10份桑種質資源親緣關系鑒定
2021-03-25 07:51:48任迎虹祁偉亮
中國野生植物資源 2021年2期
關鍵詞:資源
任迎虹,祁偉亮,陳 潔
(成都師范學院 化學與生命科學學院,四川 成都 611130)
桑樹(Mulberry)屬于重要的經濟植物[1],葉片互生、葉緣有鋸齒、花雌雄同株或異株。桑科共約53屬,1400余種,我國是目前世界上桑樹資源分布最多的國家。最初的植物學分類依據主要是形態學,如瑞典植物分類學家林奈將桑屬植物分為7個種[2]。1842年Moretti將桑屬分為10個種[3]。1873年法國植物分類學家Bureau根據桑葉和雌花特征進一步劃分為19個變種以及12個亞變種[4]。后期夏明炯在1991年發表的《桑樹分類概述》一文中指出我國桑樹分為15個桑種和4個變種,是世界上桑種分布最多的國家[5]。桑樹作為家蠶的飼料經過了上千年的栽培和雜交,在形態上發生了很多變異,形成了許多種和變種,因此單純依靠形態學依據的分類顯然可信性不高。
隨著生物技術的不斷發展,分類學家開始在分子水平上尋找分類依據。在被子植物基因組中,ITS(Internal trnscribed spacer)基因序列具有長度變異小、非常保守等特性,適合用于低分類群的系統發育分析。史全良和趙衛國[6]以蒙桑(M.mongolica)為材料采用PCR產物直接測序法,測定了核糖體基因轉錄內間隔區序列,指出ITS序列在桑樹分子系統學研究的潛在重要性。2012年,Nepal和Ferguson[7]采用ITS和trnL-trnF將全球的桑屬分為13個種,并強調桑屬可能不是單源進化,分為北美和亞洲兩個亞屬。
本研究旨在對10份收集的桑樹資源進行ITS序列分析,結合形態學調查和相關指標,從分子學和形態學角度闡明10個野外收集的桑樹資源和已知桑屬的親緣關系,這對于桑種質的指紋鑒別、親緣關系分析、育種親本選擇以及桑的分子育種等技術問題均具有重要意義。
1 材料與方法
1.1 材料
本試驗材料主要收集于涼山州各主要蠶種場。由于地方蠶桑種植戶對于引進的品種或者部分栽培品種信息不能準確描述,故將收集的10份桑種質資源以嫁接苗的形式,繁種于成都師范學院桑樹資源圃,以編號形式暫命名為1,R-2016JLJ-1;2,R-2016HYB1-2;3,R-2016XYJL-2;4,湖桑32(已知品種);5,R-2016HYB2-2;6,R-2016KJ-1;7,R-2016LH-2;8,R-2016BL-2;9,R-2016SM-3;10,R-2016CS2-2。
1.2 DNA提取及PCR產物擴增
每個樣品取2~3片嫩葉,采用CTAB法[8]提取桑樹基因組DNA。PCR擴增20 uL反應體系中,包含Tag mix(10 uL)、ddH2O(7 uL)、DNA(1 uL)、引物ITS-F(ITS 5)(5'- GGAAGTAA AAGTCGTA ACAAGG-3')1 uL、引物ITS-R(ITS 4)(5'-TCCTC CGCTTATTG ATATGC-3')1 uL(購自上海生工生物工程有限公司)。
