黃綿馬酸BB對皮膚軟組織感染致病菌的抗菌活性及機制研究
2021-03-17 00:08:46殷川平陳文浩莫梓童吳小詩唐春萍沈志濱
中草藥 2021年6期
殷川平,陳文浩,莫梓童,吳小詩,唐春萍, 2,沈志濱, 2, 3*
黃綿馬酸BB對皮膚軟組織感染致病菌的抗菌活性及機制研究
殷川平1,陳文浩1,莫梓童1,吳小詩1,唐春萍1, 2,沈志濱1, 2, 3*
1. 廣東藥科大學中藥學院,廣東 廣州 511400 2. 廣東省局部精準藥物遞藥制劑工程技術研究中心,廣東 廣州 510006 3. 廣東省化妝品工程技術研究中心,廣東 廣州 510006
研究黃綿馬酸BB對皮膚軟組織感染致病菌的抗菌活性,并探討其對金黃色葡萄球菌的抗菌機制。采用微量稀釋法測定黃綿馬酸BB對皮膚軟組織感染相關致病菌的最低抑菌濃度(minimum inhibitory concentration,MIC)和最低殺菌濃度(minimum bactericidal concentration,MBC);以對黃綿馬酸BB敏感的耐甲氧西林金黃色葡萄球菌(methicillin-resistant,MRSA)臨床分離株和金黃色葡萄球菌標準株ATCC29213為受試菌,考察黃綿馬酸BB對其生長活力和菌體形態(tài)結(jié)構的影響,采用酶標儀考察黃綿馬酸BB對其細胞壁通透性的影響,采用熒光分光光度計考察黃綿馬酸BB對細胞膜電位的影響,采用qRT-PCR法考察黃綿馬酸BB對青霉素結(jié)合蛋白1(penicillin-binding protein 1,)mRNA表達的影響。黃綿馬酸BB對ATCC29213的MIC為10.0~20.0 μg/mL,MBC為10.0~40.0 μg/mL,且對MRSA臨床分離株的MIC小于頭孢唑啉;與對照組比較,黃綿馬酸BB組ATCC29213和MRSA生長受到抑制,菌體細胞形態(tài)結(jié)構改變,細胞膜電位顯著降低(<0.05),1/2MIC黃綿馬酸BB可顯著下調(diào)mRNA表達水平(<0.01),2MIC黃綿馬酸BB可顯著上調(diào)mRNA表達水平(<0.01)。黃綿馬酸BB對MRSA臨床分離株表現(xiàn)出良好的抗菌活性,且其抗菌作用與破壞菌體細胞結(jié)構、降低細胞膜電位、影響mRNA表達有關。
黃綿馬酸BB;耐甲氧西林金黃色葡萄球菌;細胞膜電位;抗菌;青霉素結(jié)合蛋白1
皮膚軟組織感染(skin and soft tissue infection,SSTI)是由病原菌侵犯皮膚表皮、真皮及皮下組織等引起的常見感染性疾病,可誘發(fā)菌血癥、敗血癥,危及患者生命[1-2]。金黃色葡萄球菌和大腸埃希菌為引起SSTI的主要致病菌[3]。在金黃色葡萄球菌引起的SSTI患者中,耐甲氧西林金黃色葡萄球菌(methicillin-resistant,MRSA)的檢出率高達73.21%[4]。隨著抗生素的廣泛應用,SSTI致病菌對臨床中的常用藥物如頭孢唑林、達托霉素均出現(xiàn)了不同程度的耐藥性[5-6],且萬古霉素對MRSA的敏感性逐漸降低[7],萬古霉素中介耐藥的金黃色葡萄球菌檢出率逐漸升高[8]。因此,從傳統(tǒng)中藥中尋找對耐藥菌株具有較好抗菌活性的化合物迫在眉睫。
香鱗毛蕨(L.) Schott為鱗毛蕨科鱗毛蕨屬植物,主要分布于黑龍江省五大連池等地區(qū),常用于治療牛皮癬、痤瘡等病菌感染性皮膚病[9-10]。香鱗毛蕨中的間苯三酚類化合物具有良好的抗菌活性[11],黃綿馬酸BB作為其中的典型間苯三酚類化合物,其抗菌活性及機制尚未明確。本研究探究黃綿馬酸BB對SSTI相關致病菌的抗菌活性及機制,為抗SSTI藥物開發(fā)提供依據(jù)。
