長江靖江段沿岸日本鰻鱺豐度的時間格局及生物學研究
2021-04-01 07:56:08郭弘藝唐文喬倪健夫沈林宏
水生生物學報 2021年2期
郭弘藝 劉 麗 唐文喬 劉 東 張 亞 倪健夫 沈林宏
(1.上海海洋大學海洋動物系統分類與進化上海高校重點實驗室, 上海 201306; 2.江蘇省靖江市漁政管理站, 靖江 214500)
日本鰻鱺(Anguilla japonica)是一種降海產卵洄游型(Catadromous)魚類, 其產卵場位于西馬里亞納海脊附近[1—4]。孵化后的柳葉鰻(Leptocephali)隨北赤道流(North Equatorial Current)和黑潮暖流(Kuroshio Current)輸送至中國、朝鮮和日本的大陸架, 變態為玻璃鰻(Glass eel), 并在河口水域變態為線鰻(Elver)[5]。線鰻溯河至河川或滯留在河口成長為黃鰻(Yellow eel)。數年后黃鰻開始性成熟, 逐步銀化為銀鰻(Silver eel), 返回至產卵場繁殖, 最終完成生活史[6]。這種復雜的生活史特性, 增加了人工繁殖的難度, 目前養殖所用鰻苗還全部依賴于天然捕撈[7]。由于價格高, 鰻苗捕撈一直處于高強度的“掠奪”模式[8], 加上近年來沿海濕地的減少及環境污染, 已造成資源的嚴重匱乏[9]。
江蘇靖江位于長江近口段, 常年水流平順, 受潮汐和長江徑流的雙重影響, 自然形成了流態復雜的水域環境, 不僅是多種魚類棲息和繁殖的良好場所, 亦是長江鰻苗溯河和成鰻降海洄游的通道[10]。近年來, 我國學者對日本鰻鱺的遺傳結構[11,12]、早期生活史特征[13—17]及其資源變動[9,10,18,19]等作了較多研究, 但由于非洄游時期鰻鱺樣本的采集十分困難, 因此其在長江水系的資源狀況一直未見報道。本文基于2002—2017年在長江靖江段長達十余年的采樣調查數據, 對長江近口段日本鰻鱺豐度的時間格局及其年齡結構、全長與體重等的相關性作了初步分析, 以期為這一珍貴物種的保護及管理提供基礎數據。
1 材料與方法
1.1 采樣地點及方法
采樣點位于江蘇省靖江市新橋鎮的長江沿岸江灘濕地(31°56"N, 120°01"E), 其沿岸上緣為混凝土堤壩, 下緣的堤腳為寬約30 m的大型石塊帶。石塊帶下緣至泥質江岸內側有寬約100 m的密集蘆葦(Phragmites australis)帶, 夏秋季蘆葦高約3 m, 冬天被革除。
垂直于江岸設置當地漁民所用典型“丁”字形定置張網一部, 攔網40 m長, 下網綱埋于江灘, 網片露出江灘高約1.8 m, 在攔網下緣近江心處布置2個籠式網袋。攔網網目為1.86 cm; 網袋網目為0.92 cm。張網位置隨季節性的水位漲落而上下移動, 一般控制在平水期水面與攔網的上綱齊平。當網片破損或藻類附著過多時, 更換同一規格的張網。每天由專人在早潮退潮后收集一次漁獲物。2002年1月至2003年12月, 每月保存1日、15日的2次樣本;2004年1月至2017年12月, 每月均保存1日、11日和21日的3次樣本。由于暴雨、洪水等原因, 造成部分樣本缺失, 16年共獲得519份漁獲物。樣本現場用10%福爾馬林溶液固定后, 帶回實驗室分析。
1.2 實驗方法
從每份漁獲物中分離出日本鰻鱺作為研究對象, 用量魚板逐尾測量全長L, 精確到0.1 cm; 干凈紗布吸干魚體水分后, 用電子天平稱量體質量W,精確到0.01 g。采用冪函數擬合全長和體質量的關系, 表達式為:。式中:W表示魚體質量(g);L表示魚的全長(cm);a為生長的條件因子;b為冪指數。
觀測并詳細記錄每尾樣本頭部、軀干部及胸鰭的體色, 依據郭弘藝等[20]對日本鰻鱺體色特征描述和評判標準, 鑒定每尾標本所處的發育期。挖取矢耳石作為年齡鑒定材料, 依據謝正麗等[21]的日本鰻鱺耳石制備、年輪確認和計數方法, 制備耳石磨片并鑒定樣本的年齡, 耳石結構和年輪標志見圖1。