擴增程序:94℃預變性5 min,94℃變性30 s,55℃退火30 s,72℃延伸1 min,共38個循環,再72℃保溫延伸10 min,12℃條件下保存。PCR反應產物經1%瓊脂糖電泳,電壓為120 V(穩壓),電泳約10 min,溴化乙錠(Ethidium bromide,EB)染色5 min,在凝膠成像系統拍照記錄[9]。
1.3 序列數據處理
測序結果用SeqMan5.01軟件轉換,對少數誤判堿基,根據堿基峰形更正。用MEGA7.0.14軟件進行裁剪比對,除去兩端非ITS序列部分。根據GenBank Blast檢索得到親緣關系相近的白桑(M.alba,XNS0052)、魯桑(M.multicaulis,AM042003)、雞桑(M.australis,XNSK0024)和山桑(M.bombycis,XNS0277)4份桑屬ITS序列,與本試驗的10份ITS序列一起,利用MEGA7.0.14軟件進行序列長度、G + C含量對比分析,單獨分析變異位點和計算遺傳距離。根據GenBank上檢索到的暹羅桑(M.rotundiloba,AY345150)、蒙桑(M.mongolica,AY345158)、貴14號長穗桑(M.wittiorum,AY345155)、北美默里桑(M.murrayana,FJ605515)、新疆黑桑(M.nigra,XNS0218)5份桑屬ITS序列和Blast得到的4份ITS序列與本試驗桑種質材料ITS序列一起,構樹(Broussonetiapapyrifera,XNS0377)、冠毛榕(Ficusgasparriniana,JQ773883)和柘樹(Macluratricuspidata,XNS0007)作為外類群(Outgroup),通過MEGA7.0.14軟件采用最大簡約法(Maximum)分析,構建系統發育樹。
1.4 形態特征調查
本次共調查10份桑樹資源的農藝形狀,參照《桑樹種質資源描述規范和數據標準》[10],包括描述性狀,如葉形、側枝、葉色、葉尖、葉緣等,以及數量性狀,如葉寬、葉長、葉柄長、葉柄寬等,每個植株隨機抽樣9根枝條并利用數據處理軟件Excel進行數據統計。最后結合ITS數據分析結果,并參考中國高等植物圖鑒[11]、中國桑樹品種志[12]、中國數字植物標本館(中國植物智 http://www.iplant.cn/)進行資源鑒定。
2 結果與分析
2.1 ITS序列長度及堿基比例
對10份材料進行凝膠電泳,圖像大致在700 bp左右(圖1),根據來自Genbank的魯桑種(AM042003)的18S rRNA基因3′端、26S rRNA基因5′端、5.8S rDNA基因堿基序列,確定本試驗各材料的ITS1、5.8S、ITS2序列范圍[13-15]。
圖1 引物ITS4/ITS5對10份桑樹種質材料ITS 基因組DNA的PCR擴增產物電泳譜圖Fig. 1 ITS4/ITS5 10 parts of mulberry germplasm material ITS Genomic DNA PCR products electrophoresis spectrum
