轉(zhuǎn)彩色馬鈴薯StAN1基因的煙草幼苗耐旱性分析
2021-03-29 07:49:14鮑燕如聶騰坤王冬冬
西北植物學(xué)報(bào) 2021年2期
鮑燕如,聶騰坤,王冬冬,陳 勤
(1 西北農(nóng)林科技大學(xué) 農(nóng)學(xué)院,旱區(qū)作物逆境生物學(xué)國(guó)家重點(diǎn)實(shí)驗(yàn)室,陜西楊陵 712100;2 西北農(nóng)林科技大學(xué) 食品科學(xué)與工程學(xué)院,旱區(qū)作物逆境生物學(xué)國(guó)家重點(diǎn)實(shí)驗(yàn)室,陜西楊陵 712100)
馬鈴薯(Solanumtuberosum)是茄科一年生草本作物,其中富含多種花青素,具有很好的營(yíng)養(yǎng)價(jià)值和保健價(jià)值[1],有利于預(yù)防和治療一些癌癥以及一些心血管疾病[2-3],是世界范圍內(nèi)重要的糧食作物之一。馬鈴薯在溫暖和寒冷的環(huán)境中都可以生長(zhǎng),但不耐旱。在整個(gè)生育時(shí)期都容易受到干旱脅迫[4],從而導(dǎo)致產(chǎn)量下降。
隨著全球氣候變化,選擇耐旱抗旱的馬鈴薯品種對(duì)于全球糧食安全至關(guān)重要[5]。干旱脅迫是農(nóng)業(yè)生產(chǎn)中最常見的非生物脅迫,而且植物對(duì)于這些脅迫的初始反應(yīng)都是相似的,由于水分不足以及活性氧的積累進(jìn)而抑制了植物的生長(zhǎng)和發(fā)育[6],在農(nóng)業(yè)生產(chǎn)上造成大規(guī)模的減產(chǎn)。因此,提高植物的耐旱性是應(yīng)對(duì)全球氣候變化較為行之有效的方法與措施[7-8]。LEA蛋白最早在棉花(Gossypiumhirsutum)種子中發(fā)現(xiàn),在種子成熟后期大量合成,因此被稱為胚胎發(fā)育后期豐富蛋白。隨后,研究人員陸續(xù)從大豆(Glycinemax)、玉米(Zeamays)等作物中也發(fā)現(xiàn)了這類蛋白。LEA蛋白除了在種子成熟后期表達(dá)外,在植物受到不同程度干旱脅迫時(shí)也會(huì)大量表達(dá),是由于LEA蛋白中的氨基酸賦予其較好的親水性,在應(yīng)對(duì)干旱等非生物脅迫時(shí)能有效保持水分[9]。而AP2/ERF類轉(zhuǎn)錄因子調(diào)控植物的生長(zhǎng)并廣泛參與植物的次生代謝[10-11],植物自身次生代謝產(chǎn)物的積累能夠有效地調(diào)控植物應(yīng)對(duì)生物或者非生物脅迫[12]。
花青素是一類重要的黃酮類次生代謝產(chǎn)物[8],相對(duì)于其他類黃酮物質(zhì)具有更高的抗氧化性,由于其含有帶正電荷的氧原子,清除活性氧的能力更強(qiáng),能有效防止細(xì)胞膜脂過氧化,減少活性氧對(duì)植物的損傷,提高植物在干旱脅迫下的存活率[13]。彩色馬鈴薯中富含多種花青素,參與多種生化反應(yīng)過程[14],如生物脅迫、植物激素運(yùn)輸以及不同器官色素沉積,在干旱所導(dǎo)致的非生物脅迫中,花青素還可以有效清除活性氧,保持細(xì)胞穩(wěn)態(tài)環(huán)境[15]。