衛矛科溝瓣屬與衛矛屬的數量分類學研究
2021-03-24 11:48:59劉慧圓孟世勇劉全儒
廣西植物 2021年1期
劉慧圓 孟世勇 劉全儒
摘 要: 衛矛屬(Euonymus L.)和溝瓣屬(Glyptopetalum Thw.)的系統關系長期以來備受爭議,為理清它們之間的關系,該研究選擇溝瓣屬12種、衛矛屬17種和假衛矛屬(Microtropis Wall. ex Meisn.) 2種,共計31個分類單元(OTUs),測量并統計了56個形態性狀,同時利用聚類分析(UPGMA)和主坐標分析(PCoA)的方法進行了數量分類學研究。結果表明:溝瓣屬與衛矛屬具有較高的表征相似性,二者之間存在的過渡類群有縉云衛矛(Euonymus chloranthoides Yang)和淡綠葉衛矛(E. pallidifolium Hayata)以及海南溝瓣 [Glyptopetalum fengii (Chun & F. C. How) Ding Hou]、白樹溝瓣 [G. geloniifolium (Chun & F. C. How) C. Y. Cheng]。該研究認為將溝瓣屬作為衛矛屬的一個組,即溝瓣組(Sect. Glyptopetalum),放在冬青衛矛組(Sect. Ilicifolia) 附近更為合理。對性狀的主坐標排序分析顯示,葉片長度、葉片寬度、葉側脈數目、花瓣數目、心室數目、每室胚珠數、花序分枝數、是否具有假種皮、蒴果是否開裂、花期與果序梗長度在衛矛屬分類中具有重要的價值。
關鍵詞: 衛矛屬, 溝瓣屬, 形態性狀, 數量分類學, 聚類分析, 主坐標分析
Abstract: The systematic relationship between the genera Euonymus L. and Glyptopetalum Thw. has been controversial for a long time. In order to clarify it, we selected 31 operational taxonomic units (OTUs) (12 from Glyptopetalum, 17 from Euonymus, and 2 from Microtropis Wall. ex Meisn.), measured 56 morphological characters of them, and performed cluster analysis (unweighted pair group method using arithmetic average) as well as principal coordinate analysis. The results were as follows: Glyptopetalum and Euonymus had a high morphological similarity, between which were some transition groups, e. g. Euonymus chloranthoides Yang, E. pallidifolius Hayata, Glyptopetalum fengii (Chun et F. C. How) Ding Hou, and G. geloniifolium (Chun et F. C. How) C. Y. Cheng. The