肺癌線粒體DNA誘導小鼠胚胎成纖維細胞NIH3T3惡性轉化的研究
2021-03-24 13:33:56姬宏利,姬宏娟,王保健
安徽醫藥 2021年3期
細胞核內的遺傳物質,它在腫瘤的形成和發展過程中起著重要的作用。核外的線粒體DNA(mitochondrial DNA,mtDNA)是在具有DNA基因組的細胞中存在的唯一的核外遺傳物質。這種遺傳物質具有DNA基因組,能夠自主復制。因線粒體內氧濃度較高,長期暴露于這種高濃度氧環境下的mtDNA容易發生突變。其中mtDNA的1 120 bp(16025-576)組成的D環區是突變的熱點所在,其在腫瘤發生發展中的作用正備受關注,在胃癌、腸癌、鼻咽癌等實體惡性腫瘤中,已見到針對該區域相關突變位點的相關報道,但這些報道均未涉及mtDNA在惡性腫瘤發生和演變中的作用和機制。本研究于2017年1月至2019年6月將肺癌細胞突變mtDNA片段轉染小鼠胚胎成纖維細胞NIH3T3,觀察mtDNA片段在成纖維細胞核內的整合情況,并觀察轉染細胞的生物學行為變化,以探討變異的mtDNA在腫瘤發病機制中的作用。
1 材料與方法
1.1 材料
小鼠胚胎成纖維細胞NIH3T3及肺癌細胞A549、H226購于上海福祥生物科技有限公司。G480、脂質體轉染Lipofection2000轉染試劑均購自英韋創津(Invintrogen)公司;DMEM高糖培養基、胎牛血清(FBS)、胰蛋白酶購自Gibco公司;3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴鹽(MTT),二甲基亞砜(DMSO)、姬姆素染料(Giemsa stain)購自Sigma-Aldrich公司。本課題組將突變的肺癌細胞mtDNA的D環區插入到pcDNA3.1(+)載體BamH I,構建了XhoⅠ位點間的重組表達載體。分別命名為pcDNA3.1(+)-A549 mtDNA、pcDNA3.1(+)-H226 mtDNA;mtDNA提取試劑盒購自杭州維特潔(V-gene)生物技術公司;熒光原位雜交(fluorescence in situ hybridization,FISH)試劑盒購于天津灝洋生物科技公司;膜聯蛋白V-異硫氰酸熒光素(Annexin V-FITC)是從上海碧云天生物科技有限公司購得的。流式細胞儀購自Epics Elite,Coulter Corporation,U.S.A。1.2 方法
1.2.1
細胞培養 將NIH3T3、A549及H226細胞在含100 mL/L胎牛血清的高糖DMEM培養基中傳代培養,待生長至對數生長期時,用適當量的9 g/L胰蛋白酶消化后,培養貼壁生長良好的細胞。1.2.2
細胞轉染與篩選 將胰蛋白酶消化傳代培養處于對數生長期的NIH3T3細胞,先以計數板計數細胞濃度后,細胞濃度進一步調整到大約1×10個/毫升,將2 mL的細胞懸液接種于6孔板。用脂質體轉染法將重組載體(pcDNA3.1(+)-A549 mtDNA、pcDNA3.1(+)-H226 mtDNA)轉染NIH3T3細胞,具體轉染操作過程參考英偉創津(Invintrogen)公司的Lipofection2000說明書。將轉染了重組載體的6孔板,置入二氧化碳飽和濕度的培養箱中培養24 h后,以遺傳霉素418(Geneticin418,G418)行抗性篩選。將轉染了重組載體的NIH3T3分別命名為NIH3T3/A549-mtDNA、NIH3T3/H226-mtDNA。1.2.3
NIH3T3細胞染色體核型分析 將不同組別的細胞分別加入25 mg/L秋水仙素2 h,終止細胞分裂,收集細胞;加入37℃預熱的0.075 mol/L 8 mL的氯化鉀溶液20 min,加入新鮮配制的固定液,2 000 r/min離心5 min,棄上清,重復兩次;細胞懸液滴在預冷的濕片上,烤干,9 g/L胰蛋白酶,不添加乙二胺四乙酸(EDTA)EDTA,2%的Gimsa染色5 min后,油鏡下觀察染色體。1.2.4
