RseA基因影響恥垢分枝桿菌藥物敏感性的生物信息學(xué)分析與初步驗(yàn)證
2021-03-24 05:54:26熊雨菡李智穎陳俊伊唐佳玲何永林
中國(guó)人獸共患病學(xué)報(bào) 2021年2期
關(guān)鍵詞:分析
熊雨菡,李智穎,盧 楠,陳俊伊,唐佳玲,徐 蕾,楊 春,何永林
全球每年約有1 000萬(wàn)新發(fā)結(jié)核病例,其中有150萬(wàn)人死于結(jié)核病(Tuberculosis,TB)[1]。而結(jié)核分枝桿菌多藥耐藥(MDR)和廣泛耐藥(XDR)突變株的出現(xiàn)和傳播將阻礙結(jié)核病的治療[2]。2018年,全球預(yù)計(jì)有484 000人患上了利福平(結(jié)核病最有效的一線(xiàn)藥物)耐藥結(jié)核病(RR-TB),其中78%患有多藥耐藥結(jié)核(MDR-TB)[1],而結(jié)核分枝桿菌變得對(duì)藥物治療不敏感可能是由于其進(jìn)入了非復(fù)制狀態(tài)[3]。有研究表明,在大多數(shù)個(gè)體中,最初的感染轉(zhuǎn)變?yōu)闊o(wú)癥狀的潛伏狀態(tài),結(jié)核分枝桿菌以無(wú)復(fù)制的持續(xù)狀態(tài)存在于宿主肉芽腫中,且代謝活性低[4-5]。導(dǎo)致這種生理狀態(tài)建立的不利環(huán)境因素包括低氧、營(yíng)養(yǎng)不良、pH低以及宿主免疫反應(yīng)產(chǎn)生的氧化和亞硝化應(yīng)激[5]。這種適應(yīng)主要?dú)w因于RNA聚合酶參與的能夠調(diào)節(jié)其復(fù)雜基因組表達(dá)的轉(zhuǎn)錄調(diào)控網(wǎng)絡(luò)[6]。σ因子與RNA聚合酶全酶結(jié)合,提供其對(duì)特定啟動(dòng)子的特異性,并在調(diào)節(jié)基因表達(dá)和適應(yīng)原核生物的壓力中起關(guān)鍵作用[7]。結(jié)核分枝桿菌基因組編碼13個(gè)σ因子,輔助σ因子σE是結(jié)核分枝桿菌中最典型的σ因子之一,它對(duì)于在不同壓力環(huán)境下(包括表面和氧化應(yīng)激條件)的生存至關(guān)重要[8]。我們前期研究發(fā)現(xiàn),σE可通過(guò)促進(jìn)恥垢分枝桿菌進(jìn)入持留狀態(tài)影響抗結(jié)核藥物的敏感性[9]。
近來(lái),抑制σ因子功能的抗σ因子也受到越來(lái)越多的關(guān)注。結(jié)核分枝桿菌的抗σE因子RseA由Rv1222基因編碼。恥垢分枝桿菌是一種在安全性、生長(zhǎng)特性和遺傳易感性方面用于分枝桿菌研究的模式生物,已被廣泛用于抗生素的作用和耐藥性研究[10]。恥垢分枝桿菌的RseA由基因MSMEG_5071編碼。RseA編碼基因緊鄰下游σE,已被證明是可直接調(diào)控σE的特異性抗σE因子[11],而至今針對(duì)RseA在抗結(jié)核藥物敏感性的機(jī)制研究較少。因此,本實(shí)驗(yàn)構(gòu)建了重組恥垢分枝桿菌pMV261-MSMEG-5071/MS,通過(guò)轉(zhuǎn)錄組測(cè)序技術(shù)和生物信息學(xué)方法篩選出差異表達(dá)基因(differentially expression genes,DEGs),并對(duì)其進(jìn)行了基因本體論(Gene Ontology,GO)和京都基因與基因組百科全書(shū)(Kyoto Encyclopedia of Genes and Genomes,KEGG)分析,構(gòu)建了蛋白-蛋白相互作用(protein-protein interaction,PPI)網(wǎng)絡(luò),通過(guò)MCODE插件篩選出可能受RseA調(diào)控的基因及其可能的分子機(jī)制并進(jìn)行初步驗(yàn)證,為耐藥結(jié)核病的治療提供新思路。