study suggested that it is more reasonable to place Glyptopetalum as Euonymus Sect. Glyptopetalum, near Euonymus Sect. Ilicifolia. The principal coordinate analysis (PCoA) showed the great taxonomic value of some characters in Euonymus, including leaf blade length and width, number of lateral veins, petals, locules, ovules per locule, inflorescence branches, presence/absence of aril, dehiscence of capsule, flowering time, and infructescence stalk length.
Key words: Euonymus L., Glyptopetalum Thw., morphological characters, quantitative taxonomy, cluster analysis (UPGMA), principal coordinate analysis(PCoA)
衛矛屬隸屬于衛矛科(Celastraceae)衛矛族(Euonymeae),包括傳統上以蒴果形態為主要劃分依據而建立的5個組,即翅果衛矛組(Sect. Uniloculares)、深裂衛矛組(Sect. Melanocarya)、刺果組(Sect. Echinococcus)、冬青衛矛組(Sect. Ilicifolii)和衛矛組(Sect. Euonymus)。該屬約129種,模式種是歐衛矛(Euonymus europaeus L.),分布于東西兩半球的亞熱帶和暖溫帶地區,我國西南地區是其現代分化與多樣性中心(誠靜容等, 1999; Ma, 2001)。溝瓣屬(Glyptopetalum Thw.)因具有4數花、子房每室具有1個倒生胚珠等特征而從衛矛屬中獨立出來(Thwaites, 1856),模式種為錫蘭溝瓣(Glyptopetalum zeylanicum Thw.),該屬約有40種,分布于亞洲熱帶及亞熱帶,我國有10~13種,分布于廣西、云南、海南和臺灣等熱帶或亞熱帶地區,是溝瓣屬分布的北緣(Hou, 1963; 誠靜容等, 1999; Meng et al., 2012)。然而,溝瓣屬的系統地位一直存在爭議。一些學者根據溝瓣屬具有4數花、每個心皮僅有1枚胚珠、種脊具有3~7條分枝等多處生殖結構特征,堅持將其作為一個獨立的屬處理(Prain, 1891; Hou, 1962; 誠靜容等, 1999; Meng et al., 2012; Simmons et al., 2012; Savinov, 2014);而有的學者卻認為衛矛屬變異范圍較大,溝瓣屬的特征應屬于衛矛屬的變異范圍(Kurz, 1877; Baillon, 1880)。Simmons et al. (2012)結合形態學和葉綠體基因(matK, trnL-F)、核基因(ITS 和26S rDNA)對衛矛族(Euonymeae)進行了系統發育分析,結果顯示溝瓣屬與衛矛屬植物共同組成一個單系類群。