軟件分析顯示,桑樹ITS序列(包含5.8S rDNA)全長為576~578 bp,G+C含量為59.17%~59.97%(見表1),其中5.8S rDNA編碼區極為保守,在所有測得以及對照的序列中,長度均為100 bp,且G+C含量都為48.00%;而ITS1和ITS2序列表現出差異性,長度分別為174~175 bp和302~303 bp,G+C含量分別為60.00%~62.07%和62.38%~62.71%。其中R-2016SM-3的ITS1、ITS2的長度分別為174 bp、303 bp,G+C的含量分別為62.07%、62.71%,與雞桑一致,推測R-2016SM-3可能屬于雞桑(M.australis)。此外,R-2016HYB2-2、R-2016LH-2、R-2016BL-2和R-2016CS2-2都存在不同程度的變異。剩下的R-2016JLJ-1、R-2016HYB1-2、R-2016XYJL-2、湖桑32以及R-2016KJ-1的ITS序列長度及G+C含量與白桑相同,推測他們可能屬于白桑(M.alba)。
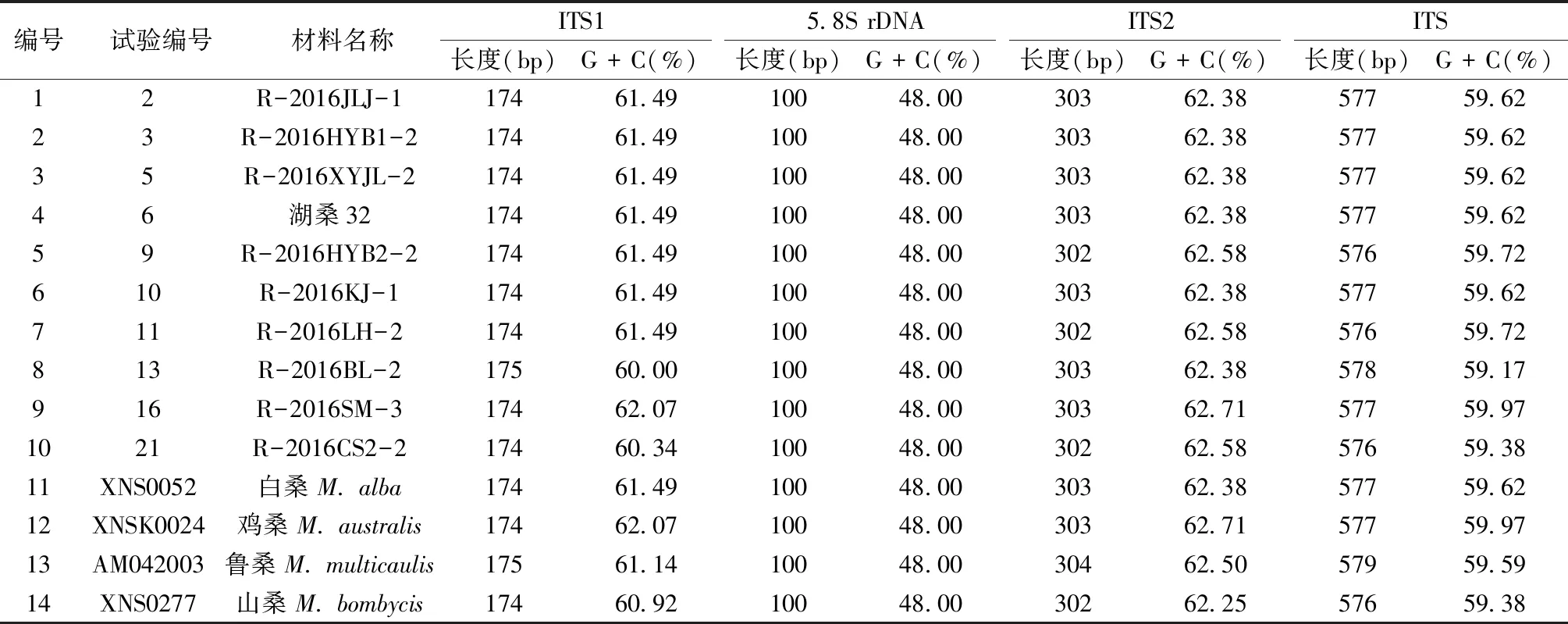
表1 桑屬種質資源ITS序列長度、G + C含量變異Table 1 The length of ITS sequences and contents of G + C in Morus
2.2 ITS序列遺傳距離及變異位點分析
利用MEGA7.0.14軟件采用Kimura 2-parameter model方法分析了10份桑種質材料的ITS序列的遺傳距離(Genetic distance)(表2)。10份桑種質材料的遺傳距離為0.000~0.102。R-2016JLJ-1、R-2016XYJL-2、湖桑32、R-2016HYB2-2、R-2016KJ-1和R-2016LH-2的遺傳距離最近(0.000),表明其親緣關系最近;R-2016HYB1-2與R-2016BL-2的遺傳距離最遠(0.102),表明其親緣關系最遠。
通過ITS序列堿基—長度、G+C的含量變化及遺傳距離分析,排除相似性高的材料,本研究單獨對R-2016HYB1-2、R-2016HYB2-2、R-2016LH-2、R-2016BL-2、R-2016SM-3以及R-2016CS2-2進行ITS序列變異位點(Variable sites)對比分析(表3)。采用王潔等[16]的方法,可以發現這6個材料共有60個變異位點,變異位點集中在ITS1與ITS2區,其中ITS1區最多,R-2016BL-2在ITS1區域變異位點最多(50個),變異類型有A-C、A-G、A-T、T-G、C-T、G-C等。ITS1區有52個變異位點,其中簡約信息位點(Parsimony informative site)占21個,分別占ITS1序列平均長度(175 bp)的29.71%和12.00%。ITS2區有8個變異位點,其中簡約信息位點占1個,分別占ITS2序列平均長度(303 bp)的2.31%和0.33%,表明ITS1序列所包含的信息含量明顯高于ITS2序列。