花青素表達(dá)受MYB轉(zhuǎn)錄因子(TFs)、堿性螺旋-環(huán)-螺旋(bHLH) TFs和WD40蛋白復(fù)合體(MYB-bHLH-WD40‘MBW’復(fù)合物)的控制[16-17]。StAN1基因是R2R3 MYB基因家族調(diào)控彩色馬鈴薯花青素合成的一個(gè)基因,在調(diào)控花青素合成過程中發(fā)揮重要作用[18-19]。因此,本研究探討了轉(zhuǎn)StAN1基因煙草在應(yīng)對(duì)干旱和高溫脅迫時(shí)的反應(yīng),為進(jìn)一步了解彩色馬鈴薯StAN1基因功能提供參考。
1 材料和方法
1.1 試驗(yàn)材料與試劑
試驗(yàn)材料為本氏煙草(Nicotianatabacum)及過表達(dá)StAN1轉(zhuǎn)基因煙草的穩(wěn)定株系,均由本實(shí)驗(yàn)室保存并提供。
試劑MS培養(yǎng)基(Murashige & Skoog)購于美國(guó)Phytotech公司,植物組DNA、RNA提取試劑盒以及熒光定量試劑盒均購于天根生化科技(北京)有限公司,2×Taq PCR StarMix購于北京康潤(rùn)誠(chéng)業(yè)生物科技有限公司,其他試劑均為國(guó)產(chǎn)分析純,購于廣州市華大化學(xué)試劑有限公司。
1.2 試驗(yàn)方法
1.2.1 轉(zhuǎn)StAN1基因煙草PCR檢測(cè)用DNA提取試劑盒提取T4代轉(zhuǎn)StAN1基因煙草的DNA。用鑒定引物進(jìn)行PCR擴(kuò)增,反應(yīng)體系50 μL,DNA模板2 μL,上、下游引物各2.5 μL(10 mmol/L),ddH2O 18 μL,2×Taq PCR StarMix 25 μL。反應(yīng)程序?yàn)椋?4 ℃預(yù)變性7 min;94 ℃變性30 s;51 ℃退火30 s;72 ℃延伸1 min,36個(gè)循環(huán);72 ℃延伸10 min,4 ℃保存。
鑒定引物為StAN1ox-F(5′-ATGACTTCACATGTAATGATCA-3′) 和 StAN1ox-R(5′-TTA-ATTAA GTAGATTCCATATATCA-3′)。
1.2.2 轉(zhuǎn)StAN1基因煙草qRT-PCR檢測(cè)提取PCR結(jié)果為陽性的煙草總RNA,按照天根反轉(zhuǎn)錄試劑盒進(jìn)行反轉(zhuǎn)錄,參照天根熒光定量試劑盒說明書,以20 μL 體系進(jìn)行實(shí)時(shí)熒光定量檢測(cè)轉(zhuǎn)基因煙草。特異性引物為actin-F(GCTTTCTTCGTCCCATCA)和actin-R(CCCCAAGTACCCTCGTAT);StAN1-F(GCAAGCCAATGCCATAATAAGA)和StAN1-R(ATTCATCCCAACCACCATCACC)。
1.2.3 轉(zhuǎn)StAN1基因煙草花青素含量測(cè)定采用pH示差法測(cè)定葉片中的花青素含量[20-21],取0.1 g煙草葉片,液氮研磨后,用2 mL 70%乙醇提取,10 000 r/min離心15 min,取500 μL上清液,分別用pH1.0和pH4.5的緩沖液稀釋4倍,40 ℃平衡30 min,用紫外分光光度計(jì)分別于525和700 nm測(cè)定吸光度值。
花青素含量(mg/g)=[(A525-A700)pH1.0-(A525-A700)pH4.5]×MW×DF×1 000/(ξ×m)