Li et al.(2014)對衛矛屬、溝瓣屬等48個物種,利用2段核基因序列(ITS和 ETS)和3段葉綠體基因 (psbA-trnH、rp136-infA-rps8和trnC-ycf6)做了最大似然法(MP)和貝葉斯分析(BI),研究結果顯示溝瓣屬與衛矛屬共同組成一個單系類群。然而,前人主要是基于少數(2個或3個)代表性的溝瓣屬物種或根據少數分子片段進行研究,從而缺乏系統的包括溝瓣屬大多數物種的分析。正如有的學者所說,溝瓣屬與衛矛屬的系統位置關系還需要進一步深入地分析(Simmons et al., 2012)。
數量分類學(numerical taxonomy)自20世紀60年代由Sokal和Sneath創立以來,逐漸興起并形成了與分支分類學(cladistics)、進化分類學(evolutionary taxonomy)并列的學派。數量分類學本身并不產生任何的系統,但它可以對現存的分類系統進行驗證分析,糾正一些錯誤觀點,發現以前被忽視的有用性狀 (鐘揚和陳家寬,1991)。通過數量分類學分析方法已經解決了許多分類上的疑難問題 (鐘揚和陳家寬, 1991; 戴思蘭等, 1995; 熊治廷等, 1997; 彭焱松等, 2007; 張景博等, 2010; 陳旭波等, 2012),并已在衛矛屬研究中運用,發現能較好地反映一些物種之間的相互關系,如鄭彥超等 (2012) 對衛矛屬刺果衛矛組做了數量分類學分析,認為厚葉衛矛 (E. hemsleyanus) 與紫刺衛矛 (E. angustatus)、棘刺衛矛 (E. echinatus) 與無柄衛矛 (E. subsessilis)等不應合并;Yao (2018b) 根據花的形態結構特征對衛矛屬做了數量分類學分析,研究結果顯示每心皮含胚珠數量等特征在分類學中具有重要價值。本文利用聚類分析(UPGMA)和主坐標分析(PCoA)等方法對衛矛科中衛矛屬與溝瓣屬植物之間的相互關系進行深入分析,以探討二者之間的系統分類關系。
1 材料與方法
1.1 材料
大部分材料來自中國科學院植物研究所標本館(PE)、北京師范大學生命科學院植物標本館(BNU),部分材料為作者野外采集的新鮮材料。此外,作者還到廣西、云南和海南等地進行了野外實地觀察,獲得了許多性狀數據。本文共選取溝瓣屬12種、衛矛屬中的冬青衛矛組3種、刺果衛矛組3種、衛矛組6種、深裂衛矛組3種、翅果衛矛組2種、假衛矛屬2種,共計31個OTUs(表1)。由于衛矛屬與假衛矛屬(Microtropis Wall. ex Meisn.)都為葉對生、蒴果,在系統地位上有一定距離(Simmonns & Jennifer, 1999)。因此,本文選取假衛矛屬作為衛矛屬和溝瓣屬的外類群。
1.2 方法
數量性狀使用測量尺和電子游標卡尺測量完成,定性多態性狀是根據性狀進行編碼。依據鐘揚等(1990)數量分類性狀選取原則,選取二元性狀36個、連續數值性狀9個、有序多態性狀11個, 共收集了56個性狀指標(表2)。性狀選取和編碼參考鄭彥超等(2012)和李超等(2013)的方法。
聚類分析和多變量分析均采用NTSYS-pc 2.10e軟件包進行分析(Rohlf, 2005)。
