海拔對烏紅雜交天麻產量與品質的影響及其酶學作用機制
2021-03-29 07:54:48王傳華曾春函王洋洋侯云波
西北植物學報 2021年2期
關鍵詞:生長
楊 婧,王傳華,曾春函,王洋洋,侯云波,楊 林
(1 三峽庫區生態環境教育部工程研究中心,湖北宜昌443002;2 湖北省三峽地區生態保護與治理國際聯合研究中心,湖北宜昌443002;3 三峽大學生物與制藥學院,湖北宜昌443002;4 貴州健康職業學院,貴州銅仁 554300)
天麻(GastrodiaelataBl.)是一種具有很高藥用價值的異養型蘭科植物[1],需要與萌發菌、蜜環菌共生才能完成生活史[2]。蜜環菌隸屬于擔子菌門(Basidiomycota)傘菌目(Agaricales)小皮傘科(Marasmiaceae)蜜環菌屬[Armillaria(Vahl) P.Kumm.][3],廣泛分布于歐洲、亞洲、非洲、北美洲等地。蜜環菌具有兼性寄生特性,需從朽木或活樹根汲取營養[4-5],目前世界上已知有近40 個生物種[6],部分種是重要的食藥用真菌[7]。天麻與蜜環菌具有特殊的共生關系,天麻球莖一方面依賴于真菌的侵染而獲得營養,另一方面又具有強有力的阻止或限制真菌侵染的防衛機制以避免自身感病[8]。
海拔(溫度)是影響天麻生產的重要因素。天麻適宜在夏季涼爽且冬季溫和的環境中生長[9],其自然分布具有明顯的海拔地帶性,垂直分布的高度由南到北逐漸下降[10],適宜溫度為-4~20 ℃[11]。外界溫度為14 ℃左右時,天麻開始生長;升高到20 ℃時,天麻進入迅速生長期;但達到28 ℃時,溫度會抑制天麻的生長,因此,天麻的最適生長溫度是15~24 ℃。另一方面,當溫度低于8 ℃或高于28 ℃時,天麻將停止生長,進入休眠狀態[10,12-14]。生產實踐中,在低海拔地區栽培天麻常因蜜環菌過量繁殖導致其腐爛[15],同時海拔不同也導致藥用化學物質含量不同[16]。因此,深入研究溫度對天麻-蜜環菌共生關系的影響并揭示其作用機制,對提高天麻產量、改善天麻品質具有重要意義。