公式中A為吸光度值,MW為矢車菊素葡萄糖苷相對(duì)分子量449.2,DF是稀釋倍數(shù),ξ是矢車菊素葡萄糖苷摩爾消光系數(shù)26 900,m為樣品的質(zhì)量。
1.2.4 植物材料處理過表達(dá)StAN1基因的煙草種子和野生型煙草種子,經(jīng)過10%次氯酸鈉消毒后,用無菌水沖洗3~5次,分別點(diǎn)在甘露醇濃度為0、50、100、150和200 mmol/L的MS培養(yǎng)基上,置于25 ℃、16 h光照/8 h黑暗的培養(yǎng)箱中培養(yǎng)。將點(diǎn)播在培養(yǎng)基上10 d的本氏煙草和轉(zhuǎn)StAN1基因的煙草幼苗移栽至甘露醇濃度為0、100、300、500、700和900 mmol/L的MS培養(yǎng)基上豎直培養(yǎng),置于25 ℃、16 h光照/8 h黑暗的培養(yǎng)箱中。
對(duì)播種45 d的煙草幼苗進(jìn)行干旱處理,隔3 d取1次煙草葉片,凍存在-80 ℃冰箱。丙二醛含量測(cè)定采用硫代巴比妥酸法[22],脯氨酸含量測(cè)定采用酸性茚三酮比色法[22],超氧化物歧化酶活性測(cè)定采用氮藍(lán)四唑(NBT)法[22];基因相對(duì)表達(dá)量的測(cè)定采用2-ΔΔCT法計(jì)算。
對(duì)播種45 d的煙草幼苗同時(shí)進(jìn)行干旱和熱處理,將煙草幼苗置于50 ℃培養(yǎng)箱中,分別于0、1、3、5、7、9和11 h時(shí)取煙草葉片,用于NBT染色和基因相對(duì)表達(dá)量測(cè)定。NBT染色的方法是用0.5 mg/mL NBT沒過所取樣本葉片,用真空泵抽濾7~10 min,將葉片放入無水乙醇中沸水浴脫色;基因相對(duì)表達(dá)量的測(cè)定采用2-ΔΔCT法計(jì)算。
1.3 統(tǒng)計(jì)學(xué)分析
用Excel 2016 對(duì)數(shù)據(jù)進(jìn)行統(tǒng)計(jì)學(xué)分析,每組均有3個(gè)以上的生物學(xué)重復(fù)。
2 結(jié)果與分析
2.1 轉(zhuǎn)基因煙草鑒定
如圖1,A所示,轉(zhuǎn)基因煙草的葉片明顯變紫。花青素含量測(cè)定結(jié)果(圖1,B)顯示,轉(zhuǎn)基因煙草葉片中的花青素含量極顯著高于野生型煙草,達(dá)到0.58 mg/g,是野生型煙草的4.83倍。
提取轉(zhuǎn)基因煙草的總DNA,PCR鑒定結(jié)果(圖1,C)顯示,檢測(cè)760 bp,與目的片段大小一致,表明StAN1基因已經(jīng)成功轉(zhuǎn)入,轉(zhuǎn)基因煙草的陽性率是82.6%。對(duì)轉(zhuǎn)基因煙草進(jìn)行qRT-PCR檢測(cè),結(jié)果(圖1,D)顯示,轉(zhuǎn)基因煙草中StAN1基因的相對(duì)表達(dá)量是本氏煙草的17倍,極顯著高于本氏煙草,充分證明了StAN1基因成功在轉(zhuǎn)基因植株中過表達(dá)。

A.轉(zhuǎn)基因煙草圖片; B.花青素含量;C.PCR鑒定轉(zhuǎn)StAN1基因煙草:1.野生型煙草(WT);2~24.轉(zhuǎn)StAN1基因煙草;D.qRT-PCR鑒定轉(zhuǎn)基因煙草StAN1基因表達(dá)量;WT.野生型煙草;StAN1ox. 過表達(dá)StAN1的轉(zhuǎn)基因煙草;**表示差異極顯著(P<0.01),*表示差異顯著(P<0.05);下同