1 材料
1.1 菌株
金黃色葡萄球菌標準株ATCC29213購自廣東省菌種保存中心;大腸埃希菌標準株ATCC8739由廣東藥科大學基礎學院提供;3株MRSA(MRSA1~3)臨床分離株及3株大腸埃希菌臨床分離株由廣州醫(yī)科大學呼吸病研究所提供。
1.2 藥品與試劑
黃綿馬酸BB(質(zhì)量分數(shù)>95%)由課題組自制;頭孢唑啉(批號N0825A)購自深圳華潤九新藥業(yè)有限公司;萬古霉素(質(zhì)量分數(shù)>97%)購自美國Sigma公司;達托霉素、尼生素(質(zhì)量分數(shù)>97%)購自大連美侖生物技術有限公司;PBS緩沖液(批號AXH44026)購自HyClone公司;二甲基亞砜(DMSO,批號DH105-2)購自鼎國昌盛生物技術有限公司;Gluta固定液(批號L17D11G134782)購自源葉生物科技有限公司;CAMHB營養(yǎng)肉湯培養(yǎng)基(批號3302092)、營養(yǎng)瓊脂培養(yǎng)基(批號1067752)購自廣東環(huán)凱微生物科技有限公司;細胞膜熒光探針DiSC3-5購自阿拉丁生化科技股份有限公司;堿性磷酸酶(alkaline phosphatase,AKP)測試盒購自南京建成生物工程有限公司;RNA提取試劑盒購自北京天恩澤基因科技有限公司;PrimeScript?RT reagent Kit with gDNA Eraser、SYBR?Premix Ex Taq?II(Tli RNaseH Plus)購自寶生物工程(大連)有限公司。
1.3 儀器
iMark酶標儀、IQ5熒光定量PCR儀(美國BIO-RAD公司);SW-CJ-1F超凈工作臺(蘇凈集團安泰公司);Memmert恒溫培養(yǎng)箱(上海一恒科學儀器有限公司);H-7500透射電子顯微鏡(TEM,日本Hitachi公司);RF-5301PC熒光分光光度計(日本島津公司)。
2 方法
2.1 菌液制備
將上述菌株用營養(yǎng)瓊脂培養(yǎng)基培養(yǎng)至對數(shù)生長期,根據(jù)美國臨床實驗室標準協(xié)會(clinical and laboratory standards institute,CLSI)制定的M07-A9方案中微量稀釋法,將對數(shù)生長期的菌落用生理鹽水稀釋,調(diào)節(jié)至0.5個麥氏濁度(1×108CFU/mL),用CAMHB營養(yǎng)肉湯培養(yǎng)基稀釋100倍(1×106CFU/mL)作為接種菌液。
2.2 藥液制備
黃綿馬酸BB溶于DMSO配制成質(zhì)量濃度為16 mg/mL的貯備液;頭孢唑林、萬古霉素和達托霉素分別溶于超純水配制成質(zhì)量濃度為256 mg/mL的貯備液,于4 ℃避光保存。
2.3 黃綿馬酸BB對SSTI相關致病菌的抗菌活性測定
根據(jù)CLSI的M07-A9方案,采用微量稀釋法測定黃綿馬酸BB和陽性對照藥物(頭孢唑林、萬古霉素、達托霉素)的最小抑菌濃度(minimum inhibitory concentration,MIC)。用CAMHB營養(yǎng)肉湯培養(yǎng)基將藥物貯備液進行稀釋,取100 μL藥物貯備液加入96孔板中,再加入100 μL菌液,使黃綿馬酸BB的終質(zhì)量濃度為2.5~1 280.0 μg/mL,頭孢唑林的終質(zhì)量濃度為0.625~5 120.000 μg/mL,萬古霉素和達托霉素的終質(zhì)量濃度為0.25~32.00 μg/mL,對照組只加入菌液不加入藥液。測定MIC后,選擇質(zhì)量濃度≥MIC的菌懸液,吸取20 μL涂于固體培養(yǎng)基上,于37 ℃恒溫培養(yǎng)箱中培養(yǎng)18~24 h后肉眼觀察,以無細菌生長的最低藥物濃度作為最低殺菌濃度(minimum bactericidal concentration,MBC)。