天麻抵御蜜環菌的入侵機制中,多種消化酶和抗真菌蛋白扮演著重要角色。研究表明,天麻皮層的抗真菌蛋白(GAFP)是抵御蜜環菌入侵的主要物質[17-20],而皮層細胞與消化細胞溶酶體細胞釋放的消化酶及幾丁質酶(EC3.2.1.14)在消化入侵菌絲流過程中發揮關鍵作用[10,21-22]。研究還發現,幾丁質酶作為一種糖苷水解酶,在蜜環菌的生長發育及其與天麻的共生過程中發揮重要作用[23-25],而β-1,3-葡聚糖酶(EC3.2.1.39)在天麻初生球莖消化蜜環菌菌絲的過程中起重要作用[13]。同時,植物病理學的研究表明,一些酶類如多酚氧化酶(PPO,EC1.10.3.1)、苯丙氨酸解氨酶(PAL,EC4.3.1.5),也參與植物的抗病過程。王曼玲等認為,PPO在水稻、煙草、棉苗及蘋果抵御各種病菌的過程中起重要作用[26],而植株感染真菌后PPO 活性明顯增強,且其活性沿著感染部位到未感染的邊緣有明顯的梯度變化[27]。另外,當植物受到病原菌的侵染時,PAL活性也有明顯的變化,因此,PAL活性的變化與植物的抗病性也密切相關[28]。迄今,溫度變化對天麻抗侵染及消化酶系統影響的研究仍未見報道,有關植物抗病酶類是否在天麻-蜜環菌的共生機制中扮演角色尚不清楚。
那么,天麻的產量及品質隨海拔升高呈現何種變化趨勢?溫度升高對天麻體內消化酶、防御酶及組織液抑制菌絲體生長的活性有何影響?進而對天麻-蜜環菌共生關系有何影響?為了回答以上問題,本研究以在中國廣泛種植的烏紅雜交天麻作為實驗材料,在不同海拔開展天麻栽培實驗,以考察海拔影響天麻生物量與藥用品質(天麻素、對羥基苯甲醇含量)的效應;同時,在室內控制條件下測定溫度變化對天麻消化酶、防御酶活性,以及天麻組織液抗蜜環菌菌絲體生長效果的影響,揭示溫度對天麻-蜜環菌共生體的生長與存活的作用機制。本研究結果對于揭示蜜環菌-天麻的共生機制具有重要科學價值,并對天麻栽培的區劃具有重要實際指導意義。
1 材料和方法
1.1 研究區域概況
本研究選擇湖北宜昌市五峰縣五峰鎮和牛莊鄉開展天麻田間栽培實驗。近30年來,該地區已經形成了成熟的天麻栽培管理技術體系,是中國著名的天麻主產區之一。試驗場地處鄂西南的武陵山余脈,境內山地資源豐富,最高海拔達到2 320.3 m,年均日照1 533 h,年均氣溫13~17 ℃,無霜期240 d,年均降水量1 600 mm,部分地區擁有獨特的沙性壤土。
1.2 試驗材料
供試的天麻種質為烏紅雜交天麻。父本烏天麻(G.elataB1. f.glauca)和母本紅天麻(G.elataB1. f.elata)種源均為野生天麻的自交系。根據前期實驗結果,選擇CBS F二倍體菌株用于培養天麻栽培用蜜環菌材;選擇CBS A、CBS B、CBS D、CBS F、CBS H、CBS J、CBS M和CBS N的單倍體菌絲作為天麻組織液抑菌效果的蜜環菌株(表1)。
