽視了對茶樹地下部分的研究.植物地上部分受到蟲害后,會通過植物體內的激素信號網絡,系統性激活植物各部位包括根系的防御反應.例如:入侵植物矢車菊(Centaureamaculosa)受到害蟲取食后根系會釋放大量兒茶素,從而影響本地鄰近植物的生長[91];外源施加JA可以影響擬南芥(Arabidopsisthaliana)的根系分泌物和微生物群落[99];當茶樹地上部分葉片被茶尺蠖取食為害后,地上部分葉片和地下部分根系組織均在轉錄水平上發生響應,且JA會由地上部向根系部位進行長距離轉運,而碳水化合物則由根部向葉片部位進行轉運[100].因此,茶小綠葉蟬為害對茶樹地下部分的影響有待進一步研究.
茶小綠葉蟬對茶樹的影響關系到后期成品茶的品質,因鮮葉所產生的揮發物和揮發物的前體對茶的品質有著重要影響[10,35,55,79].在制茶的過程中鮮葉中的代謝產物或酶會發生進一步的化學反應產生新的代謝產物,因此需要將蟲害和加工過程結合起來研究茶品質.雖然茶小綠葉蟬取食可能會提高茶葉香氣、品質,但是也可能會對茶葉產量和外觀產生負面影響;同時,不同程度蟲害會造成茶葉香氣質量參差不齊,有時候保證了茶的品質而影響了產量[101].因此,如何平衡茶葉品質與產量值得思考.這需要更加深入理解茶小綠葉蟬與茶樹復雜的互作機制,減少生產中對茶小綠葉蟬的依賴,可以考慮利用茶小綠葉蟬相關化學成分進行模擬處理,以此增強茶樹的抗蟲性,減少蟲害,提高茶的產量,同時有利于改善茶的品質.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