2.2 干旱脅迫對(duì)轉(zhuǎn)基因煙草種子萌發(fā)的影響
野生型煙草和轉(zhuǎn)基因煙草種子經(jīng)過10%次氯酸鈉消毒后(圖2,A),點(diǎn)播在含不同濃度甘露醇的MS培養(yǎng)基上,每日觀察煙草的萌發(fā)情況。鑒定煙草萌發(fā)的標(biāo)準(zhǔn)是種子露白超過種子自身長(zhǎng)度的一半。
萌發(fā)結(jié)果(圖2,B、C)顯示,甘露醇濃度為0時(shí),野生型煙草和轉(zhuǎn)基因煙草的萌發(fā)情況一致,在點(diǎn)播5 d后萌發(fā)率均可達(dá)到96%以上。甘露醇濃度為50和100 mmol/L時(shí),兩者的萌發(fā)情況沒有明顯差異。甘露醇濃度為150 mmol/L時(shí),兩者的萌發(fā)率出現(xiàn)了明顯的變化,轉(zhuǎn)StAN1基因煙草在第7天時(shí)的萌發(fā)率達(dá)到了52%,是野生型煙草萌發(fā)率的2.01倍,顯著高于野生型煙草萌發(fā)率。
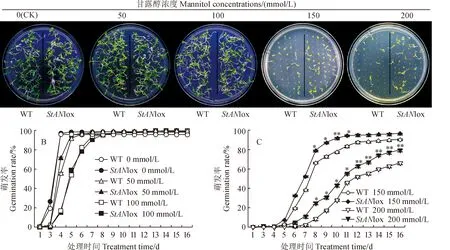
A.煙草種子點(diǎn)播在培養(yǎng)基上第7天時(shí)萌發(fā)情況;B.0、50、100 mmol/L甘露醇濃度下的煙草種子萌發(fā)率;C. 150、200 mmol/L甘露醇濃度下的煙草種子萌發(fā)率
2.3 干旱脅迫對(duì)轉(zhuǎn)基因煙草幼苗根長(zhǎng)的影響
將萌發(fā)10 d后的煙草幼苗移栽至不同濃度甘露醇的MS培養(yǎng)基上豎直培養(yǎng)(圖3),觀察根的生長(zhǎng)情況。結(jié)果表明,野生型煙草和StAN1轉(zhuǎn)基因煙草根的生長(zhǎng)都會(huì)受到干旱脅迫的影響;在甘露醇濃度為0和100 mmol/L培養(yǎng)基上,轉(zhuǎn)基因煙草的根長(zhǎng)分別達(dá)到1.9和1.7 cm,分別是野生型煙草的1.46和1.30倍,比野生型煙草根長(zhǎng)顯著增加;而當(dāng)甘露醇濃度上升至300、500、700和900 mmol/L時(shí),兩者之間沒有顯著性差異。

圖3 不同濃度甘露醇下煙草根的生長(zhǎng)情況Fig.3 Tobacco root growth under different mannitol concentrations
2.4 干旱脅迫對(duì)轉(zhuǎn)基因煙草生理生化指標(biāo)的影響
如圖4,A所示,對(duì)野生型煙草和轉(zhuǎn)基因煙草同時(shí)進(jìn)行干旱處理,每隔3 d取樣1次,進(jìn)行生理指標(biāo)測(cè)定。在未進(jìn)行干旱處理時(shí)(圖4,B),兩者的丙二醛(MDA)含量相當(dāng),沒有顯著性差異;在第3天時(shí),干旱脅迫程度較輕,MDA含量變化差異不大;隨著干旱脅迫程度的加深,野生型煙草和轉(zhuǎn)基因煙草中的MDA含量均有不同程度的增加。在干旱脅迫9 d時(shí),野生型煙草葉片中的MDA含量達(dá)到4.05 μmol/g,是轉(zhuǎn)基因煙草葉片MDA含量的1.14倍,顯著高于轉(zhuǎn)基因煙草;在干旱脅迫12和15 d時(shí),野生型煙草葉片中的MDA含量分別是轉(zhuǎn)基因煙草的1.29和1.38倍,極顯著高于轉(zhuǎn)基因煙草。研究表明干旱脅迫下野生型煙草葉片中的MDA含量總體高于轉(zhuǎn)基因煙草,表明野生型煙草葉片在干旱脅迫下膜脂過氧化程度相對(duì)于轉(zhuǎn)基因煙草更嚴(yán)重,尤其在干旱脅迫后期,野生型煙草葉片受干旱影響更大。