聚類分析:將原始數據轉換后,進行標準化處理;計算距離矩陣; 依據距離矩陣進行聚類分析,計算聚類樹矩陣;將聚類樹矩陣和距離矩陣加以比較并進行Mantel檢測,計算協表距離矩陣以及相關性系數,以表示該聚類樹對原始距離矩陣的代表程度;繪制聚類樹圖。運算方法采用UPGMA (unweighted pair-group method using arithmetic averages)。
多變量分析:主成分分析(PCA)和主坐標分析(PCoA)是數量分類學研究中常用的兩種多變量分析方法,由于前者不適用于對離散的定性特征分析(Sneath & Sokal, 1973; Schilling & Heiser, 1976; Kent & Coker, 1992),因此近年來主坐標分析比主成分分析應用更多(Sneath, 2000; Khalik et al., 2002; Sebola & Balkwill, 2009),本文也采用主坐標分析。具體步驟如下: 在標準化矩陣基礎上求相關系數;求出相關系數的特征值和特征向量;得到特征性狀在前三維主坐標分析中的負荷量,利用Origin 2018軟件將前三主坐標展示三維散點圖。
2 結果與分析
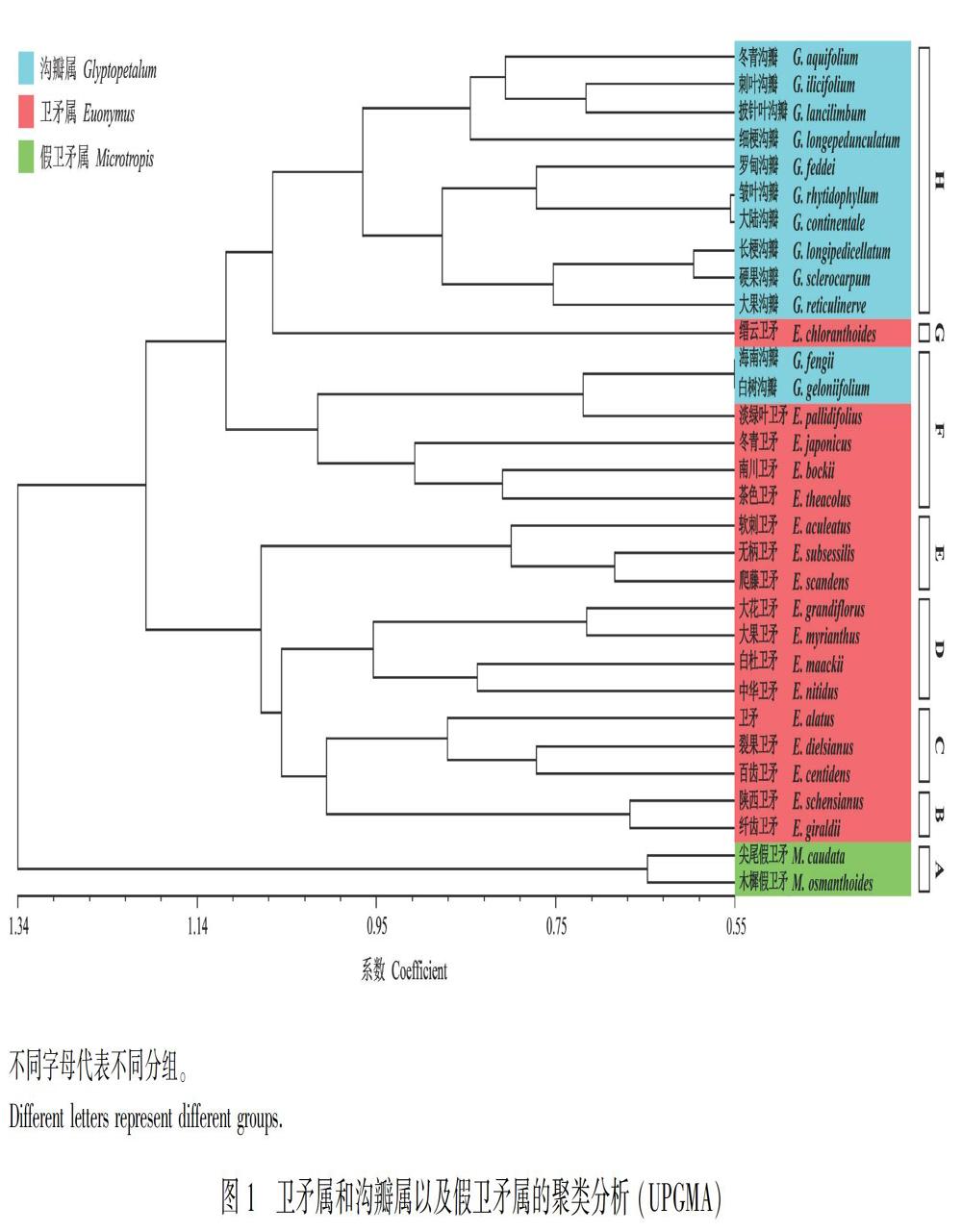
2.1 聚類分析
對衛矛屬和溝瓣屬以及假衛矛屬組成的31個OTUs進行UPGMA聚類分析的結果(圖1)顯示,大致可以將31個OTUs分成8個類群:
A類群,包括假衛矛屬(Microtropis)的2個種,即尖尾假衛矛(M. caudata)和木樨假衛矛(M. osmanthoides)。這一類群與其他幾個類群在結合線1.3處分開,其主要特征是葉對生、全緣,花5數,花瓣白色,花柱粗短,心皮2,子房每室具有2個胚珠,蒴果二裂,果皮光滑,無假種皮。
B類群,包括翅果衛矛組(Sect. Uniloculares)的陜西衛矛(Euonymus schensianus)和纖齒衛矛(E. giraldii)。其主要特征是葉對生,葉緣有纖毛狀細鋸齒,聚傘花序多花,花4數,黃綠色,心皮4,蒴果背棱延伸成翅,開裂后果皮不卷,種子全部被橘黃色假種皮包圍。
C類群,包括深裂衛矛組(Sect. Melanocarya)的衛矛(E. alatus)、百齒衛矛(E. centidens)和裂果衛矛(E. dielsianus)。其主要特征是葉對生,花4數,淺黃綠色,子房每室具有1~2個胚珠,假種皮包圍種子一半以上。
D類群,包括衛矛組(Sect. Euonymus)雙籽亞組(Subsect. Euonymus)的白杜衛矛(E. maackii)、中華衛矛(E. nitidus) 和多籽亞組(Subsect. Multiovulatae)的大果衛矛(E. myrianthus)、大花衛矛(E. grandiflorus)。其主要特征是葉對生,邊緣有鋸齒,花4數,黃白色,子房每室含4個到6個直立胚珠,蒴果淺裂,蒴果開裂后果皮不卷,假種皮半包或全包種子。
E類群,包括刺果衛矛組(Sect. Echinococcus)的軟刺衛矛(E. aculeatus)、無柄衛矛(E. subsessilis)和爬藤衛矛(E. scandens)。其主要特征是藤本狀灌木,葉對生,邊緣有鋸齒,花4數,花瓣黃白色,子房每室具有1個以上胚珠,蒴果具刺,假種皮全包種子。