表2 15份桑屬ITS序列遺傳距離及相似性Table 2 Genetic distances and percent identities of 15 mulberry germplasm materials based on ITS
2.3 ITS序列親緣關系分析
加上GenBank上的白桑(M.alba,XNS0052)等9份外源桑屬材料,用構樹(B.papyrifera,XNS0377)等3份材料作為外類群,最大簡約(MP)法聚類分析結果顯示,分支圖顯示樹長為378,一致性指數(Consistency index,CI)為0.8862,趨同性指數(Homoplasy index,HI)= 0.1138,保持性指數(Retention index,RI)為0.7529,調整后一致性指數(Rescaled consistency index,RC)為0.6672。對比序列共計580個位點(含Gap),其中有517個不變位點,38個變異非信息位點,22個簡約信息位點。

表3 ITS基因序列比較Table 3 Comparison of ITS gene sequences

續表3 ITS基因序列比較Table 3 Comparison of ITS gene sequences
從聚類結果來看,系統發育樹首先將柘樹、冠毛榕分出,自舉檢驗(Bootstrap)支持率為99%。可以明顯看出,在桑科植物的構屬、柘屬和榕屬中,構屬和桑屬親緣關系最近。R-2016SM-3與雞桑單獨聚在一起,其自舉檢驗支持率為77%,推測R-2016SM-3屬于雞桑(M.australis)。R-2016BL-2和R-2016CS2-2聚在一起,與蒙桑的自檢支持率為96%,推測R-2016BL-2和R-2016CS2-2屬于蒙桑(M.mongolica),為蒙桑的一個變種。R-2016HYB1-2與暹羅桑單獨聚為一小支,自檢支持率為86%,推測R-2016HYB1-2屬于暹羅桑(M.rotundiloba)。剩下的R-2016HYB2-2、R-2016KJ-1、R-2016JLJ-1、湖桑32、R-2016LH-2和R-2016XYJL-2與白桑和魯桑聚在一起,由于魯桑也是白桑的一個變種,表明它們屬于白桑(M.alba)(圖2)。
3 結論
中國是桑屬植物集中分布和分化中心,又是桑屬植物的遺傳多樣性中心,許多桑種起源于我國,如蒙桑(M.mongolica),雞桑(M.australis),滇桑(M.yunnanensts),白桑(M.alba),山桑(M.bombycis)等。桑屬由林奈[2]建立之后,先后有許多學者在分子學方面對桑屬的系統分類進行了研究[17-20]。陳仁芳[21]采用MP法對桑ITS序列進行聚類分析,結果表明桑樹進化的順序依次為:新疆黑桑、北美默里桑→白桑、暹羅桑(AM042005)、瑞穗桑、廣東桑、雞桑、魯桑(AM041999)、蒙桑、山桑→華桑、奶桑、川桑→利川長穗桑、咸豐長穗桑;73份桑資源只有33份材料有不同程度的變異,同源性高達93.1%~100%。