2.4 黃綿馬酸BB對ATCC29213和MRSA1的時間-殺菌曲線的測定
根據(jù)抗菌活性結(jié)果,選用對黃綿馬酸BB敏感的ATCC29213和MRSA1作為受試菌,測定黃綿馬酸BB對其生長的影響。設置對照組及黃綿馬酸BB(1/2MIC、MIC、2MIC)組,各給藥組加入含相應藥物的菌液,對照組只加入菌液不加入藥液,于37 ℃振蕩培養(yǎng),每2小時采用酶標儀測定24 h內(nèi)600 nm處的吸光度()值。以時間為橫坐標,以值為縱坐標,繪制時間-殺菌曲線。
2.5 TEM觀察黃綿馬酸BB對ATCC29213和MRSA1細胞形態(tài)的影響
設置對照組及黃綿馬酸BB(1/2MIC、MIC、2MIC)組,各給藥組加入含相應藥物的菌液,對照組只加入菌液不加入藥液,于37 ℃振蕩培養(yǎng),分別在菌種對數(shù)生長期(4 h)和生長穩(wěn)定期(18 h)取出菌懸液,4 ℃、4000 r/min離心15 min,收集菌體,PBS緩沖液漂洗3次,棄去上清液,菌體加入2.5% Gluta固定液,于4 ℃固定10 h,制成TEM樣品,于TEM下觀察[12]。
2.6 黃綿馬酸BB對ATCC29213和MRSA1細胞壁通透性的影響
設置對照組及黃綿馬酸BB(1/2MIC、MIC、2MIC)組,各給藥組加入含相應藥物的菌液,對照組只加入菌液不加入藥液,于37 ℃振蕩培養(yǎng)4 h,經(jīng)0.22 μm濾膜濾過,按照試劑盒說明書測定上清液中AKP活性。
2.7 黃綿馬酸BB對ATCC29213和MRSA1細胞膜電位的影響
課題組前期研究中尼生素對ATCC29213和MRSA1的MIC均為25 μg/mL,設置對照組、黃綿馬酸BB(MIC)組、尼生素(MIC)組,各給藥組加入含相應藥物的菌液,對照組只加入菌液不加入藥液,于37 ℃振蕩培養(yǎng)1 h,加入0.4 μmol/L DiSC3-5,培養(yǎng)30 min。激發(fā)波長為622 nm,發(fā)射波長為670 nm,用熒光分光光度計測定熒光強度。
2.8 黃綿馬酸BB對MRSA1中青霉素結(jié)合蛋白1(penicillin-binding protein 1,PBP1)mRNA表達的影響
是金黃色葡萄球菌細胞分裂的關鍵基因,按照文獻報道方法[13],設置對照組及黃綿馬酸BB(1/2MIC、MIC、2MIC)組,各給藥組加入含相應藥物的菌液,對照組只加入菌液不加入藥液,于37 ℃培養(yǎng)4 h,收集菌體,按照試劑盒說明書提取RNA并合成cDNA,進行qRT-PCR分析。引物序列:16S rRNA上游引物5’-GAGAGAAGGTGGGGATGACGT-3’,下游引物5’-AGGCCCGGGAACGTATTCAC-3’,產(chǎn)物大小為217 bp;上游引物5’-AGAGTCGGTTGGG- GTGAAAT-3’,下游引物5’-ACCAGGTGCTTCTCC- ATCAA-3’,產(chǎn)物大小為178 bp。qRT-PCR程序:95 ℃預變性30 s,95 ℃變性5 s,60 ℃退火30 s,40個循環(huán);55 ℃升溫至95 ℃,每升溫0.5 ℃進行1次讀數(shù)。