F類群,包含冬青衛矛組(Sect. Ilicifolii)的冬青衛矛 (E. japonicus)、 南川衛矛(E. bockii)、 茶色衛矛(E. theacolus)、淡綠葉衛矛(E. pallidifolius)和溝瓣屬溝瓣組(Sect. Glyptopetalum)的海南溝瓣(Glyptopetalum fengii)、白樹溝瓣(G. geloniifolium)。其主要特征是葉對生,邊緣有疏鋸齒,花4數,黃綠色,花絲短,子房每室具有1個或2個倒生胚珠,蒴果圓形,光滑。
G類群,僅有1種,即縉云衛矛(Euonymus chloranthoides)。其主要特征是葉對生,邊緣有鋸齒,聚傘花序只有3花,花5數,紫紅色,花瓣邊緣有不整齊淺齒,雄蕊花絲短,子房每室含1~2個倒生胚珠,蒴果淺裂,種脊分枝,假種皮包圍種皮大部分。
H類群,主要為溝瓣屬盤狀組(Sect. Patelliformia)植物。其主要特征是葉對生,花4數,子房每室具有1個倒生的胚珠,蒴果開裂后具有宿存中軸,果皮外卷,假種皮包圍種子近一半,種脊分枝。本類群可分成2個小類群:一類為皺葉溝瓣(Glyptopetalum rhytidophyllum)、長梗溝瓣(G. longipedicellatum)、硬果溝瓣(G. sclerocarpum)、大陸溝瓣(G. continentale)、羅甸溝瓣(G. feddei)和大果溝瓣(G. reticulinerve),突出特征是喬木,葉片長超過15 cm;另一類為冬青溝瓣(G. aquifolium)、刺葉溝瓣(G. ilicifolium)、披針葉溝瓣(G. lancilimbum)和細梗溝瓣(G. longepedunculatum),主要特征是灌木,葉片長小于15 cm。
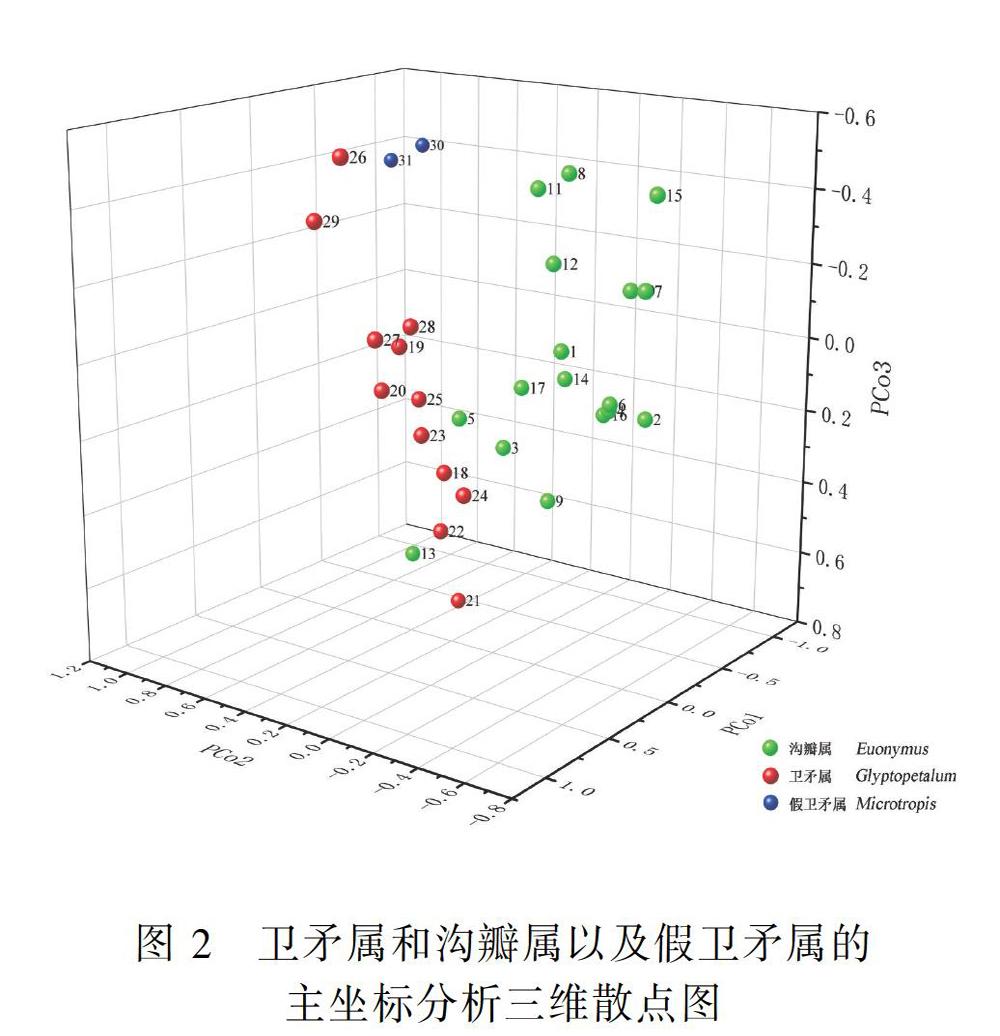
2.2 主坐標分析 (PCoA)
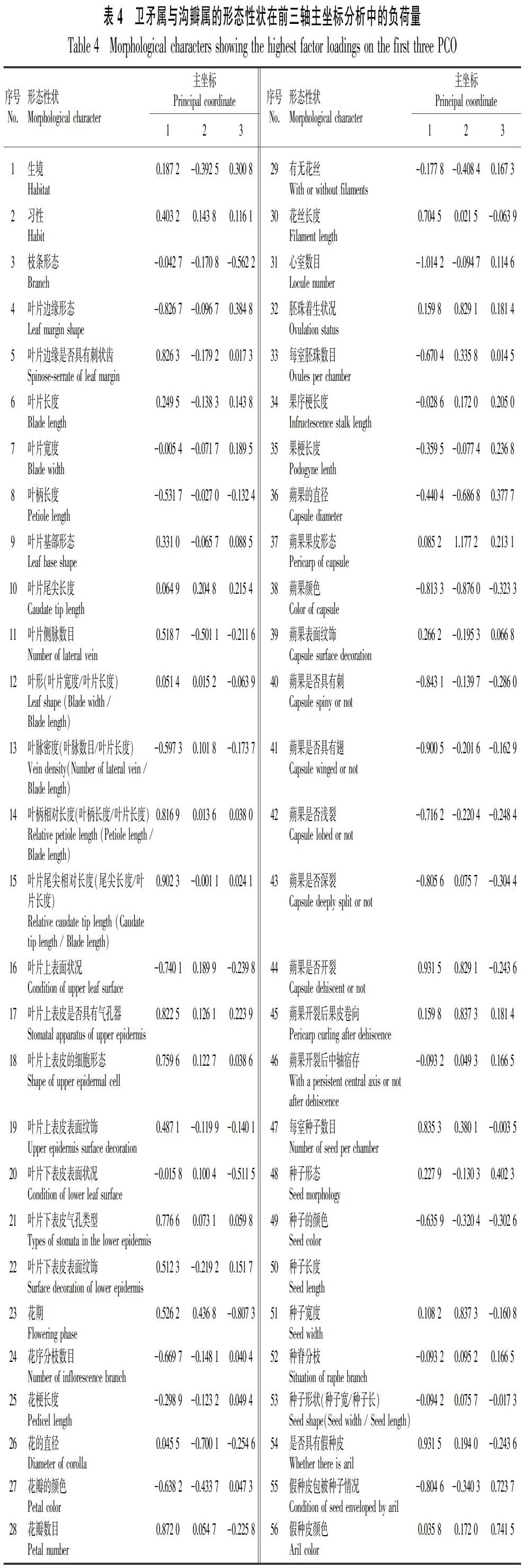
主坐標分析結果(表3)顯示,前2個主坐標的貢獻率分別為18.2652%和14.3652%,前3個主坐標的累積貢獻率為43.3994%。可見,主坐標分析的方法較適合于衛矛屬、溝瓣屬和假衛矛屬的分析。由表4可知,在第1軸坐標中,葉片長、葉片寬、葉脈數、花被數、花期、子房室數、果序梗長度、是否具有假種皮、蒴果是否開裂的負載值絕對值都在0.6以上;在第2軸坐標中,葉片長、花期、每室胚珠數的負載值絕對值在0.65以上;在第3軸坐標中,花序分枝數與果序梗長度的負載值絕對值都在0.8以上。
主坐標分析結果(圖2)顯示,31個OUTs分成2個群體,其中一個群體是假衛矛屬,另一個群體是溝瓣屬和衛矛屬的混合體。根據第1軸(占全部變量的18.2652%),從溝瓣屬逐漸過渡到衛矛屬,中間雖沒有明顯的界線,但具有過渡種,如白樹溝瓣、海南溝瓣、 縉云衛矛和冬青衛矛組的淡綠葉衛矛、冬青衛矛和茶色衛矛。根據第2軸(占全部變量的14.3652%),衛矛屬和溝瓣屬則完全混在一起。
3 討論與結論
本研究對56個性狀進行聚類分析(UPGMA)的結果顯示,溝瓣屬被聚成2個類群,這與《中國植物志》(誠靜容等, 1999)的結果一致,即一個為盤狀組(Sect. Patellifolrmia),另一個為溝瓣組(Sect. Glyptopetalum)。衛矛屬則被聚成6個類群,其中刺果衛矛組(Sect. Echinococcus)、翅果衛矛組(Sect. Kalonymus) 、深裂衛矛組(Sect. Melanocarya)和淺裂衛矛組(Sect. Euonymus)都分別聚在了一起。由此可知,聚類分析的結果基本上符合前人(誠靜容等, 1999; Ma, 2001)的研究結果。然而,溝瓣屬與衛矛屬不是如大多學者預想的那樣分成兩大獨立的類群,而是與衛矛屬交叉在一起。其中,溝瓣組(Sect. Glyptopetalum)的海南溝瓣和白樹溝瓣與冬青衛矛組(Sect. Ilicifolia)幾個代表物種聚在一起。這是因為二者的果實紋飾相似,都屬于圓果類。縉云衛矛插入溝瓣屬中,介于溝瓣屬的2個組之間。因此,由聚類圖可知,溝瓣屬與衛矛屬冬青衛矛組具有最強的表征相似性。