圖1 日本鰻鱺矢耳石橫截面的結構和年輪Fig.1 Structure and annuli character on transversal section of the sagittal otolith of Anguilla japonica collected at the Jingjiang section of the Yangtze River
1.3 數據分析
采用Kruskal-Wallis檢驗對樣本年齡、全長、體質量在時間尺度上的差異顯著性進行檢驗。采用互相關函數(Cross-correlation function)對靖江段沿岸日本鰻鱺豐度與長江口鰻苗數量的時間序列進行互相關分析(Cross-correlation analysis), 長江口鰻苗為單船年捕撈量(尾/年), 時間分辨率為年, 數據來源于長江三峽工程生態與環境監測公報。數據處理和分析、圖形繪制用SPSS Statistics 20和GraphPad Prism 8.0軟件進行。
由于豐度較低, 0值較多, 本研究以日本鰻鱺的月出現率作為響應變量建立GAM模型, 分析各環境因子包括輸沙量(萬噸/月)、徑流量(m3/s)、水溫(℃)、潮差(m)、氣象因子氣壓(hPa)、降水量(mm)、渾濁度(NTU)、化學需氧量CODMn(mg/L)和氨氮(mg/L)等的影響。GAM表達式為:

式中, 函數g()為聯結函數, IO為日本鰻鱺的月出現率,為常數截距項。模型中的輸沙量和徑流量選取距離靖江段最近的長江干流大通水文站的數據, 數據來源于水利部長江水利委員會發布的長江泥沙公報。表層水溫和潮汐數據(水位基準為吳淞高程)來源于靖江市水利局, 潮差為2次漲落潮潮差的平均值。氣壓、降水量來源于靖江市氣象局;渾濁度、化學需氧量CODMn和氨氮來源于靖江市生態環保局。環境因子的時間尺度與日本鰻鱺出現率對應, 為2002年1月至2017年12月, 時間分辨率為月。
利用池信息量準則(AIC, Akaike Information Criterion)檢驗逐步加入因子后模型的擬合度,AIC值越小, 模型擬合度就越好[22,23]。根據F檢驗評估因子的顯著性[22]。最終GAM模型采用樣條平滑法,分布函數族為二項式分布, 模型構建和檢驗采用R 3.1.1軟件中的mgcv包實現[22]。
2 結果
2.1 豐度和出現率的時間尺度變化
分析顯示, 在519次漁獲物中, 日本鰻鱺出現了113次, 總體出現率為21.8%。共采集到日本鰻鱺137尾、8181.61 g, 僅占漁獲物數量和重量的0.08%和0.69%。
年際分析顯示, 在長江靖江段沿岸, 每年均采集到日本鰻鱺個體, 年均豐度為(9±4.2)尾。其中,2010年之前的年均出現率為28.2%, 2002年曾高達50.0%; 但此后7年的出現率大幅降低, 年平均僅為15.1%, 總體呈波動式遞減的趨勢(圖2)。
月際比較顯示, 日本鰻鱺在5—11月的出現率較高, 12月至次年3月的出現率較低(圖3)。此外,2010年以前每年平均有6.0月可采集到樣本, 2010年以后僅4.1月可采集到樣本, 同時出現率呈明顯的季節性后移, 1—4月未曾采集到樣本。
2.2 生物學特征
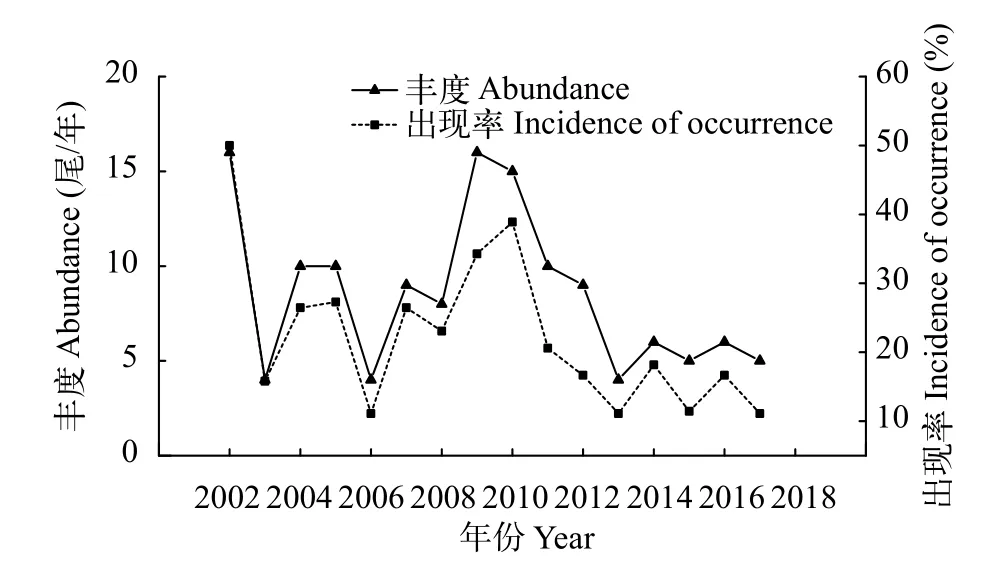
圖2 2002—2017年長江靖江段沿岸日本鰻鱺豐度和出現率的年際變化Fig.2 The abundance and incidence of occurrence of Anguilla japonica collected at the Jingjiang section of the Yangtze River during 2002—2017