1 材料與方法
1.1實(shí)驗(yàn)材料 質(zhì)粒pMV261、E.coliDH5α和恥垢分枝桿菌(Mycobacteriumsegmatismc2155,MS)和RpfE蛋白由實(shí)驗(yàn)室保存。HindⅢ、BamH Ⅰ和T4連接酶(Thermo Fisher Scientific),DAB顯色試劑盒(康為世紀(jì)),卡那霉素(Genview),膠回收試劑盒、質(zhì)粒提取試劑盒、蛋白Marker(Takara),小鼠抗組氨酸單克隆抗體(Bioworld Technology),HRP標(biāo)記的羊抗兔IgG(美國(guó)Signalway Antibody),SDS-PAGE凝膠配置試劑盒(EpiZye),異煙肼、利福平、乙胺丁醇、左氧氟沙星、環(huán)丙沙星、鏈霉素(北京睿博興科)。
1.2方 法
1.2.1重組恥垢分枝桿菌的構(gòu)建 根據(jù)NCBI GenBank(基因序列號(hào)NC_008596.1)中恥垢分枝桿菌MSMEG_5071的基因序列設(shè)計(jì)引物。以MS基因組為模板進(jìn)行PCR擴(kuò)增,將PCR產(chǎn)物和質(zhì)粒pMV261分別經(jīng)BamHⅠ/HindⅢ酶切并用T4連接酶連接后,轉(zhuǎn)化至E.coliDH5α感受態(tài)細(xì)胞中,涂布在含有卡那霉素的LB固體培養(yǎng)基上,長(zhǎng)出單菌落后,挑取單菌落培養(yǎng)并測(cè)序驗(yàn)證。將構(gòu)建成功的pMV261-MSMEG_5071重組質(zhì)粒經(jīng)2.5 KV、4 ms電轉(zhuǎn)至恥垢分枝桿菌感受態(tài)中,涂布在含有卡那霉素的7H10固體培養(yǎng)基上,長(zhǎng)出單菌落后,挑取單菌落培養(yǎng)以獲得重組恥垢分枝桿菌pMV261-MSMEG_5071/MS。
1.2.2Western blot驗(yàn)證蛋白R(shí)seA的表達(dá) 將構(gòu)建成功的重組恥垢分枝桿菌pMV261-MSMEG_5071/MS接種于含有卡拉霉素的7H9培養(yǎng)基中,37 ℃培養(yǎng)至OD600為0.6~0.8,分別于40 ℃、42 ℃、44 ℃和46 ℃熱誘導(dǎo)3 h后,37 ℃,160 r/min培養(yǎng)4 h,離心收集菌體后,于-80 ℃反復(fù)凍融3次,加入無(wú)菌PBS混勻,超聲碎菌,離心后分別收集上清與沉淀。將樣品經(jīng)金屬浴100 ℃ 10 min變性,通過(guò)Western blot驗(yàn)證。
1.2.3樣本準(zhǔn)備及轉(zhuǎn)錄組測(cè)序 將重組菌接種于含有卡那霉素的7H9培養(yǎng)基中,培養(yǎng)至OD600為0.8,42 ℃熱誘導(dǎo)3 h,加入10 μg /mL EMB,37 ℃,180 r/min培養(yǎng)48 h,離心收集菌體后,PBS洗滌3次,置于液氮盒中,送至上海美吉生物醫(yī)藥科技有限公司進(jìn)行轉(zhuǎn)錄組學(xué)測(cè)序和生物信息學(xué)分析。