本研究ITS進化分析結果,首先將桑樹近緣屬植物柘樹、冠毛榕和構樹分開,同時將黑桑、北美默里桑獨立出一支,這一結果與陳仁芳研究結果吻合,表明試驗分析結果可靠。吳征鎰等[22]研究報道,白桑(M.alba)在早期系統演化上是最大的一個干支,也是現存數量最多一類桑種,其中魯桑(M.multicaulis)屬于白桑的變種[23]。本研究中R-2016HYB2-2、R-2016KJ-1、R-2016 JLJ-1、湖桑32、R-2016LH-2和R-2016XYJL-2與白桑和魯桑聚在一起,10份桑樹資源多以白桑為主,這也證明了吳征鎰的觀點,同時鑒定出1份雞桑、2份蒙桑和1份暹羅桑。
為了更準確的評價系統歸類結果的可靠性,本研究參考《中國植物志》等資源對收集的10個桑樹資源進行了形態特征及相關數量結果統計(表4),結果表明10個桑樹資源的形態特征存在差異性和相似性。R-2016BL-2和R-2016CS2-2與蒙桑(M.mongolica)親緣關系較近,且R-2016BL-2(圖3g)和R-2016CS2-2(圖3h)的葉形為裂葉形、表面光滑,葉長分別為9.20 cm、12.90 cm,葉寬分別為8.80 cm、10.20 cm,葉柄長分別為2.43 cm、2.63 cm,尖端尾尖,葉緣單鋸齒形,形態學特征[11,24]與裂葉蒙桑的相似度高(圖3m),該結果表明R-2016BL-2和R-2016CS2-2屬于蒙桑(M.mongolica)。R-2016SM-3(圖3j)與雞桑(圖3l)單獨聚在一起,且葉形態特征符合雞桑(M.australis)的生物學特征描述[25],因此鑒定R-2016SM-3為雞桑。據《中國桑樹品種志》介紹,四川在1982年桑樹品種譜查中還發現蜀雅桑、雅中里、插桑、雅周桑、蜀名桑等扦插易生根的雞桑[12],這也證實四川地區的確存在雞桑。
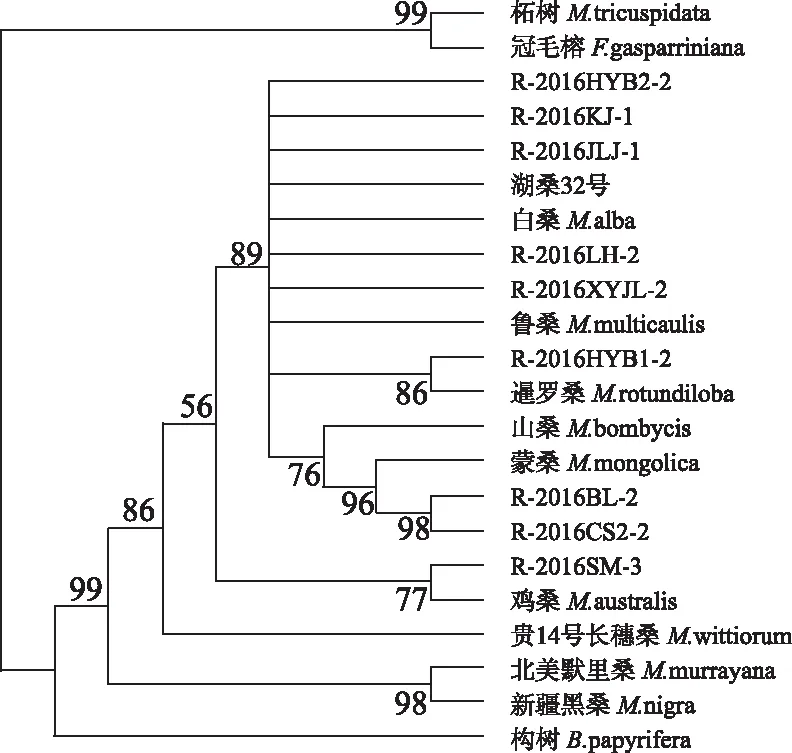
圖2 22份桑種質材料基于ITS的系統進化樹Fig. 2 A phylogenetic tree of 22 mulberry germplasm materials based on ITS
白桑不僅分布廣泛、且遺傳背景變異多,這也形成很多變種,如魯桑(M.multicaulis-俗名女桑、湖桑、白桑)、泰國的暹羅桑(M.rotunbiloba)等[21,26]。R-2016HYB2-2(圖3a)、R-2016KJ-1(圖3b)、R-2016JLJ-1(圖3c)、湖桑32(圖3d)、R-2016LH-2(圖3e)和R-2016XYJL-2(圖3f)的葉長在9.60~14.55 cm,葉寬在6.50~10.00 cm,葉柄長在2.43~4.77 cm,葉均為卵形、表面光滑、鮮綠色,葉形態特征符合魯桑的生物學特征描述(圖3k)。因此,鑒定R-2016HYB2-2、R-2016KJ-1、R-2016JLJ-1、湖桑32、R-2016LH-2和R-2016XYJL-2屬于白桑類。R-2016HYB1-2與暹羅桑單獨聚為一小支,該品種應該屬于國外引進品種。