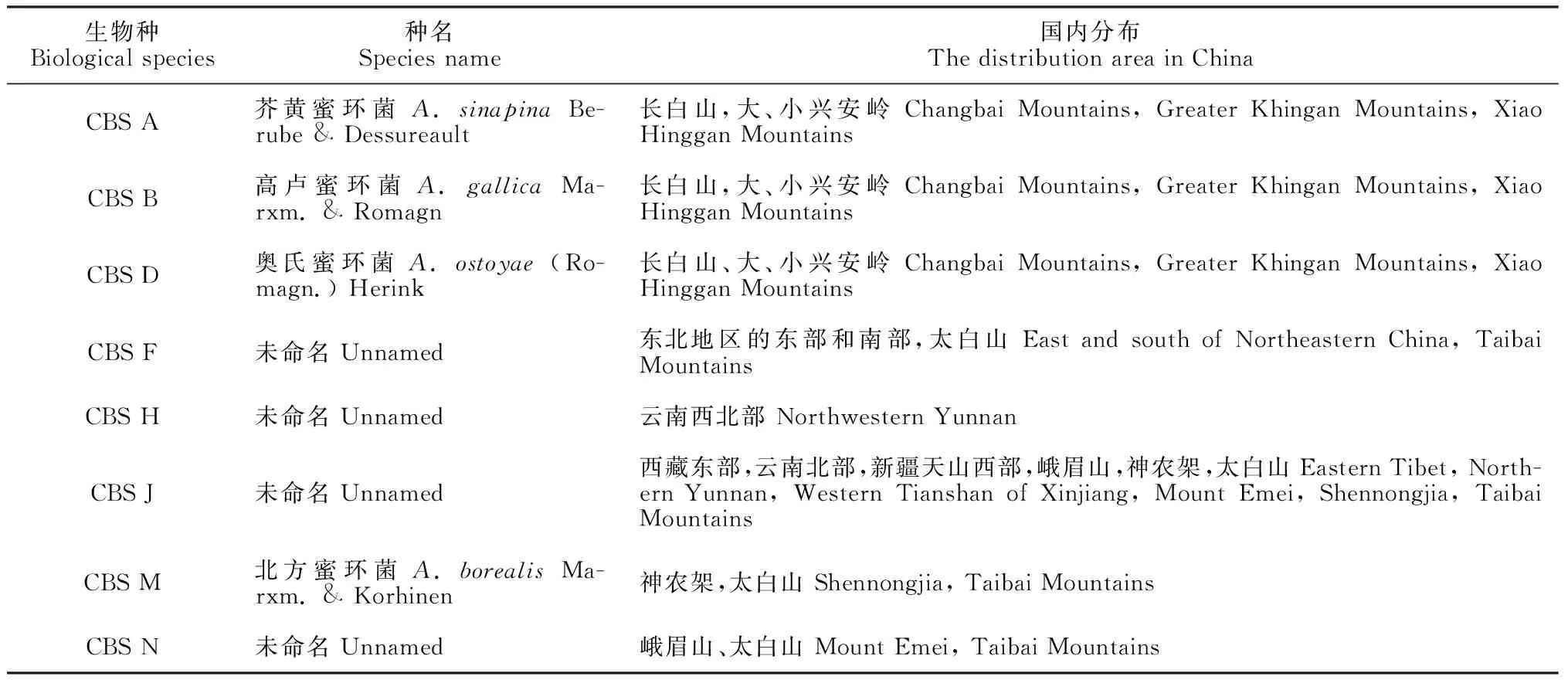
表1 參試天麻組織液抑制蜜環菌菌絲體生長試驗的蜜環菌生物種
1.3 海拔對天麻生物量與品質的影響試驗
1.3.1 實驗設計本試驗采用單因素3水平設計,以海拔為固定因子。由于中國雜交天麻適生海拔為800~1 900 m[27],故在研究區域內五峰縣五峰鎮、牛莊鄉選擇850 m、1 600 m和1 900 m 三個海拔梯度,地勢較為平坦、坡向一致的撂荒農地(5年以上)設置試驗場。
1.3.2 大田栽培2018年元月,采用熟料段木法準備蜜環菌材。具體方法為:選擇長20 cm、直徑約15 cm、鮮重約2 500 g(±50 g)的白櫟(QuercusfabriHance)段木,裝入聚氯乙烯袋常壓滅菌8 h,接種CBS F后在25 ℃下培養60 d備用。烏紅雜交天麻種麻的質量約10 g/頭,用種量每段木為m0(約50 g)。2018年4月初,參考徐錦堂的方法[10],采用單棒法栽培天麻。行寬30 cm,每根菌材作為一個處理單位,處理間距離30 cm,每個處理20個重復。
1.3.3 生物量測定2018年10月收獲栽培的天麻。以菌棒為單位,測定天麻的鮮重(m1)和個體數,計算增殖率[G= (m1-m0)/ m0×100%]。帶回實驗室后,分揀個體鮮重大于50 g的所有個體,用天平(精度0.01g)測定總鮮重(W1),然后在常壓下將天麻樣品蒸制20 min,在60 ℃的烘箱內烘干至恒重[28],用天平(精度0.01 g)測定干重(W2),計算折干率(D= W2/ W1×100%)。將每一菌棒產生的天麻烘干后,粉碎過60目篩,烘干后低溫保存,用于天麻素和對羥基苯甲醇的含量測定。
1.3.4 天麻有效藥用成分的測定取樣品粉末,準確稱定1.0 g,置于具塞錐形瓶中,用移液管加入稀乙醇25 mL,超聲處理30 min,放至室溫,5 000 r/min離心20 min;取上清液5 mL,在50 ℃旋蒸濃縮至近干無醇味,殘渣加乙腈-0.05%磷酸溶液(5∶95)混合溶解,定容至10 mL容量瓶,用1 mL一次性無菌注射器吸入,從0.22 μm的微孔濾膜濾頭過濾至EP管。從EP管中吸取15 μL樣品液,注入色譜儀進樣器中,檢測波長為220 nm,測定峰面積[29]。