1.2.4測(cè)序數(shù)據(jù)質(zhì)控 通過(guò)Illumina Hiseq 測(cè)序獲得的原始圖像經(jīng)base calling轉(zhuǎn)換為序列數(shù)據(jù),即原始數(shù)據(jù)raw data。使用Sickle和SeqPrep質(zhì)控軟件除去reads中的adapter序列、5′端含有非A、G、C、T的堿基和含N的比例達(dá)到10%的reads,修剪測(cè)序質(zhì)量較低的reads末端 (測(cè)序質(zhì)量值小于Q20),棄置經(jīng)去adapter及質(zhì)量修剪后長(zhǎng)度小于25 bp的小片段,得到高質(zhì)量reads,即clean data,并對(duì)數(shù)據(jù)進(jìn)行質(zhì)量評(píng)估。通過(guò)Diamond分析軟件進(jìn)行核糖體RNA污染率評(píng)估,若rRNA含量低于15%,則可進(jìn)行后續(xù)分析。
1.2.5DEGs分析 使用Bowtie2軟件計(jì)算出每組中各個(gè)基因的表達(dá)量,通過(guò)DEGseq分析RseA基因過(guò)表達(dá)恥垢分枝桿菌組及空質(zhì)粒對(duì)照組的DEGs。篩選差異倍數(shù)(fold change)≥2且P≤0.001的基因?yàn)轱@著DEGs。
1.2.6GO功能富集和KEGG信號(hào)通路分析 GO是一個(gè)在生物信息學(xué)領(lǐng)域中廣泛使用的本體,是實(shí)現(xiàn)統(tǒng)一基因相關(guān)數(shù)據(jù)、進(jìn)行數(shù)據(jù)轉(zhuǎn)換和開(kāi)展數(shù)據(jù)挖掘的基礎(chǔ),包括分子功能(molecular function,MF)、細(xì)胞組分(cellular components,CC)和生物學(xué)過(guò)程(biological process,BP)。KEGG是從分子水平信息了解高級(jí)功能和生物系統(tǒng)的實(shí)用程序數(shù)據(jù)庫(kù)資源。本實(shí)驗(yàn)通過(guò)DAVID6.8對(duì)DEGs進(jìn)行GO分析及KEGG分析。P<0.05為功能和通路具有統(tǒng)計(jì)學(xué)意義。
1.2.7PPI網(wǎng)絡(luò)的構(gòu)建 STRING(https://string-db.org/)是可用于PPI網(wǎng)絡(luò)功能富集分析的在線(xiàn)數(shù)據(jù)庫(kù)之一。在STRING中輸入DEGs,以得到其PPI關(guān)系(sore>0.9),通過(guò)Cytoscape 3.7.2軟件構(gòu)建PPI網(wǎng)絡(luò)。通過(guò)復(fù)雜分子檢測(cè)(molecular complex detection,MCODE)插件篩選出最重要模塊,對(duì)模塊進(jìn)行GO功能富集分析和KEGG信號(hào)通路分析,P<0.05差異具有統(tǒng)計(jì)學(xué)意義。并篩選出模塊中連接度最高的前8個(gè)hub基因。