采集到的137尾個體全長在17.0—65.0 cm, 平均為(31.1±9.8) cm; 體質量在5.03—499.50 g, 平均(57.64±91.94) g; 豐滿度在0.80—2.38, 平均為1.23±0.30(n=137)。據全長和體質量數據,擬合所得關系式為:W=0.0002415×L3.483(r2=0.9537,n=137,圖4)。經耳石磨片鑒定年齡的126尾個體, 年齡在1—6齡, 平均(1.8±1.0)齡。
時間尺度分析顯示, 個體全長(P=0.387>0.05)和體質量(P=0.560>0.05)在年際間均無顯著差異,但在月間存在顯著差異(P<0.05)。由圖5可見, 全長大于45 cm、體質量大于200 g的樣本僅出現在9—11月的成鰻降海洄游季節。年齡的月分布圖亦顯示, 1齡和2齡個體分別占總樣本數的48.4%和40.5%, 3齡及以上樣本僅占11.1%, 且僅出現在9—12月(圖6)。
依據性腺形態, 僅有15尾個體可鑒定性別(8尾雄性、7尾雌性), 其他89.1%的樣本均為未成熟個體。依據體表的色素發育程度判別, 樣本中有黃鰻期130尾、銀化前期(Pre-silver stage)7尾, 分別占個體總數的94.9%和5.1%, 且銀化前期僅出現在10—11月。
2.3 豐度與環境因子的關系
廣義可加模型(GAM)分析顯示, 徑流量、泥沙量、潮差、氣壓、降水量和化學需氧量CODMn對日本鰻鱺的出現率影響均不顯著(P>0.05), 而水溫及氨氮和渾濁度對其出現率卻有顯著影響(P<0.05)。從偏差變化及AIC值可以看出, 模型中環境因子對日本鰻鱺出現率的總偏差解釋率為34.0%。其中以水溫影響最大, 偏差解釋率為25.8%, 水質指標氨氮和渾濁度偏差解釋率分別僅為4.0%和4.2%(表1)。
由圖7可見, 在10—25℃水溫區間內, 日本鰻鱺的出現率呈增加趨勢, 水溫至25℃左右, 出現率達到最大。而氨氮和渾濁度與出現率均呈負相關關系,即隨著水體中氨氮濃度和渾濁度的增大, 日本鰻鱺的出現率呈下降趨勢。當水體中氨氮高于0.35 mg/L、渾濁度大于130 NTU時, 日本鰻鱺幾乎就不出現。
3 討論
3.1 長江沿岸日本鰻鱺群體的組成
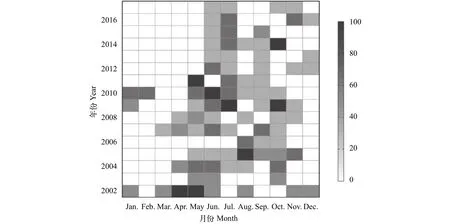
圖3 2002—2017年長江靖江段沿岸日本鰻鱺出現率的月際變化Fig.3 Monthly incidence of occurrence of Anguilla japonica collected at the Jingjiang section of the Yangtze River during 2002—2017
長江靖江段處于長江下游與河口段的交匯地帶, 受長江淡水徑流及海洋咸水潮汐的交互作用,形成了獨特的生態環境, 為多種魚類提供了育幼和索餌的良好場所[24]。其中的沿岸水域由于具有獨特的自然資源特征和生態系統過程, 具有很高的生態服務價值, 在維持河口魚類多樣性和漁業資源上具有獨特的作用[25]。本文采集到的137尾日本鰻鱺平均全長為(31.1±9.8) cm, 體質量為(57.64±91.94) g,平均年齡僅(1.8±1.0) 齡。由此可見, 線鰻溯河成功、已在淡水育肥1—2齡的黃鰻是日本鰻鱺長江近口段沿岸群體的主要成分, 也預示著長江沿岸是日本鰻鱺幼魚良好的棲息和索餌場所之一。已有研究表明, 在日本鰻鱺生殖洄游過程中消化道會逐步萎縮, 也不再攝食, 洄游所需的能量消耗來源于洄游前的積累[26,27]。但本研究顯示, 10—11月間有部分已啟動生殖洄游的銀化前期個體出現本研究水域, 表明日本鰻鱺在降海生殖洄游的初期也需要沿岸生境的依托。