根據主坐標排序圖,在排序圖中雖然溝瓣屬植物和衛矛屬都分別排列在一起,但是溝瓣屬與衛矛屬在整個主坐標排序圖中排列得很緊密,中間沒有明顯的界限。在排序圖中,海南溝瓣和白樹溝瓣與冬青衛矛組的淡綠葉衛矛、南川衛矛、冬青衛矛、茶色衛矛和縉云衛矛混在一起。海南溝瓣和白樹溝瓣的種子具有明顯的分枝種脊且是4數花。縉云衛矛的花瓣具泡狀皺紋,假種皮包圍種子大部分,與種皮同色,種脊分枝雖然都是溝瓣屬的特征,但是5數花,子房每室具有1~2個胚珠又與衛矛屬相似(誠靜容等, 1999)。聚類分析圖以及主坐標排序圖顯示,縉云衛矛的位置不確定,在表征聚類分析圖中,縉云衛矛位于溝瓣屬和衛矛屬其他種之外,是介于假衛矛屬與衛矛屬和溝瓣屬之間的一個種。在主坐標排序圖中,縉云衛矛的位置是介于溝瓣屬和衛矛屬之間。綜合表征聚類分析和主坐標排序,本研究認為縉云衛矛是衛矛屬與溝瓣屬之間的過渡種。淡綠葉衛矛以前一直放在衛矛屬中(Lu & Yang, 1993; 誠靜容等, 1999; Ma, 2001; Liu & Funston, 2008),但由于具有分枝的種脊和每室具有一枚胚珠和4數花,因此最近Meng et al.(2012)將其組合到溝瓣屬,本研究結果也顯示淡綠葉衛矛在表征上與溝瓣屬更接近。南川衛矛具有4數花,果實具有糠秕狀斑點,但假種皮全部包圍種子,種脊不分枝。我們在進行中國溝瓣屬的分類學研究時,進行了大量的野外考察以及標本觀察,發現溝瓣屬與衛矛屬之間存在一些過渡種,如在衛矛屬中發現了具有4數花的植物,如白杜衛矛(E. maackii Rupr.)等;子房每室僅具有1個倒生胚珠,如曲脈衛矛(E. venosus Hemsl.)等;蒴果常具有糠秕狀斑點,如南川衛矛(E. bockii Loes.);種脊具有多條分枝,如縉云衛矛(E. chloranthoides Yang)等。這與聚類分析和主坐標分析結果一致。因此,溝瓣屬與冬青衛矛組的特征具有很多重疊,其中一些種具有過渡的特征。
Yao et al. (2018a, b)對衛矛屬和溝瓣屬的花結構研究發現,二者具有相似的花盤結構,溝瓣屬大部分種類與冬青衛矛組部分種類的蒴果上的糠秕狀斑點與刺果衛矛組的刺在發育上同源,都是花盤結構發育的結果;花粉研究顯示,溝瓣屬與衛矛屬尤其是冬青衛矛組的花粉形態結構具有較高的相似性(張芬等, 2018)。結合本研究基于葉表皮微形態特征和宏觀形態共56個性狀特征的聚類分析(UPGMA)、主坐標分析(PCoA)結果和分子系統學研究結果(Simmons et al., 2012; Li et al., 2014), 以及孟世勇(2010)關于溝瓣屬的種皮微形態特征與衛矛屬的比較研究,我們認為溝瓣屬并入到衛矛屬,置于冬青衛矛組附近更為合理。
參考文獻:
BAILLON HE, 1880. Thenatural history of plants, Vol.6 [M]. London: Reeve & Co.: 14-16.