1.2.8復(fù)蘇指數(shù)的測(cè)定 將重組菌培養(yǎng)至生長(zhǎng)對(duì)數(shù)期,調(diào)節(jié)OD600至0.5,用含有卡那霉素的7H9液體培養(yǎng)基洗滌2次,離心后分別用含有100 μg /mL異煙肼(isoniazid,INH)、25 μg /mL利福平(rifampin,RIF)、10 μg /mL乙胺丁醇(ethambutol,EMB)、10 μg /mL左氧氟沙星(levofloxacin,LVFX)、10 μg /mL環(huán)丙沙星和6.4 μg /mL鏈霉素(streptomycin,STRE)的7H9培養(yǎng)基重懸,37 ℃,180 r/min培養(yǎng)48 h。離心棄上清,用7H9培養(yǎng)基重懸后分為2組。一組等倍梯度稀釋至106,所有濃度的菌液各吸取10 μL加至7H9固體培養(yǎng)基上,37 ℃培養(yǎng)3 d,計(jì)數(shù);另一組用含有 20 ng/mL RpfE(可以促進(jìn)非復(fù)制的持留菌復(fù)蘇生長(zhǎng)[12])的7H9液體培養(yǎng)基等倍梯度稀釋至106,每濃度菌液吸取200 μL加至96孔板,并設(shè)置復(fù)孔,96孔板四周用200 μL 7H9液體培養(yǎng)基封板,于37 ℃培養(yǎng)2周后,觀察并記錄陽(yáng)性細(xì)菌孔,查表得到最大可能數(shù)(Most probable number,MPN)。通過(guò)log10(MPN)-log10(CFU)計(jì)算得出的復(fù)蘇指數(shù)(Resuscitation Index,RI)來(lái)表示其復(fù)蘇能力[13]。
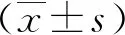
2 結(jié) 果
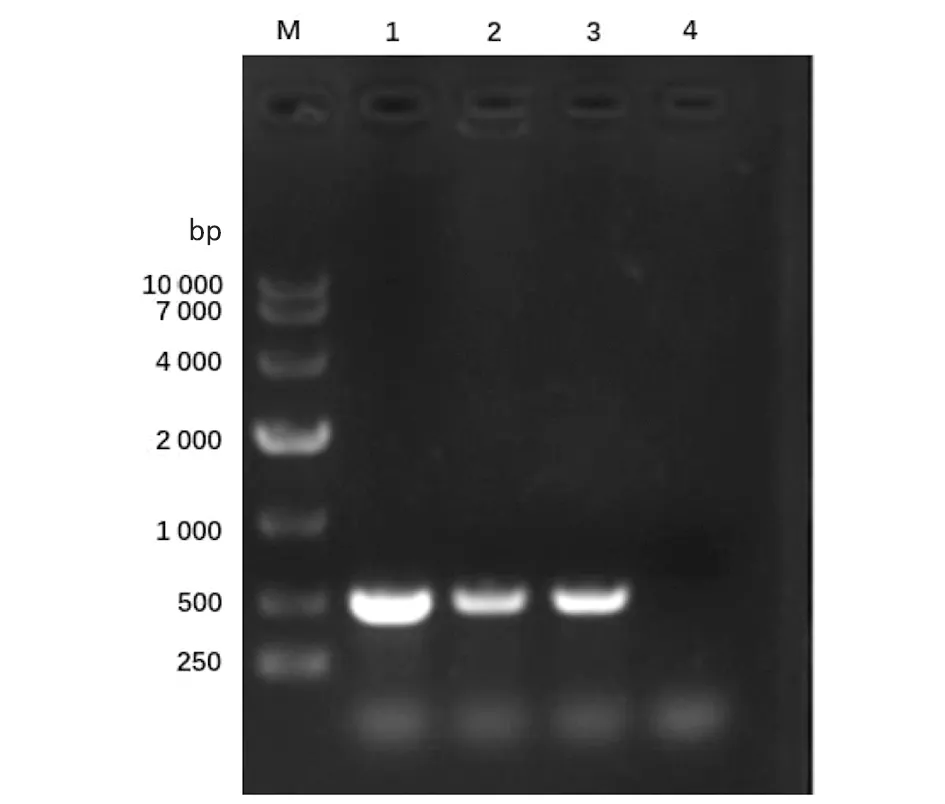
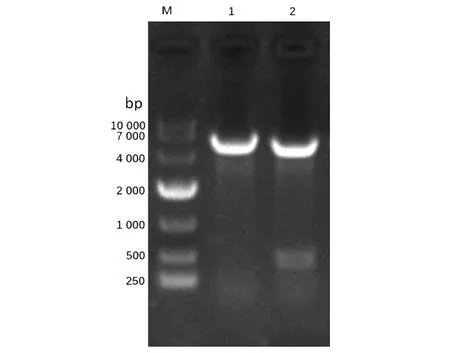
2.1重組恥垢分枝桿菌pMV261-MSMEG_5071/MS的構(gòu)建及驗(yàn)證 以MS基因組為模板,擴(kuò)增得到的PCR產(chǎn)物大小約為400 bp,與預(yù)期大小一致(見(jiàn)圖1)。將構(gòu)建成功的重組質(zhì)粒pMV261-MSMEG_5071經(jīng)BamHⅠ/HindⅢ酶切后,酶切片段大小與預(yù)期一致(圖2)。將重組質(zhì)粒pMV261-MSMEG_5071電轉(zhuǎn)入恥垢分枝桿菌,40 ℃、42 ℃、44 ℃和46 ℃誘導(dǎo)表達(dá),超聲碎菌處理,離心后,收集上清與沉淀,經(jīng)Western blot驗(yàn)證,獲得分子量為17 kD左右的條帶,與預(yù)期大小一致(圖3)。
2.2兩組間DEGs分析結(jié)果 轉(zhuǎn)錄組測(cè)序結(jié)果顯示,兩組間共篩選出2 403個(gè)DEGs,其中2 335個(gè)上調(diào)基因,68個(gè)下調(diào)基因。將2 403個(gè)DEGs繪制成火山圖,其中紅色代表上調(diào)基因,綠色代表下調(diào)基因(圖4)。
M.DNA marker;1-3.MSMEG-5071;PCR product is about 400bp;4.H2O as a control.
M.DNA Marker;1.pMV261-MSMEG-5071;2.Double digestion
M.Marker;1-8.supernatant and precipitation of pMV261-MSMEG-5071 lysate induced at 40 ℃,42 ℃,44 ℃,46 ℃;9.No-load strain induced at 42 ℃ for 3 h.