精密量取對照品天麻素和對羥基苯甲醇原溶液適量,加乙腈-0.05%磷酸溶液(5∶95)溶液制成質量濃度梯度的溶液,依次測定出峰面積,建立峰面積(y)與質量濃度(x)間的標準曲線及其方程(表2)。根據樣品的出峰面積按標準曲線計算樣品的天麻素和對羥基苯甲醇含量。根據每棒的天麻總鮮重(m1)、增值率(G)和天麻樣品藥用成分含量C(%)計算每菌棒所產天麻含有的天麻素總量、對羥基苯甲醇總量及二者合量。

表2 天麻素和對羥基苯甲醇標準曲線
1.4 溫度對天麻抗蜜環菌侵染及消化系統的影響試驗
1.4.1 天麻消化酶和防御酶活性對溫度的響應1)天麻樣品的采集 2018年10月,從海拔1 900 m、1 600 m和800 m三處野外種植的烏紅雜交天麻中隨機選取5窩,每窩取100 g同一規格(長約2~3 cm大小)的白麻放入自封袋,置-80 ℃保存[30],用于背景值測定;另在海拔1 600 m處(3個海拔點表現最好)的每窩取100 g規格一致的白麻用于溫度脅迫實驗。2)溫度處理 將天麻樣品置于15 ℃、25 ℃和30 ℃的濕潤河沙中暗培養7 d[31-32]。 3)粗酶液的制備 分別參考Boller T[33]、史益敏[34]、張溫典[35]及胡月[36]的研究方法制備β-1,3-葡聚糖酶、幾丁質酶、多酚氧化酶(PPO)、苯丙氨酸解氨酶(PAL)的粗酶液。4)酶活性測定 采用用酶免ELISA試劑盒測定4種酶的活性(江蘇酶免實業有限公司,批號LOT NO.:1909P)。將試劑盒標準品稀釋成濃度梯度為12.5、25、50、100、200 U/L的標準品稀釋液,根據說明書步驟進行實驗測定,以標準物濃度為橫坐標(x),其吸光度值(OD)為縱坐標(y),繪制標準曲線(表3)。然后,測定樣品的OD值,計算各酶的活性。

表3 4種酶的標準曲線方程與活性計算公式
1.4.2 不同培養溫度天麻粗提物對蜜環菌生長的抑制作用1)天麻材料收集與處理 2018年10月選取野外海拔1 600 m處栽培(3個海拔點表現最好)的烏紅雜交天麻3窩,每窩選取長度約2~3 cm的白麻1~1.5 kg,4 ℃保存備用。2)溫度處理 設15 ℃、25 ℃和30 ℃ 三個溫度梯度[39]。每個處理約700 g鮮天麻,裝入透氣的泡沫盒中,處理時濕沙覆蓋[38],處理時間1周。每處理3個重復。3)天麻組織粗提液制備 參考胡忠等的方法提取天麻具有抗真菌蛋白活性的組織粗提液[18,39]。