CHEN XB, MENG SY, LIU QR, 2012. Numerical taxonomic analysis of Stellaria and Myosoton (Caryophyllaceae) [J]. Bull Bot, 47(3): 271-277. [陳旭波, 孟世勇, 劉全儒, 2012. 石竹科繁縷屬與鵝腸菜屬的數量分類 [J]. 植物學報, 47(3): 271-277.]
CHENG JR, HUANG PH, MA JS, 1999. Flora Reipublicae Popularis Sinicae [M]. Beijing: Science Press, 45(3): 80-91. [誠靜容, 黃普華, 馬景盛, 1999. 中國植物志 [M]. 北京: 科學出版社, 45(3): 80-91.]
DAI SL, ZHONG Y, ZHANG XY, 1995. Study on numerical taxonomy of some chinese species of Dendranthema (DC) Des Moul. [J]. J Beijing For Univ, 4: 9-15. [戴思蘭, 鐘楊, 張曉艷, 1995. 中國菊屬植物部分種的數量分類研究 [J]. 北京林業大學學報, 4: 9-15.]
HOU D, 1962. Celastraceae [M]//Van Steenis Flora Malesiana Series Ⅰ, Vol.6. Leiden: National Herbarium of the Netherlands: 255-265.
HOU D, 1963. Two addition alasiatic species of Glyptopetalum (Celastraceae) [J]. Blumea, 12(1): 57-60.
KENT M, COKER P, 1992. Vegetation description and analysis. A practical approach [M]. London: Belhaven Press: 1-363.
KHALIK KA, MAESEN LJGVD, KOOPMAN WJM,et al., 2002. Numerical taxonomic study of some tribes of Brassicaceae from Egypt [J]. Plant Syst Evol, 233(3-4): 207-221.
KURZ S, 1877. Forest Flora of British Burma, Vol.2 [M]. Calcutta: Office of the Superintendent of Government printing: 247-260.
LI C, LI J, ZHANG ML, 2013. A numerical taxonomical study of Epimedium L. [J]. Acta Bot Boreal-Occident Sin, 33(11):2339-2345. [李超, 李娟, 張明理, 2013. 淫羊藿屬植物的數量分類學研究 [J]. 西北植物學報, 33(11): 2339-2345. ]
LI YN, XIE L, LI JY, et al., 2014. Phylogeny of Euonymus inferred from molecular and morphological data [J]. J Syst Evol, 52(2): 149-160.
LIANG ZX, ZHANG YL, NIU LX, et al., 2014. Phenotypic diversity of Lilium browniinative to Qinba mountainous area [J]. Guihaia, 34(6): 727-733. [梁振旭, 張延龍, 牛立新, 等, 2014. 秦巴山區野百合表型多樣性 [J]. 廣西植物, 34(6): 727-733.]
LIU QR, FUNSTON AM, 2008. Glyptopetalum Thw. [M]//WU ZY, RAVEN PH, HONG DY. Flora of China, Vol. 11. Celastraceae. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press: 463.
LU SY, YANG YP, 1993. Celastraceae [M]//HUANG ZQ. Flora of Taiwan, 2nd ed, Vol. 3. Taiwan: Editorial Committee of the Flora of Taiwan: 644-651.
MA JS, 2001. A review of Euonymus (Celastraceae) [J]. Thaiszia J Bot, 11: 1-268.
MENG SY, 2010. Arevision of the Glyptopetalum Thw. (Celastraceae) in China [D]. Beijing: Beijing Normal University: 1-118. [孟世勇, 2010. 中國溝瓣屬(Celastraceae)的分類學修訂 [D]. 北京: 北京師范大學: 1-118.]
MENG SY, WANG JL, LIU QR, 2012. On the identity of Euonymus pallidifolia (Celastraceae) [J]. Ann Bot Fenn, 48(2): 185-187.