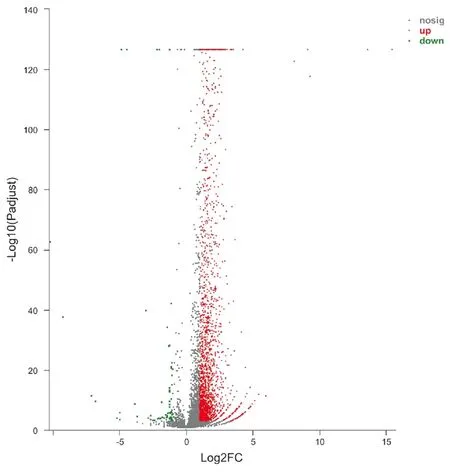
圖4 DEGs火山圖
2.3GO功能富集和KEGG信號(hào)通路分析 將篩選出的DEGs進(jìn)行GO分析,發(fā)現(xiàn)DEGs共參與了155個(gè)生物學(xué)過(guò)程(P<0.05),其中MF主要富集在轉(zhuǎn)錄因子活性,序列特異性DNA結(jié)合、核酸結(jié)合轉(zhuǎn)錄因子活性和核酸結(jié)合;CC主要富集在細(xì)胞內(nèi),轉(zhuǎn)移酶復(fù)合物,轉(zhuǎn)移含磷基團(tuán);BP主要富集在氮化合物代謝過(guò)程的調(diào)控、RNA代謝過(guò)程的調(diào)控和細(xì)胞代謝過(guò)程的調(diào)控等過(guò)程。GO富集的部分生物學(xué)過(guò)程見(jiàn)圖5。KEGG分析顯示,DEGs參與了8個(gè)KEGG信號(hào)通路(P<0.05),主要富集在萜類(lèi)骨架的生物合成、果糖和甘露糖代謝和同源重組等通路(表1)。
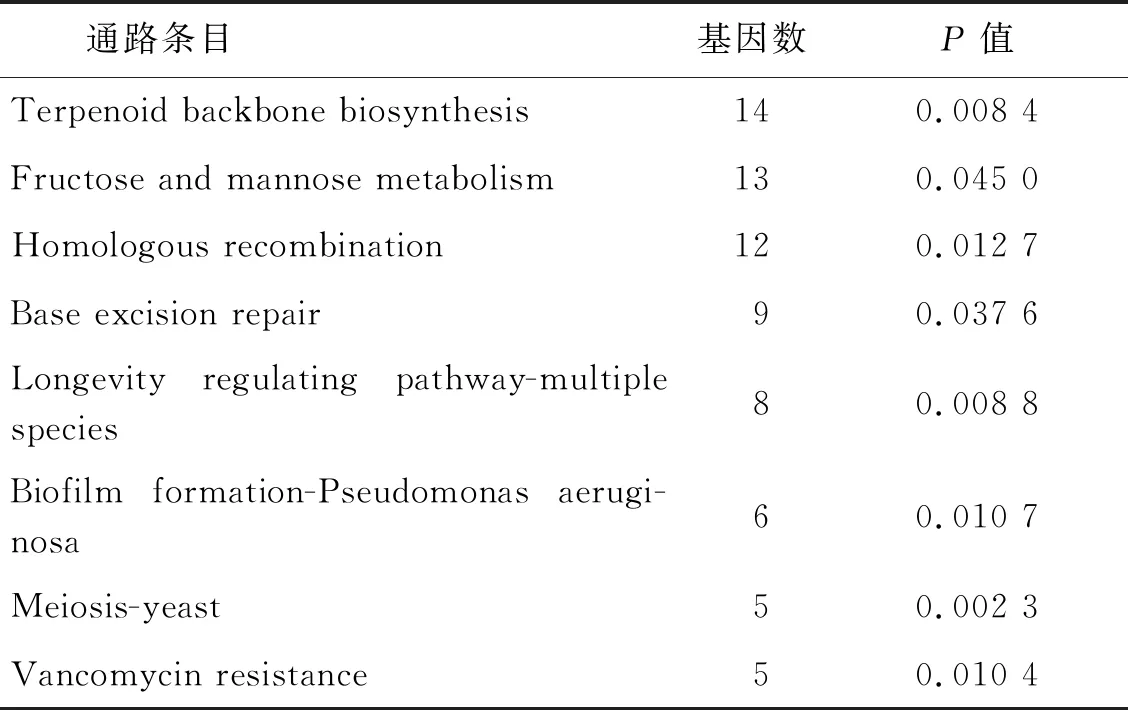
表1 DEGs的KEGG通路分析
2.4PPI網(wǎng)絡(luò) 通過(guò)STRING在線(xiàn)分析軟件獲得DEGs的PPI關(guān)系,構(gòu)建PPI網(wǎng)絡(luò)。結(jié)果顯示,共有1 136條邊緣數(shù)和594個(gè)hub基因(圖6)。使用插件MCODE分析出的最重要模塊含有16個(gè)hub基因和119條邊緣數(shù)(圖7)。對(duì)這16個(gè)hub基因進(jìn)行GO分析和KEGG分析。GO分析結(jié)果顯示,這16個(gè)hub基因主要富集在翻譯、核糖體的結(jié)構(gòu)成分和核糖體等功能(表2)。KEGG分析結(jié)果表明,其主要富集在核糖體通路(表3)。
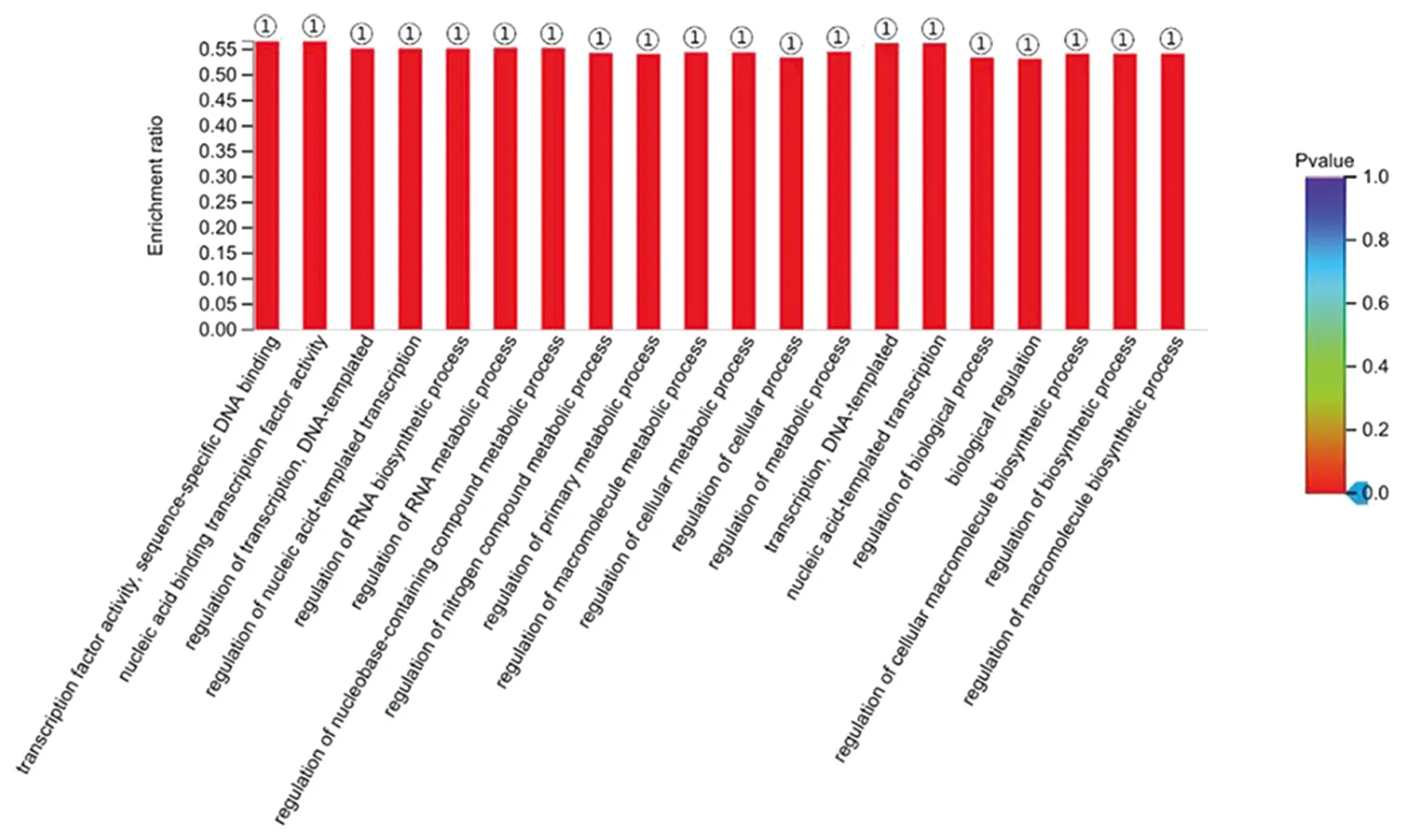
①:P<0.001

表2 模塊內(nèi)基因的GO富集分析

表3 模塊內(nèi)基因的KEGG通路分析

圖6 PPI網(wǎng)絡(luò)圖

圖7 PPI中MCODE分析得到的最高分子模塊