圖4 干旱脅迫下轉(zhuǎn)StAN1基因煙草丙二醛含量、超氧化物歧化酶活性以及脯氨酸含量變化Fig.4 Changes of malondialdehyde content,superoxide dismutase activity and proline content in StAN1 transgenic tobacco under drought stress
同時(shí),干旱處理后測(cè)定煙草植株葉片中超氧化物歧化酶(SOD)的活性,結(jié)果如圖4,C所示。在沒有受到干旱脅迫時(shí),野生型煙草和轉(zhuǎn)基因煙草葉片中的SOD活性沒有顯著性差異;在受到干旱脅迫后,野生型煙草葉片中的SOD活性有緩慢上升,而轉(zhuǎn)基因煙草SOD活性出現(xiàn)了明顯上升。在干旱處理6 d時(shí),轉(zhuǎn)基因煙草SOD活性達(dá)到了83 U/g,是野生型煙草的2.13倍;干旱處理9 d時(shí),轉(zhuǎn)基因煙草的SOD活性是野生型煙草的1.61倍,在干旱處理6~9 d內(nèi)轉(zhuǎn)基因煙草的SOD活性均極顯著高于野生型煙草。隨著干旱脅迫程度的加深,葉片水分散失,SOD活性也隨之降低,不能有效消除活性氧,葉片出現(xiàn)了萎蔫、壞死等狀況。
另外,對(duì)野生型和轉(zhuǎn)基因煙草進(jìn)行干旱處理后,測(cè)定煙草葉片中的脯氨酸含量,結(jié)果如圖4,D所示。在剛開始受到干旱脅迫時(shí),兩者葉片中的脯氨酸含量沒有顯著性差異,隨著干旱脅迫程度的加深,兩者葉片中脯氨酸含量呈顯著性差異。在干旱處理6 d時(shí),轉(zhuǎn)基因煙草葉片中的脯氨酸含量顯著高于野生型煙草;干旱處理9 d時(shí),轉(zhuǎn)基因煙草的脯氨酸含量達(dá)到了野生型煙草的2.06倍;干旱處理15 d時(shí),轉(zhuǎn)基因煙草葉片中的脯氨酸含量達(dá)到117 μg/g,極顯著高于野生型煙草(89 μg/g)。研究表明,在應(yīng)對(duì)干旱脅迫時(shí),轉(zhuǎn)基因煙草能夠很快地提高葉片中脯氨酸含量,從而增加葉片保水的能力,維持細(xì)胞滲透壓,使光合作用等生命活動(dòng)正常進(jìn)行,保持細(xì)胞穩(wěn)態(tài)。
2.5 干旱脅迫下ERF和LEA基因表達(dá)量的變化
分別在干旱脅迫1、3、5、7和9 d選取野生型煙草和轉(zhuǎn)基因煙草葉片,提取總RNA并反轉(zhuǎn)錄成cDNA, qRT-PCR鑒定相關(guān)抗旱基因的相對(duì)表達(dá)量。如圖5所示,ERF基因的相對(duì)表達(dá)量在干旱處理的第7、9、11天時(shí),是野生型煙草的8、66和3倍,極顯著高于野生型煙草的表達(dá)量;LEA基因的相對(duì)表達(dá)量在第5、7、9天時(shí),分別達(dá)到了野生型的4、3和3倍,極顯著高于野生型煙草的表達(dá)量。