FISH mtDNA探針由天津灝洋生物制品科技有限公司設計,通過PCR法合成。探針3"端標記地高辛,5"端標記熒光素。探針1:A aactccaccattagcaccc;A"ggtggctggcagtaatgta,產物大小144;探針2:B ctgccagccaccatgaatat;B"gggacgagaagggatttg,產物大小262。將探針稀釋成終濃度為5 mg/L,當在75℃恒溫水浴中孵育5 min,并立即放置在0℃5 min以變性mtDNA探針,用70%甲酰胺/2×檸檬酸鈉緩沖液(SSC)在75℃下變性2~3 min。將樣品用70%、90%和100%的冰乙醇梯度脫水5 min,然后干燥空氣。標本加100μL含變性的mtDNA探針的雜交液[雜交液里含50%去離子甲酰胺、5×SSC、10%硫酸葡聚糖、5×Denhardt液、2%十二烷基磺酸鈉(SDS)100μg/mL,硅精魚DNA],42℃雜交過夜。用50%甲酰胺/2×SSC洗滌3次,5分鐘/次,1×SSC洗滌3次,5分鐘/次,室溫干燥玻片。滴加抗地高辛熒光抗體免疫球蛋白G(IgG)液(0.2 g/L)與小牛血清(終濃度為5%)的混合液,37℃避光溫育20 min以放大信號,經封片,在激光顯微鏡下觀察染色體間期細胞核上的黃綠色熒光。1.2.5
四甲基偶氮唑鹽微量酶反應比色法(MTT法)測定細胞增殖 以每孔1.5×10的單細胞懸液種植于96孔板,加入200μL的DMEM PRMI-1640高糖培養基,每組設3個復孔,在不同的時間點,添加MTT溶液30μL,繼續培養4 h,并丟棄上清液,加入DMSO 200μL,30 min后在570 nm波長下,通過酶聯免疫吸附測定(ELISA)測定孔光的吸收值,并記錄結果。實驗重復3次,記錄不同組細胞0 h、24 h、48 h、72 h 570 nm所測吸光度的數據。1.2.6
流式細胞儀檢測細胞凋亡 棄培養基,細胞與1μL與Annexin V-FITC混合2 min后,加入5μL碘化丙啶。細胞在室溫下培養5 min。流式細胞儀檢測細胞凋亡。
2 結果
2.1 轉染前較轉染后NIH3T3細胞染色體核型分析
NIH3T3是小鼠胚胎成纖維細胞,正常小鼠染色體數目為40條,20對(包括性染色體)。圖1示:NIH3T3/A549-mtDNA、NIH3T3/H226-mtDNA的染色體出現了易位(箭頭1所示)、斷裂(箭頭2所示)的染色體結構畸形。結果表明,轉染肺癌細胞mtDNA的NIH3T3細胞具有惡性轉化的趨勢。不同視野下畸變的多倍體數目和染色體數目占全部染色體數目百分比具體見表1。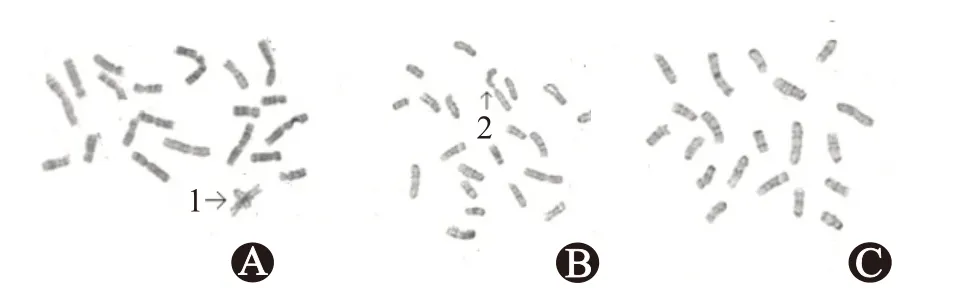
圖1 轉染前后小鼠胚胎成纖維細胞NIH3T3染色體核型(×1 000):A為NIH3T3/A549-線粒體DNA(mtDNA)的核型;B為NIH3T3/H226-mtDNA的核型;C為NIH3T3的核型
表1 不同視野小鼠胚胎成纖維細胞NIH3T3的畸形的多倍體數目和染色體數目占全部染色體數目百分比/(%,±s)
2.2 熒光原位雜交結果
MtDNA探針是一種分離重排探針,針對處于有絲分裂間期細胞核,此期細胞核因未進行有絲分裂而處于無絲狀結構的狀態,根據堿基互補配對原則,有熒光物質的探針與目標DNA結合后,融合成黃色的陽性雜交信號。本實驗中,通過熒光顯微鏡觀察到2條mtDNA探針在NIH3T3/A549-mtDNA、NIH3T3/H226-mtDNA的細胞染色體間期核中均出現黃色的陽性雜交信號;在未轉染重組載體的NIH3T3細胞染色體間期核中無陽性雜交信號。說明突變的mtDNA能夠整合在處于有絲分裂間期的NIH3T3細胞核中。2.3 轉染后NIH3T3細胞的增殖率較轉染前NIH3T3增加
用MTT法檢測不同組別NIH3T3細胞在570 nm波長下的吸光度,發現經轉染了不同重組載體的NIH3T3,經培養24 h、48 h、72 h的吸光度高于未轉染的NIH3T3細胞;且同一組別NIH3T3細胞隨著培養時間的增加,吸光度逐漸增加。進一步反映出,隨時間增加轉染后NIH3T3細胞的增殖率高于轉染前NIH3T3細胞。見表2。2.4 轉染后NIH3T3細胞的凋亡率較轉染前NIH3T3降低