2.5RseA過(guò)表達(dá)對(duì)藥物誘導(dǎo)的持留菌復(fù)蘇情況的影響 分別用高濃度藥物INH、RIF、EMB、LVFX、CPFX和STRE處理細(xì)菌48 h,誘導(dǎo)細(xì)菌處于非復(fù)制狀態(tài),用含有20 ng/mL RpfE蛋白的7H9促進(jìn)持留菌復(fù)蘇,觀察兩組菌復(fù)蘇情況。INH、RIF、EMB、LVFX和CPFX 5種藥物處理后,MSMEG_5071過(guò)表達(dá)組復(fù)蘇指數(shù)明顯低于對(duì)照組(圖8),差異有統(tǒng)計(jì)學(xué)意義(tINH=4.37,tRIF=7.26,tEMB=6.75,tLVF=11.13,tCPFX=8.24,tSTRE=0.02;P<0.05,PINH=0.011 966,PRIF=0.001 912,PEMB=0.002 512,PLVF=0.000 371,PCPFX=0.001 185,PSTRE=0.987)。
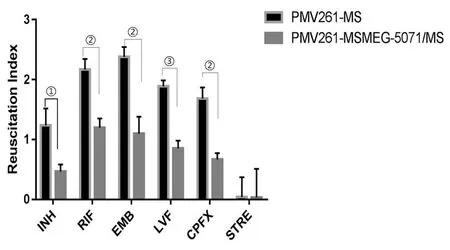
①P<0.05,②P<0.01,③P<0.001
3 討 論
本研究通過(guò)生物信息學(xué)分析,比較了pMV261-MSMEG_5071和含有空質(zhì)粒pMV261的恥垢分枝桿菌的轉(zhuǎn)錄組測(cè)序結(jié)果,共篩選出2 403個(gè)DEGs,其中上調(diào)基因2 335個(gè),下調(diào)基因68個(gè)。同時(shí),分析了這些基因的PPI關(guān)系,構(gòu)建了PPI網(wǎng)絡(luò)并通過(guò)MCODE插件分析,篩選了RseA影響恥垢分枝桿菌藥物敏感性的關(guān)鍵靶向基因。其中,篩選出的前8個(gè)基因,rpsG、rplM、rpsO、rpsT、rpmH、rplS、rpsJ、rplT可能在RseA影響恥垢分枝桿菌藥物敏感性過(guò)程中具有重要作用。
rpsG編碼核糖體蛋白S7,該蛋白位于核糖體小亞基(30S)頭部,是唯一與mRNA交聯(lián)的顯著靶標(biāo),也是通過(guò)翻譯反饋機(jī)制控制核糖體蛋白合成的主要調(diào)控元件之一,對(duì)于30S亞基的組裝至關(guān)重要[14-16]。rplM編碼核糖體蛋白L13,是核糖體大亞基(50S)組裝中的必需蛋白[16]。