2.9 數(shù)據(jù)統(tǒng)計與分析
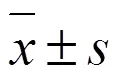
3 結(jié)果
3.1 黃綿馬酸BB對SSTI相關致病菌抗菌活性
如表1所示,頭孢唑啉對3株MRSA臨床分離株的MIC為320.000~640.000 μg/mL,對3株大腸埃希菌臨床分離株的MIC為80.000~1 280.000 μg/mL,CLSI頭孢唑林對葡萄球菌屬和腸桿菌科的耐藥標準分別為MIC≥32.000 μg/mL及MIC≥8.000 μg/mL,表明頭孢唑林對MRSA和大腸埃希菌表現(xiàn)為高度耐藥。萬古霉素對3株MRSA臨床株的MIC為3.54~10.00 μg/mL,對3株大腸埃希菌臨床分離株的MIC>32.00 μg/mL;達托霉素對3株MRSA臨床株的MIC均為2.00 μg/mL;黃綿馬酸BB對3株MRSA臨床分離株的MIC小于陽性對照藥物頭孢唑啉,均為20.0 μg/mL,其MBC與MIC相近,為20.0~40.0 μg/mL,表明黃綿馬酸BB對MRSA臨床株具有較好的抑菌和殺菌作用;3株大腸埃希菌臨床株對黃綿馬酸BB的敏感性較差,MIC和MBC均大于1 280.0 μg/mL,與文獻報道[14]一致。

表1 黃綿馬酸BB對受試菌株的MIC和MBC
3.2 黃綿馬酸BB對ATCC29213和MRSA1的時間-殺菌曲線
如圖1所示,對照組菌株接種2 h后開始出現(xiàn)對數(shù)生長,1/2MIC、MIC黃綿馬酸BB組菌株對數(shù)生長期為第6小時和第14小時,2MIC黃綿馬酸BB組菌株24 h內(nèi)未見生長。表明黃綿馬酸BB抑制ATCC29213和MRSA1生長,呈劑量相關性。
3.3 黃綿馬酸BB對ATCC29213和MRSA1細胞形態(tài)的影響
如圖2所示,對照組ATCC29213和MRSA1細胞形態(tài)結(jié)構完好,壁-膜結(jié)構完整,原生質(zhì)體分布均勻;與對照組比較,MIC黃綿馬酸BB處理至對數(shù)生長期(4 h),細菌形態(tài)皺縮,細胞質(zhì)不均勻,部分壁-膜區(qū)域變得模糊;MIC黃綿馬酸BB處理至生長穩(wěn)定期(18 h),細胞內(nèi)部結(jié)構異常化,染色體固縮,細胞內(nèi)出現(xiàn)大面積的低電子密度空白區(qū)。
3.4 黃綿馬酸BB對ATCC29213和MRSA1細胞壁通透性的影響
如圖3所示,與對照組比較,黃綿馬酸BB組細菌培養(yǎng)液中AKP活性無明顯變化,表明黃綿馬酸BB對ATCC29213和MRSA1細胞壁的通透性無明顯影響。

圖1 黃綿馬酸BB對ATCC29213 (A) 和MRSA1 (B) 的時間-殺菌曲線

圖2 黃綿馬酸BB對ATCC29213和MRSA1細胞形態(tài)的影響(×150 000)

圖3 黃綿馬酸BB對ATCC29213 (A) 和MRSA1 (B) 細胞壁通透性的影響
3.5 黃綿馬酸BB對ATCC29213和MRSA1細胞膜電位的影響
當細胞膜電位降低時,DiSC3-5熒光染料釋放到胞外,熒光強度上升。如表2所示,與對照組比較,MIC黃綿馬酸BB組ATCC29213和MRSA1的DiSC3-5熒光強度明顯提高(<0.05),表明黃綿馬酸BB能夠引起ATCC29213和MRSA1細胞膜電位的降低。

表2 黃綿馬酸BB對ATCC29213和MRSA1細胞膜電位的影響