將提取液分為兩部分,2/3體積提取液用于實驗組,1/3的提取液121 ℃高溫高壓滅菌20 min,作為對照組;試驗和對照組均采用0.22 μm微孔濾膜抽濾制備無菌濾液,得到抗真菌蛋白(GAFP)粗提液。4)抑菌效果測定 在無菌環境下將抗真菌蛋白粗提液添加到PDA培養基中[20,40],并接入8個種的蜜環菌生物種單倍菌絲體(表1),放入恒溫培養箱24~25 ℃恒溫培養20 d。然后用掃描儀(EPSON PERFECTION 4870 PHOTO)掃描并用根系分析系統WinRHIZO測定菌落面積,計算菌落面積變化比率(HC),HC=(Si-Sa)/Sa×100,其中Si和Sa分別為試驗組和對照組各蜜環菌生物種的菌落面積。將長有蜜環菌單倍體的培養基放入沸水中煮至培養基融化,然后將菌索65 ℃烘干至恒重,稱重。計算生物量變化比率(HB),HB=(Bi-Ba)/Ba×100,其中Bi和Ba分別為試驗組和對照組的菌索生物量。
1.5 數據分析
以海拔為固定因子,采用SPSS20.0軟件對增長率(G)、折干率(D)、天麻素含量、對羥基苯甲醇含量和總藥用成分含量(S,天麻素含量+對羥基苯甲醇含量)進行單因素方差分析;采用Duncan進行多重比較,顯著性水平設置為0.05。另外,對β-1,3-葡聚糖酶、幾丁質酶、PPO、PAL活性、蜜環菌菌落面積、生物量及抑菌效果,以溫度為固定因子,采用SPSS22.0進行單因素ANOVA分析,然后采用鄧肯法進行處理間顯著性差異檢驗(P<0.01);采用Origin2018作圖。
2 結果與分析
2.1 海拔對烏紅雜交天麻生長及藥用品質的影響
實際栽培結果表明,海拔烏紅雜交天麻的增長率、天麻素含量及總對羥基苯甲醇含量存在顯著影響,而對折干率無顯著影響(表4)。其中,烏紅雜交天麻增長率在海拔1 600 m最高,并顯著高于其他海拔,分別比海拔850 m 和 1 900 m顯著提高64.06%和98.93%;天麻素含量卻在1 900 m海拔處最高,并顯著高于其他海拔,分別比海拔850 m 和 1 600 m顯著提高20.1%和25.13%; 總對羥基苯甲醇含量在1 600 m處最高,分別比海拔850 m 和 1 900 m顯著提高46.15%和40.74%。綜合而言,烏紅雜交天麻有效成分總量在海拔1 600 m處最高,分別比海拔850 m和1 900 m提高31.10%和8.39%,但差異均未達到顯著水平。可見,海拔1 600 m環境最有利于烏紅雜交天麻的生長和藥用有效成分的積累。