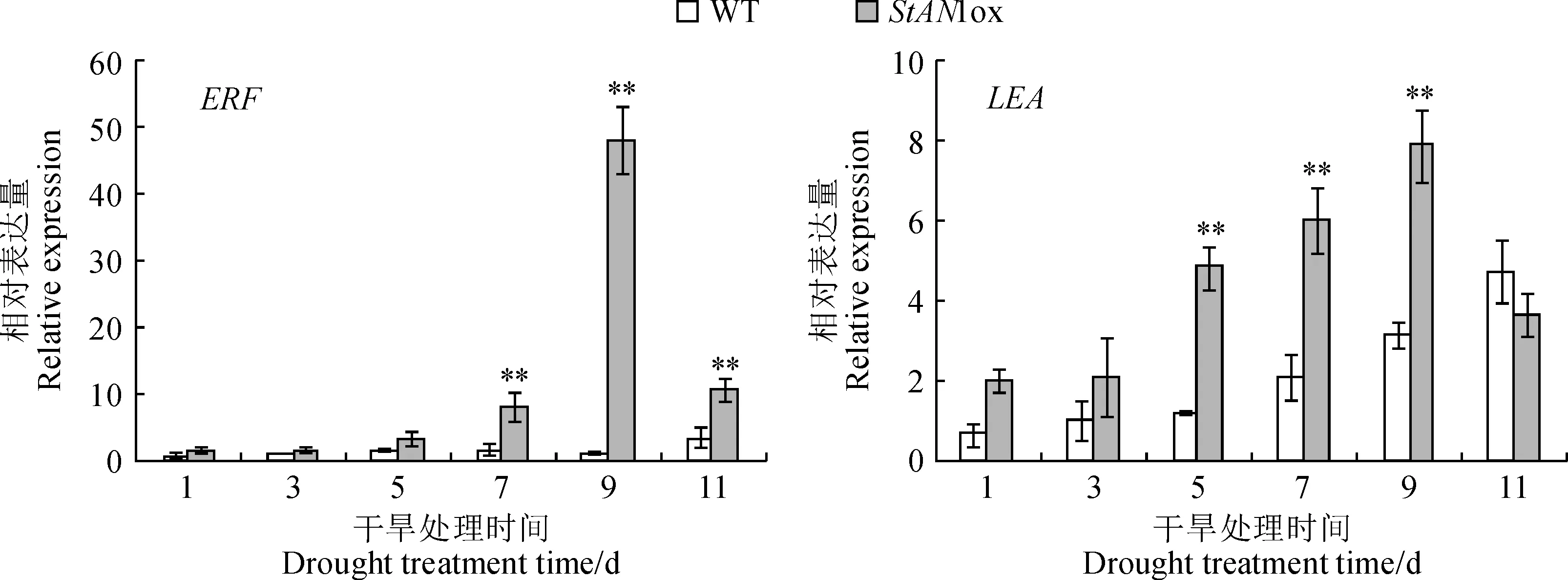
圖5 生長(zhǎng)45 d的煙草幼苗干旱處理后基因相對(duì)表達(dá)量的變化 Fig.5 Relative expression of resistance gene in 45-day-old tobacco seedlings under drought treatments
2.6 旱熱脅迫下NBT染色以及基因表達(dá)量的變化
對(duì)野生型煙草和轉(zhuǎn)基因煙草同時(shí)進(jìn)行干旱和50 ℃熱處理,分別在0、1、3、5、7、9和11 h時(shí)取樣,進(jìn)行NBT染色和實(shí)時(shí)熒光定量PCR。結(jié)果(圖6,A)顯示,野生型煙草進(jìn)行染色后的顏色明顯比轉(zhuǎn)基因煙草的葉片顏色深,說明野生型煙草在受到旱熱脅迫時(shí),葉片中有更多的過氧化物與NBT反應(yīng),產(chǎn)生了藍(lán)色甲腙;而轉(zhuǎn)基因煙草葉片中的活性氧含量低,產(chǎn)生的甲腙較少,藍(lán)色較淺(圖6,B)。

A.野生型煙草;B.轉(zhuǎn)基因煙草
qRT-PCR鑒定相關(guān)抗逆基因的相對(duì)表達(dá)量結(jié)果(圖7)顯示,旱熱處理時(shí),LEA和ERF基因的相對(duì)表達(dá)量相對(duì)于干旱處理時(shí)都出現(xiàn)了顯著的增加;在旱熱處理的第3小時(shí),LEA和ERF基因的相對(duì)表達(dá)量均極顯著的高于野生型煙草,分別是野生型煙草的20.9和2.25倍。

圖7 生長(zhǎng)45 d的煙草幼苗旱熱處理后基因相對(duì)表達(dá)量的變化Fig.7 Relative expression of resistance gene in 45-day-old tobacco seedlings under drought heat treatments
3 討 論
生物體內(nèi)的花青素合成主要受到結(jié)構(gòu)基因和轉(zhuǎn)錄因子的調(diào)節(jié)。結(jié)構(gòu)基因主要控制合成苯丙素途徑中的一些關(guān)鍵酶,苯丙素經(jīng)過PAL、CHS、CHI、F3H和DFR等酶的催化反應(yīng),最終合成花青素;此外,MYB、bHLH和WD40這3類轉(zhuǎn)錄因子在花青素合成的過程中也起著關(guān)鍵作用,可以形成MBW蛋白復(fù)合體調(diào)控基因的表達(dá)[23],進(jìn)一步調(diào)控花青素的合成。StAN1轉(zhuǎn)錄因子是MYB轉(zhuǎn)錄因子中的一個(gè),能夠有效促進(jìn)花青素的合成與積累。當(dāng)植物響應(yīng)生物或非生物脅迫時(shí),花青素是一種有效的保護(hù)物質(zhì),以減輕植物所受到的傷害。在自然條件下,花青素多與糖苷結(jié)合以花色苷的形式存在,常見的有天竺葵素、矢車菊素和花翠素等[23]。
在受到干旱脅迫之后,轉(zhuǎn)基因煙草由于花青素含量高,其較強(qiáng)的活性氧清除能力是由花青素化學(xué)結(jié)構(gòu)中的羥基和酰基等這些化學(xué)基團(tuán)決定[24],從而保護(hù)植物組織以及生命活動(dòng)正常進(jìn)行,使得植物具有一定的耐旱性。野生型煙草花青素含量較低,不能及時(shí)清除多余的活性氧,植株受到的損傷嚴(yán)重。