圖4 2002—2017年長江靖江段沿岸日本鰻鱺全長體質量關系Fig.4 The relationship between body weight and total length of Anguilla japonica collected at the Jingjiang section of the Yangtze River during 2002—2017

圖5 2002—2017年長江靖江段沿岸日本鰻鱺全長和體質量月際小提琴變化圖Fig.5 The monthly violin plot of total length (a) and body weight(b) of Anguilla japonica collected at the Jingjiang section of the Yangtze River during 2002—2017

圖6 2002—2017年長江靖江段沿岸日本鰻鱺年齡月際小提琴變化圖Fig.6 The month violin plot of age of Anguilla japonica collected at the Jingjiang section of the Yangtze River during 2002—2017
全長-體質量關系式中的特征參數b反映魚類在不同生活史階段、季節和環境中的生長情況[28]。本研究發現長江沿岸生境鰻鱺群體的b值達3.483,高于福建萩蘆河的黃鰻群體(b=3.268)[29], 也高于同一水域敞水生境的銀鰻群體(b=3.246)[21]及珠海口(b=3.051)[30]和日本濱名湖(b=3.400)的銀鰻群體[31](表2)。可見, 長江更適合于日本鰻鱺的生長和肥育。
3.2 長江日本鰻鱺資源動態
大量研究顯示, 由于圍湖造田、江湖阻隔、水環境污染及過度捕撈等人類活動, 造成了世界各地的日本鰻鱺自然種群均處于枯竭狀態[9,19,32]。本研究連續16年的監測結果顯示, 日本鰻鱺在靖江段沿岸魚類漁獲物的出現率為21.8%, 僅占漁獲物數量與重量的0.08%和0.69%, 資源相當匱乏。這一結果與帥方敏等[9]對珠江水系的調查結果相似。雖然長江三峽工程生態與環境監測公報顯示, 2002—2017年長江口鰻苗單船捕撈量平均高達19161尾/年(圖8),資源量并不貧乏[32], 但由于日本鰻鱺的生活史很復雜, 真正能溯河至長江并且能發育至黃體鰻的數量極有限[10]。互相關分析亦證實, 靖江段沿岸當年日本鰻鱺豐度與前1—2年進入長江口的鰻苗數量并沒有顯著的相關性(圖9)。