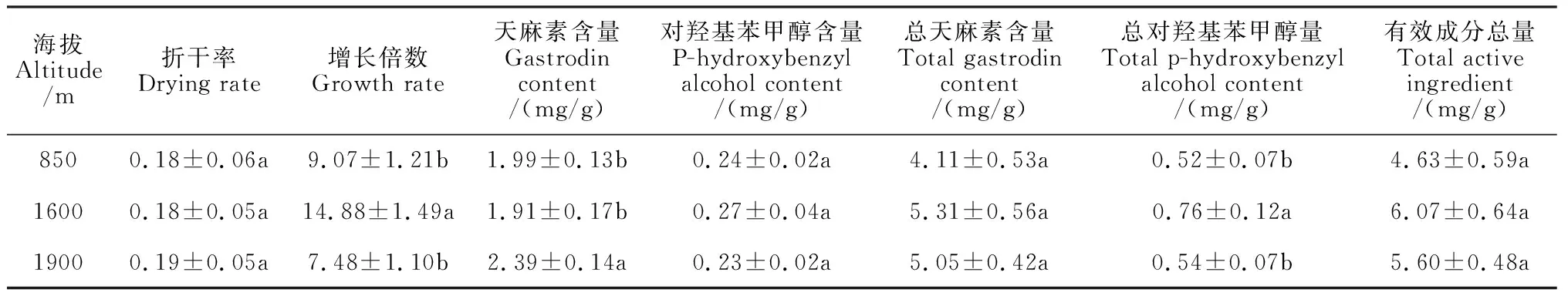
表4 不同海拔下雜交天麻生長與藥用品質的變化
2.2 培養溫度對烏紅雜交天麻體內消化酶與防御酶活性的影響
圖1顯示,烏紅雜交天麻體內消化酶(幾丁質酶、β-1,3-葡聚糖酶)活性隨溫度而降低,表現為15 ℃處理顯著高于25 ℃和30 ℃處理,這表明高溫顯著破壞了雜交天麻的消化酶活性。同時,β-1,3-葡聚糖酶活性對溫度的反應更敏感,受溫度的影響更大,且在各溫度處理間具有顯著性差異(圖1)。雜交天麻體內多酚氧化酶(PPO)和苯丙氨酸解氨酶(PAL)活性在不同溫度下也差異顯著。PPO活性隨著溫度的上升而逐漸增強,且各溫度處理間均存在顯著性差異;PAL活性隨著溫度的升高先下降然后上升,且25 ℃處理顯著低于15 ℃和30 ℃處理(圖1)。以上結果表明烏紅雜交天麻各防御酶活性在溫度升高時變化趨勢不盡相同,PPO活性具有增強趨勢,而PAL活性卻有降低趨勢。
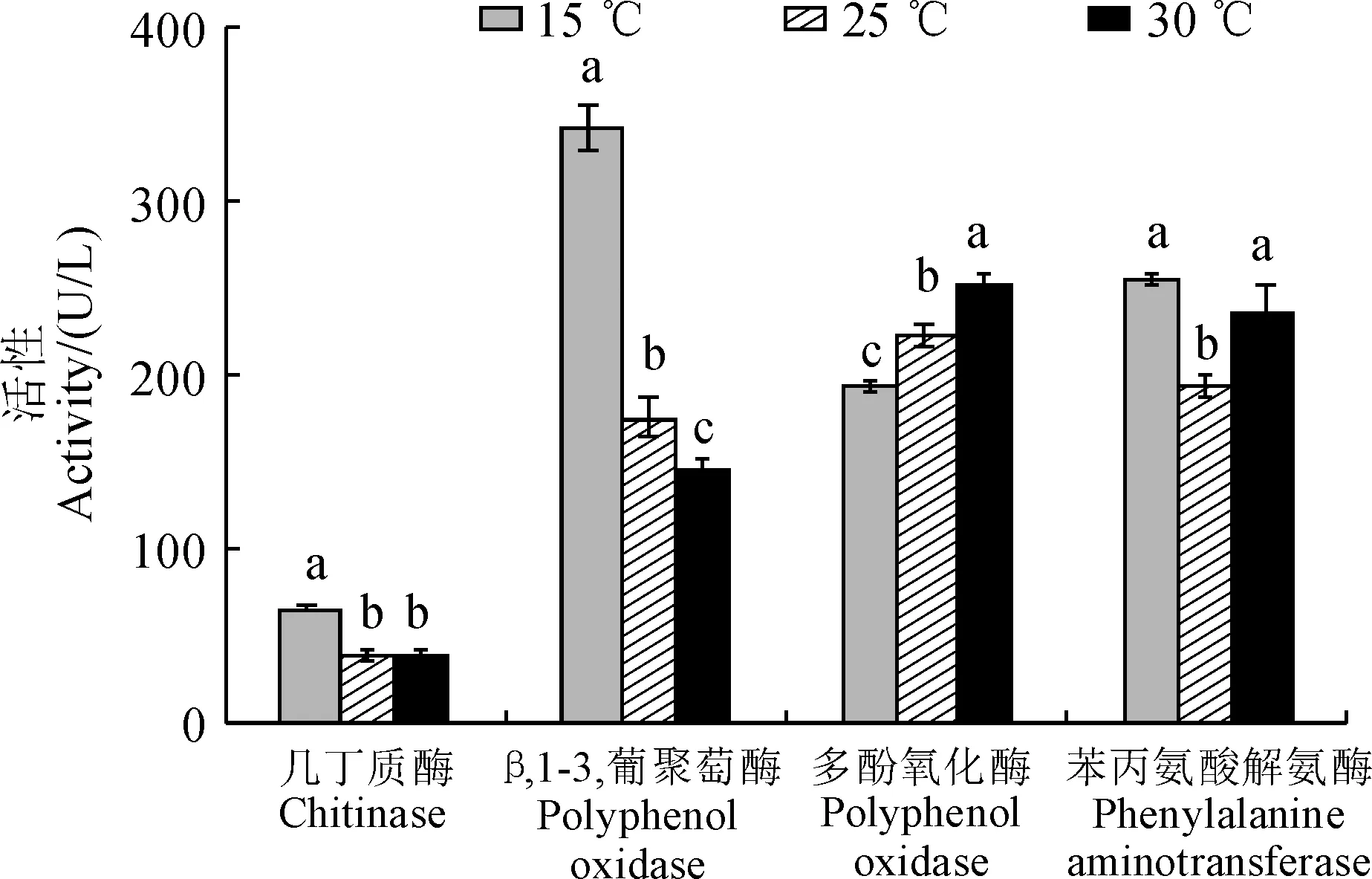
每種酶內不同小寫字母表明溫度處理間在0.05水平上存在顯著性差異(Duncan,P<0.05)
2.3 培養溫度對天麻組織粗提液抑制蜜環菌生長的影響
圖2,A顯示,在不同培養溫度天麻提取物(抗真菌蛋白,GAFP)處理下,各蜜環菌菌種單倍體菌絲菌落面積變化率表現復雜多樣,且溫度處理間均存在顯著性差異;菌落面積變化率不僅有上升和下降差異,更有正值和負值之別。其中,在各溫度提取液處理下,菌種CBSA菌落面積變化率均為正值,菌落生長得到促進;而菌種CBSF和CBSG菌落面積變化率均為負值,菌落生長均受到抑制;其余菌種的菌落面積變化率既有正值,也有負值,說明菌落生長有些情況下得到促進,有些情況受到抑制。在15 ℃培養天麻提取物處理下,菌落面積變化率僅菌種CBSA和CBSN為正值,其余菌種菌落面積變化率為負值,即多為抑制作用且大多較大;在25 ℃培養天麻提取物處理下,菌落面積變化率除菌種CBSF和CBSJ外均為正值,即多為促進作用;30 ℃培養天麻提取物處理下,菌落面積變化率僅菌種CBSA、CBSH和CBSM為正值但促進作用很明顯,其余菌種均為負值但抑制作用相對較小。可見,大多數蜜環菌菌種菌落生長的抑制效果均以15 ℃培養天麻提取物處理最顯著,隨著培養天麻溫度的上升,抑菌效果有顯著降低的趨勢,但其抑制效應在8個生物種間存在顯著差異。