PENG YS, CHEN L, LI JQ, 2007. Study on numerical taxonomy of Quercus L.(Fagaceae) in China [J]. J Wuhan Bot Res, 25(2): 149-151. [彭焱松, 陳麗, 李建強, 2007. 中國櫟屬植物的數量分類研究 [J]. 武漢植物學研究, 25(2): 149-151.]
PRAIN D, 1891. Noviciae Indicae IV. Two additional species of Glyptopetalum [J]. J Asiatic Soc Bengal, 60(2): 205-210.
ROHLF FJ, 2005. NTSYS-PC: Numerical taxonomy and multivariate analysis system, version 2.2 [M]. Setauket: Exeter Software: 1-44
SAVINOV IA, 2014. Taxonomic revision of Asian genus Glyptopetalum Thwaites (Celastraceae B. Br.) [J]. Reinwardtia, 14(1): 183-192.
SCHILLING EE, HEISER CB, 1976. Re-examination of a numerical taxonomic study of Solanum species and hybrids [J]. Taxon, 25(4): 451-462.
SEBOLA RJ, BALKWILL K, 2009. Numerical phenetic analysis of Olinia rochetiana sensu lato (Oliniaceae) [J]. Kew Bull, 64(1): 95-121.
SIMMONS MP, JENNIFER PH, 1999. Relationships and morphological character change among genera of Celastraceae sensu lato (including Hippocrateaceae) [J]. Ann Mo Bot Gard, 86(3):723-757.
SIMMONS MP, MCKENNA MJ, BACON CD,et al., 2012. Phylogeny of Celastraceae tribe Euonymeae inferred from morphological characters and nuclear and plastid genes [J]. Mol Phylogenet Evol, 62(1): 9-20.
SNEATH PHA, 2000. Numerical classification of the chemical elements and its relation to the periodic system [J]. Found Chem, 2(3): 237-263.
SNEATH PHA, SOKAL RR, 1973. Numerical taxonomy:The Principles and Practices of Numerical Classification [M]. Freeman, San Francisco 1-573.
THWAITES GK, 1856. Description of new genera and species of Ceylon plants [J]. Hookers J Bot Kew Gard Misc, 8: 266-271.
XIONG ZT, CHEN XQ, HONG DY, 1997. Numerical taxonomic studies of Humerocallis (Liliaceae) from China [J]. Acta Phytotax Sin, 35(4) : 311-316. [熊治廷, 陳心啟, 洪德元, 1997. 中國萱草屬數量分類研究 [J]. 植物分類學報, 35(4): 311-316.]
YAO CY, DU C, ZUO YJ, et al., 2018a. The significance of floral features within infrageneric classification of Euonymus (Celastraceae) [J]. Flora, 242: 53-60.
YAO CY, ZUO YJ, DU C, et al., 2018b. Morphological study of floral nectaries in Euonymus and the probable origin of the echinate fruit surface [J]. Plant Divers, 40(1): 35-40.
ZHANG F, WANG S, LIU QR, 2018. Comparing pollen morphology of Glyptopetalum Thwaites and Euonymus L.(Celastraceae) [J]. J Beijing Norm Univ(Nat Sci Ed), 54(5): 616-621. [張芬, 王珊, 劉全儒, 2018. 溝瓣屬與衛矛屬花粉微形態比較研究 [J]. 北京師范大學學報(自然科學版), 54(5): 616-621.]
ZHANG JB, LI XD, LI JQ, 2010. Anumerical taxonomic study of the Carex section Racemosae (Cyperaceae) in China [J]. J Wuhan Bot Res, 28(3) : 279-285. [張景博, 李曉東, 李建強, 2010. 中國薹草屬黑穗薹草組的數量分類研究 [J]. 武漢植物學研究, 28(3): 279-285.]
ZHENG YC, MU XY, LI YN, et al., 2012. A numerical taxonomic study of the Euonymus section Echinococcus (Celastraceae) [J]. Plant Divers Resour, 34(3) : 271-286. [鄭彥超, 沐先運, 李研南, 等, 2012. 衛矛屬刺果衛矛組的數量分類學分析 [J]. 植物分類與資源學報, 34(3): 271-286.]
ZHONG Y, CHEN JK, 1991. A numerical taxonomic study of populations of Sagittaria pygmaea [J]. Guihaia, 11(4) : 304-307. [鐘揚, 陳家寬, 1991. 矮慈菇居群的數量分類研究 [J]. 廣西植物, 11(4): 304-307.]
ZHONG Y, CHEN JK, HUANG DS, 1990. The methods and programs for numerical taxonomy [M]. Wuchang: Whuhan University Press: 14-27. [鐘揚, 陳家寬, 黃德世, 1990. 數量分類的方法與程序 [M]. 武昌: 武漢大學出版社: 14-27.]
(責任編輯 蔣巧媛)