近年來, 隨著長江禁漁期的實施和水產種質資源保護區的建立, 長江靖江段沿岸的刀鱭、胭脂魚和烏鱧等的幼魚資源都有增長的趨勢[34]。但本研究分析卻顯示, 日本鰻鱺的豐度和出現率均呈逐年波動式下降的趨勢。有研究指出, 大壩修建的阻隔、河流寬度與水深是造成鰻鱺空間分布結構差異的最主要因子[9]。但本文的GAM分析結果顯示,水溫、氨氮和渾濁度是造成日本鰻鱺的出現率差異的主要影響因子。由于靖江沿岸水域近年岸線圍墾、堤岸硬化和碼頭建設的加劇, 河岸植被大多消失, 加之污染物的超標排放, 漁業資源維持的生態服務功能也正受到威脅[35]。值得注意的是, 環境因子對日本鰻鱺出現率的解釋率僅為34%, 其他影響其資源波動的因子仍有很多, 有待于更深入研究。此外, 本研究僅分析了靖江段沿岸的一個調查點, 難以有效評估整個長江水系的資源狀況和時間分布特征。
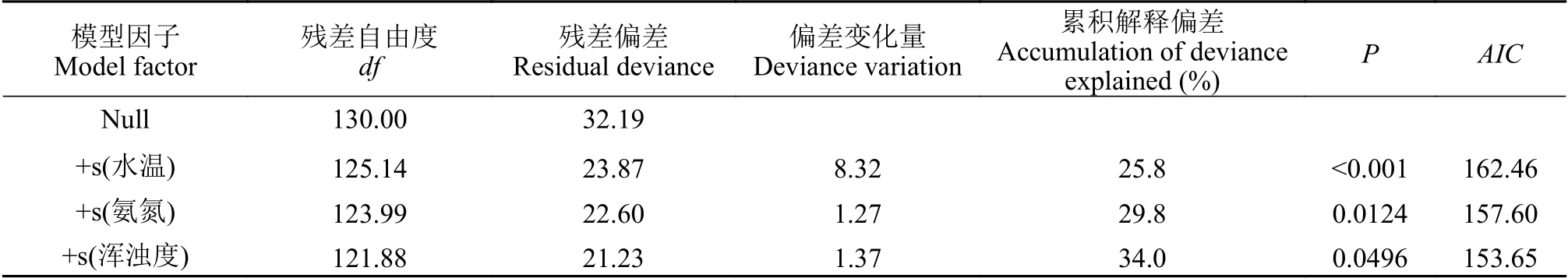
表1 長江靖江段日本鰻鱺出現率與環境因子的GAM檢驗Tab.1 Significance of explanatory variables effects on incidence of occurrence of Anguilla japonica collected at the Jingjiang section of the Yangtze River as determined by generalized additive model

圖7 廣義可加模型(GAM)所揭示的顯著影響長江靖江段日本鰻鱺出現率的環境因子Fig.7 Generalized additive model (GAM)-derived effects on incidence of occurrence of Anguilla japonica collected at the Jingjiang section of the Yangtze River

表2 不同水域的日本鰻鱺生長參數比較Tab.2 Comparison of growth parameters of Japanese eels in different waters

圖8 2002—2017年長江口日本鰻鱺鰻苗單船年捕撈量Fig.8 Annual catch of Anguilla japonica glass eels collected on the Yangtze River estuary during 2002—2017

圖9 長江靖江段沿岸日本鰻鱺豐度與長江口日本鰻鱺鰻苗單船年捕撈量間的互相關分析Fig.9 Cross-correlation analysis of Anguilla japonica abundance collected at the Jingjiang section of the Yangtze River and annual catch of Anguilla japonica glass eels collected on the Yangtze River estuary during 2002—2017