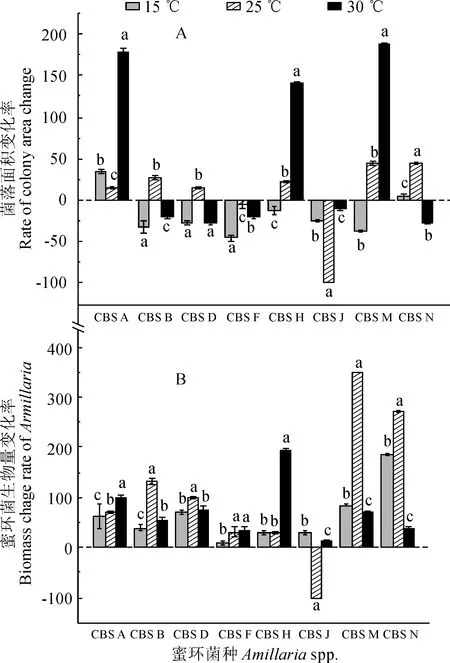
正值代表促進生長,負值代表抑制生長;同一菌種內不同小寫字母代表溫度處理間在0.05水平存在顯著性差異(Duncan,P<0.05)
同時,與試驗預期相反,菌落生物量的變化率除CBSJ外均為正值,即未滅活天麻組織液(天麻抗真菌蛋白)均明顯促進各蜜環菌菌種的生長,但不同培養溫度組織液的促進效應有顯著差異,在低溫時促進效應較弱,而在中溫時促進效應相對較強(圖2,B)。其中,除菌種CBSJ以外,隨著培養溫度的升高,大部分蜜環菌生物量先增加后降低,也就是天麻提取物對生物量增加的相對抑制能力先降低后升高,在25 ℃達到最低值,大多數在15 ℃達到最高值。上述結果表明,盡管天麻組織液中存在具有抑菌活性的抗真菌蛋白(GAFP),但天麻組織粗提液中可能同時存在對高溫敏感的未知活性物質,二者綜合作用呈現出對蜜環菌生長具有促進作用。
3 討 論
經過實驗地栽培研究,發現在海拔1 900 m處由CBS F伴栽的紅天麻有效成分含量最高,雜交天麻適合種植在1 600 m海拔范圍。也有學者發現,無論在原位還是模擬條件下,植物與真菌共生體雙方的海拔來源及海拔梯度上的環境(或溫度)變化均顯著影響其生長[41]。結合彭華勝的研究結果[11],天麻的分布有一定海拔要求,其中西南地區以1 200~1 800 m較多,東北地區多分布在海拔300~1 000 m。而在鄰近緯度,低海拔地區溫度較高,高溫抑制了天麻中消化酶的活性,但是蜜環菌胞外酶活性隨溫度的升高先增加后降低,這就可能使天麻-蜜環菌的共生關系遭到破壞。
本研究的結果表明,天麻體內消化酶和防御酶系統對于溫度升高的響應具有差異。低溫下天麻消化酶活性升高,而高溫下天麻的保護酶活性則增加。早在20世紀80年代研究發現,天麻皮層細胞具有消化功能,菌絲的細胞壁就是在其中被消化的,發揮作用的主要是天麻皮層分泌的幾丁質酶、β-1,3-葡聚糖酶[42-43]。因此,這些消化酶活性的變化直接影響天麻的營養供給,也與天麻免受蜜環菌侵染密切相關。同時,本研究發現,高溫下天麻體內防御酶活性增強,這與戴明凱等以馬鈴薯為研究對象,發現高溫脅迫下馬鈴薯體內保護酶活性隨時間推移增加的結果相似[44]。因此,天麻響應蜜環菌侵染的策略隨著溫度的升高發生變化,由低溫的消化蜜環菌菌絲體供自生需要,轉向高溫下保護自身免受侵害為主。