有研究發(fā)現(xiàn),恥垢分枝桿菌MSMEG_1556(rplM)敲除株無(wú)法在42 ℃條件下生長(zhǎng),由此證明了MSMEG_1556(rplM)對(duì)于恥垢分枝桿菌的生長(zhǎng)至關(guān)重要[17]。rpsO編碼核糖體蛋白S15,是位于30S亞基平臺(tái)上的唯一一級(jí)結(jié)合蛋白。S15與16S rRNA的預(yù)先結(jié)合是二級(jí)和三級(jí)核糖體蛋白(r蛋白)S6、S18、S11、S21與新興復(fù)合物結(jié)合所必需的[18]。由此證明,S15參與了亞基締合并在30S亞基組裝中發(fā)揮了重要作用[19]。rpsT編碼核糖體30S亞基蛋白S20,該蛋白直接與16S rRNA結(jié)合,并獨(dú)立于其他蛋白,被定義為一級(jí)結(jié)合蛋白,由此表明S20在30S亞基的組裝中起著關(guān)鍵作用[20]。并有研究表明,缺乏S20的菌株生長(zhǎng)速度明顯降低[21]。rpmH編碼核糖體大亞基蛋白L34,L34的缺乏會(huì)導(dǎo)致70S核糖體形成的嚴(yán)重缺陷和生長(zhǎng)速率的降低[22]。rplS編碼核糖體50S蛋白L19,L19可以抵御抗體,并且與氯霉素的結(jié)合有關(guān)[23]。rpsJ編碼核糖體30S亞基蛋白S10,有實(shí)驗(yàn)表明,該蛋白是參與轉(zhuǎn)錄抗終止作用的細(xì)胞成分之一[24]。rplT編碼核糖體50S蛋白L20,參與50S亞基的早期組裝步驟以及自身基因表達(dá)的反饋控制[25]。
PPI網(wǎng)絡(luò)模塊分析顯示,RseA過(guò)表達(dá)導(dǎo)致多種核糖體蛋白異常表達(dá),核糖體的結(jié)構(gòu)蛋白合成和翻譯通路富集。核糖體是核糖核蛋白復(fù)合物,由具有21個(gè)蛋白和16S rRNA的小亞基(30S)以及具有36個(gè)蛋白,23S rRNA和5S rRNA的大亞基(50S)組成,已成功被多種抗生素所靶向[26]。核糖體執(zhí)行蛋白質(zhì)生物合成的關(guān)鍵功能,主要分為翻譯起始、延伸、終止和再循環(huán)4個(gè)階段,而翻譯是適應(yīng)某些環(huán)境條件(例如饑餓,物理或化學(xué)壓力,尤其是休眠)的主要監(jiān)管目標(biāo)[27]。因此,推測(cè)RseA過(guò)表達(dá)后,主要通過(guò)涉及核糖體的翻譯過(guò)程影響恥垢分枝桿菌對(duì)環(huán)境的適應(yīng)能力,從而影響恥垢分枝桿菌的藥物敏感性。