流式細胞儀檢測不同組別NIH3T3細胞的凋亡率,NIH3T3/A549-mtDNA組(5.20±0.20)%、NIH3T3/H226-mtDNA組(7.75±0.11)%較NIH3T3組(19.04±0.08)%細胞的凋亡率明顯下降(F
=8 381.21,P<
0.001)。說明轉染了重組表達載體后,在一定程度上抑制了NIH3T3細胞的凋亡。3 討論
腫瘤的發生、發展和人類對腫瘤的易感性、基因突變,遺傳物質的改變等多因素密切相關。核DNA(cDNA)是細胞核內的重要遺傳物質,具有一定的穩定性,但突變的cDNA通過影響細胞的轉錄、翻譯及蛋白表達變化從而導致腫瘤的發生。MtDNA具有更加脆弱、突變率高的特點,因而,在近年來,突變的mtDNA與腫瘤的發生關系越來越得到國內外學者的關注。Altafi等也報道了mtDNA D-環區幾乎全是外顯子,長期持續暴露于線粒體高氧濃度的環境中,由于缺乏組蛋白保護和完整的修復系統,mtDNA容易與化學致癌物質結合,造成損傷和突變,同時釋放大量活性氧自由基,從而引起腫瘤的發生。人們在鼻咽癌、大腸癌、肝癌、血液系統等多種惡性腫瘤中,在mtDNA D-Loop均檢測到的突變或多態性變化位點。雖然先前的文獻報道在多種腫瘤細胞或腫瘤組織中發現的mtDNA D-環區的突變,但在具體涉及到腫瘤發生過程中的可能機制則為少見。NIH3T3是研究惡性轉化的靶細胞,當腫瘤細胞中提取的DNA導入到該細胞中,能夠通過將其惡性轉化,從而形成惡性轉化灶。在本研究中,為探討突變的mtDNA D環在腫瘤發生過程中的初步機制,我們通過選取NIH3T3細胞,將突變的肺癌A549、H226細胞的mtDNA的D環區導入其中,通過研究其在細胞中的整合、被轉染的NIH3T3細胞染色體核型、增殖率及凋亡率的變化,以期初步探明mtDNA在腫瘤發生演變中的作用。
表2 轉染前后小鼠胚胎成纖維細胞NIH3T3的570 nm處吸光度變化/±s
有研究報道,在物理、化學及某些生物因素的作用下,mtDNA常會發生缺失和重組,而線粒體膜也可發生破碎、裂解,從而導致胞質中的mtDNA碎片的形成。細胞質中含有大量游離mtDNA及其片段,核酸降解酶活性高,線粒體RNA反轉錄多。此外,在細胞核中還有核孔和DNA連接酶,當細胞質中DNA酶和DNA酶樣物質的數量和質量受到影響時,游離mtDNA或mtDNA片段有機會整合到核DNA中,像一些腫瘤病毒一樣,這些核孔會隨機整合到核DNA中。Ju發現突變的mtDNA片斷隨機整合入核基因組可能在腫瘤發生過程中起重要作用。本研究發現,通過FISH法檢測到外源性的突變的mtDNA能夠隨機整合到NIH3T3細胞的核基因組內,引起導致NIH3T3細胞染色體核型發生了突變和斷裂,呈多倍體表現。染色體核型部分缺失或者異位異常,可能激活或滅活該部位基因的功能的改變,常引起一些染色體異常相關性疾病。Ardern-Holmes等研究發現2型神經纖維瘤病(NF2)是由于22q12號染色體上NF2基因的突變而形成了中樞神經系統腫瘤。但染色體核型異常與腫瘤的發生關系尚不明確。本研究進一步研究發現外源性突變mtDNA誘導NIH3T3細胞的生物學行為發生變化,具有惡性轉化的傾向。主要體現在空NIH3T3和轉染了pcDNA3.1(+)-A549 mtDNA、pcDNA3.1(+)-H226 mtDNA的NIH3T3細胞的增殖率升高,而凋亡率則下降。
肺癌細胞突變的mtDNA導入NIH3T3細胞能夠導致其生物學行為發生一系列改變,初步具有惡性傾向,表明高突變肺癌mtDNA D環區可誘導NIH3T3細胞惡性轉化。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
保健醫苑(2023年2期)2023-03-15 09:03:04
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
癌變·畸變·突變(2016年3期)2016-02-27 06:15:34
醫學研究雜志(2015年12期)2015-06-10 06:57:46
鄭州大學學報(醫學版)(2015年1期)2015-02-27 14:50:26
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