與此同時,我們還研究了溫度變化對天麻體內組織粗提液抑制蜜環菌單倍體菌絲生長的影響,發現溫度升高削弱了天麻對蜜環菌菌絲體的抑制能力。本研究中采用的是天麻組織的粗提液,包含各種酶類和抗真菌蛋白以及其他成分不明的物質[45-47]。抗真菌蛋白(GAFP)是一類對真菌具有抑制作用的蛋白質,在芝麻菜種子和天麻的塊莖中均有發現[48-49]曾從新生麻塊莖中檢測并純化出一種抗真菌蛋白,并對其進行了免疫熒光定位,發現抗真菌蛋白定位于新生麻表皮及皮層細胞的外緣,是新生麻避免被蜜環菌侵入的物質基礎。而且GAFP是越冬前新生麻阻止蜜環菌侵入的化學防御物質[50],說明低溫對于GAFP的抑菌效果顯著,可能是高溫抑制GAFP的表達,或者活性。這與本試驗所發現的天麻組織粗提液在低溫下抑菌效果明顯較為一致。由于幾丁質酶和葡聚糖本身不具有抑菌活性[48],我們推斷本研究中發揮抑菌作用的主要物質是GAFP。本研究還發現,天麻組織粗提液中存在促進蜜環菌生長的活性物質,這種活性物質可能存在于天麻皮層中GAFP的外側,以誘導和促進蜜環菌趨向天麻組織的生長。這也說明天麻與蜜環菌在長期的協同進化過程中具有十分復雜的關系。
溫度還可能對天麻伴生蜜環菌的侵染特性產生影響。蜜環菌菌絲體最適宜的生長溫度為20~25 ℃,超過30 ℃停止生長[51],施漢鈺等也認為最佳的蜜環菌培養溫度為恒溫25 ℃[37]。也有學者研究發現,蜜環菌的生長量在高溫條件下顯著高于低溫條件下,其差異主要是菌絲生長,而菌索生長的差異并不大[52]。另外,溫度還影響蜜環菌胞外酶的分泌。如曾春函等在研究中發現,溫度對蜜環菌胞外漆酶和果膠酶活性大小有影響,對產酶時間沒有影響;對纖維素酶活性達到峰值的時間和活性大小都有影響[53]。
4 結 論
溫度是影響蜜環菌-天麻共生關系的重要環境因子。隨著溫度升高,天麻抵御蜜環菌入侵的策略發生變化,天麻的消化酶活性和粗提液的抑菌活性下降,但是保護酶活性上升。而且不同海拔下栽培的天麻產量及有效成分也有明顯差異,這種適應策略的轉變,可能是溫度升高導致蜜環菌病理性侵染天麻的重要機制。同時,天麻體內含有某種促進蜜環菌生長的活性物質,說明天麻體內同時存在促進蜜環菌生長和抑制蜜環菌侵染的復雜系統。而且經過研究發現,在海拔1 900 m處,由CBS F伴栽的紅天麻產量及有效成分含量均最高。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