LEA蛋白的表達(dá)量與植物耐受非生物脅迫有著密切的調(diào)控關(guān)系,LEA的啟動(dòng)子序列除了含有核心的啟動(dòng)子元件外,還有順式元件MYB的結(jié)合位點(diǎn)[24-25],同時(shí)也有研究表明MYB類轉(zhuǎn)錄因子在植物應(yīng)對(duì)非生物脅迫時(shí)能夠響應(yīng)更大范圍的下游調(diào)控[26]。因此,轉(zhuǎn)基因煙草中,StAN1基因的表達(dá),可能會(huì)上調(diào)表達(dá)MYB家族的基因,從而結(jié)合在LEA基因的啟動(dòng)子序列,上調(diào)表達(dá)LEA蛋白,增強(qiáng)轉(zhuǎn)基因煙草的耐熱耐旱性。ERF類轉(zhuǎn)錄因子參與植物種子萌發(fā)以及非生物脅迫的調(diào)控[27],還廣泛參與植物的次生代謝。干旱可以誘導(dǎo)小麥中ERF基因的轉(zhuǎn)錄[28],轉(zhuǎn)ERF基因的擬南芥以及煙草對(duì)非生物脅迫的耐受性有所提高,植物自身次生代謝產(chǎn)物的積累也能夠有效地調(diào)控植物應(yīng)對(duì)生物或者非生物脅迫。已有研究表明,ERF能夠有效地促進(jìn)木質(zhì)素的積累,而木質(zhì)素的上游合成途徑是從苯丙素合成途徑開始,這也是花青素合成的上游途徑之一,因此,ERF調(diào)控木質(zhì)素合成的同時(shí),也可能間接提高花青素的合成,從而提高植物耐熱耐旱性。而且,也有研究表明,ERF會(huì)與一些MYB家族的轉(zhuǎn)錄因子結(jié)合,形成轉(zhuǎn)錄因子復(fù)合物調(diào)控次級(jí)代謝產(chǎn)物的合成[29]。
綜上所述,本試驗(yàn)通過干旱處理不同時(shí)期的轉(zhuǎn)基因煙草,并且測(cè)定丙二醛和脯氨酸等生理指標(biāo),發(fā)現(xiàn)StAN1在提高轉(zhuǎn)基因煙草花青素含量的同時(shí),在植物應(yīng)對(duì)干旱和熱脅迫時(shí)也有一定的作用。干旱和旱熱處理時(shí),轉(zhuǎn)基因煙草中LEA和ERF基因的表達(dá)均顯著高于野生型煙草,一定程度上提高了轉(zhuǎn)基因煙草的耐熱耐旱性。本研究結(jié)果為進(jìn)一步深入研究StAN1基因功能奠定了堅(jiān)實(shí)的基礎(chǔ)。
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
中國(guó)民間療法(2021年5期)2021-06-09 09:21:26
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
浙江中西醫(yī)結(jié)合雜志(2017年2期)2017-01-12 18:23:59
當(dāng)代化工研究(2016年9期)2016-03-20 16:22:08
食藥用菌(2016年6期)2016-03-01 03:24:27
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國(guó)藥業(yè)(2014年24期)2014-05-26 09:00:09