我們采用復(fù)蘇指數(shù)來(lái)衡量持留菌的數(shù)量多少。使用高濃度藥物INH、RIF、EMB、LVFX、CPFX和STRE處理細(xì)菌,使其處于非復(fù)制的持留狀態(tài),通過(guò)復(fù)蘇促進(jìn)因子(RpfE)復(fù)蘇后,計(jì)數(shù)最大可能數(shù)MPN,即包括持留菌在內(nèi)的所有活菌;同時(shí)計(jì)數(shù)未通過(guò)RpfE處理的菌落CFU,即不包含持留菌的活菌,兩者之差即為復(fù)蘇指數(shù)RI,以此來(lái)反應(yīng)持留菌的多少。實(shí)驗(yàn)結(jié)果觀察到,INH、RIF、EMB、LVFX、CPFX藥物作用后,RseA過(guò)表達(dá)組的復(fù)蘇指數(shù)明顯低于對(duì)照組,即RseA過(guò)表達(dá)可能導(dǎo)致持留菌減少;STRE作用后,RseA過(guò)表達(dá)組與對(duì)照組無(wú)差異。而STRE是一種氨基糖苷類(lèi)抗生素,通過(guò)靶向核糖體,引起遺傳基因誤讀干擾翻譯啟動(dòng),抑制細(xì)菌蛋白質(zhì)合成,從而殺滅細(xì)菌或抑制細(xì)菌生長(zhǎng)[28]。由此可見(jiàn),過(guò)表達(dá)RseA可能影響了核糖體通路,從而影響恥垢分枝桿菌對(duì)藥物的敏感性。
綜上所述,本研究通過(guò)生物信息學(xué)分析,發(fā)現(xiàn)RseA在恥垢分枝桿菌藥物敏感性的影響中發(fā)揮了重要作用,為RseA以后在結(jié)核分枝桿菌的治療應(yīng)用提供了新的思路,后續(xù)將開(kāi)展大量的基礎(chǔ)研究對(duì)此結(jié)果進(jìn)行進(jìn)一步的驗(yàn)證。
利益沖突:無(wú)
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財(cái)經(jīng)界(學(xué)術(shù)版)(2015年20期)2015-12-23 09:20:13
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