4 討論
很多科研工作者對桑屬的分類和系統進化工作做了大量的研究。1995年,向仲懷首次采用RAPD技術構建了9個桑品種的DNA指紋圖譜[17]。隨后,許多子標記技術如ISSR、SSR和ITS等相繼應用到桑樹系統學分類研究工作中[27]。如Vijayan等[28]利用形態解剖和ISSR 分子標記技術鑒定了17種野生桑樹資源的遺傳多樣性。2006年,印度人完成了首個桑樹品種的葉綠體全基因組測序[29],同年,Venkateswarlu等[30]利用ISSR、RAPD和SSR 三種分子標記繪制了印度桑K2的遺傳圖譜,該研究為桑樹近緣種屬分類提供了重要的參考。2012年,Nepal首次采用ITS和trnL-trnF將全球的桑屬分為13個種,研究發現桑可能不是單源進化,分為北美和亞洲兩個亞屬[7]。曾其偉等[31]利用ITS序列對桑屬做了較為系統的分析,提出白桑是最大的一種且種質資源多,但因白桑資源形態多樣,將他們歸為一個大種存在爭議[32]。在本研究中,也發現超過一半的桑品種屬于白桑。因此,下一步將采用ITS與trnL-trnF進行多片段聯合分析,同時綜合細胞學研究、解剖學等[33],進行桑樹品種的親緣關系分析,并為桑樹的資源選擇和雜交育種提供依據。

圖3 10份種質資源的形態圖和標本圖Fig. 3 Morphological maps and specimen figures of 10 mulberry 注:a,R-2016HYB2-2;b,R-2016KJ-1;c,R-2016JLJ-1;d,湖桑32;e,R-2016LH-2;f,R-2016XYJL-2;g,R-2016BL-2;h,R-2016CS2-2;i,R-2016HYB1-2;j,R-2016SM-3;k,魯桑標本,中國植物智條碼SZG0Q005128;l,雞桑標本,中國植物智條碼CSH0090591;m,蒙桑標本,中國植物智條碼QFNU0018312
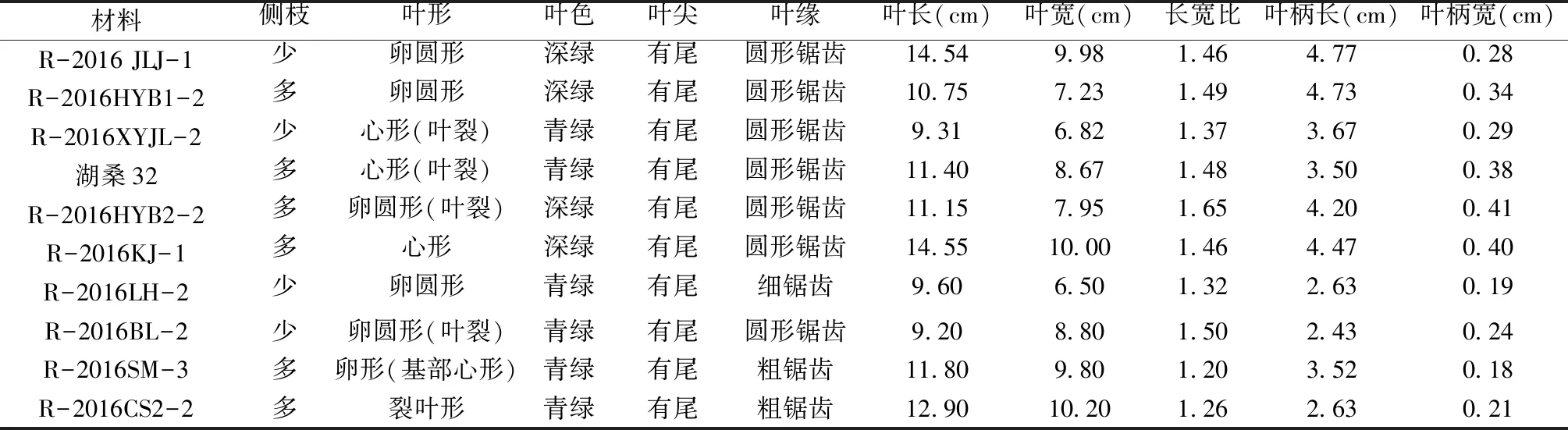
表4 10份桑樹種質資源形態數據調查Table 4 Survey of morphological data of 10 mulberry germplasm resources
猜你喜歡
江蘇安全生產(2023年1期)2023-02-08 05:58:38
資源節約與環保(2022年8期)2022-09-20 02:25:22
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
藝術品鑒(2020年7期)2020-09-11 08:04:44
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
當代貴州(2018年28期)2018-09-19 06:39:04
資源再生(2017年3期)2017-06-01 12:20:59
決策(2015年9期)2015-09-10 07:22:44