與對照組比較:*<0.05***<0.001
*< 0.05***< 0.001control group
3.6 黃綿馬酸BB對MRSA1中PBP1 mRNA表達的影響
如圖4所示,與對照組比較,1/2MIC黃綿馬酸BB組MRSA1中mRNA表達水平顯著降低(<0.01),2MIC黃綿馬酸BB組MRSA1中mRNA表達水平顯著升高(<0.01)。
4 討論
SSTI為局限性感染疾病,隨著抗菌藥物的濫用,致病菌產(chǎn)生耐藥性,臨床常用的抗菌藥物治療效果降低[15-16]。黃綿馬酸BB抗菌活性良好[16],對皮膚癬菌有較好的抑制作用[17],但對SSTI致病菌的抗菌活性尚不明確。CLSI規(guī)定金黃色葡萄球菌對萬古霉素的MIC≤2 μg/mL即為敏感,MIC 4~8 μg/mL即為中介,本研究結(jié)果顯示,3株MRSA臨床分離株對萬古霉素的敏感性均下降,呈中介耐藥性。當藥物對微生物的MBC/MIC≥32時,可判定微生物對受試藥產(chǎn)生了耐受性[18],本研究結(jié)果顯示,黃綿馬酸BB對3株MRSA臨床分離株的MBC/MIC≤2,表明黃綿馬酸BB對萬古霉素中介耐藥的MRSA臨床分離株具有顯著的敏感性。

與對照組比較:**P<0.01
完整的菌體細胞結(jié)構是維持生命的必要條件,MIC黃綿馬酸BB能夠破壞菌體細胞壁-膜結(jié)構的完整性,造成染色體固縮、細胞空化。細胞膜電位對三磷酸腺苷(adenosine triphosphate,ATP)的產(chǎn)生和細胞功能具有重要意義[19],MIC黃綿馬酸BB可顯著降低ATCC29213和MRSA1臨床分離株細胞膜電位,從而影響ATP生成、干擾細菌的正常代謝。PBP1是MRSA細胞分裂的關鍵基因,敲除PBP1后,菌株停止分裂[20],本研究發(fā)現(xiàn),1/2MIC黃綿馬酸BB能顯著下調(diào)mRNA表達水平,從而抑制菌體的分裂,阻礙細菌的正常繁殖。2MIC黃綿馬酸BB能顯著上調(diào)mRNA表達水平,黃綿馬酸BB能使MRSA的染色體發(fā)生固縮。研究發(fā)現(xiàn),MRSA的DNA受損引發(fā)的反應會使PBP1表達代償性增加[21],表明2MIC黃綿馬酸BB能夠造成MRSA的DNA的損傷,從而上調(diào)mRNA表達水平。
綜上所述,本研究以造成SSTI的主要病原菌金黃色葡萄球菌和大腸埃希菌為研究對象,闡述了黃綿馬酸BB抗菌活性及機制,為抗SSTI新藥開發(fā)提供理論基礎。細菌的耐藥性與其生物被膜的形成密切相關[22],課題組后續(xù)將對黃綿馬酸BB抑制生物被膜的作用及機制進行深入探究。
利益沖突 所有作者均聲明不存在利益沖突
[1] Salatin S, Lotfipour F, Jelvehgari M. A brief overview on nano-sized materials used in the topical treatment of skin and soft tissue bacterial infections [J]., 2019, 16(12): 1313-1331.
[2] 劉韜滔, 劉亞林, 何清, 等. 軟組織感染致膿毒性休克的早期診斷與治療 [J]. 中華急診醫(yī)學雜志, 2020, 29(1): 76-81.
[3] 柯曉蘋, 蔡良奇, 林維嘉. 皮膚軟組織感染人群病原菌分布與其耐藥性分析 [J]. 中國醫(yī)院藥學雜志, 2019, 39(20): 2073-2076.
[4] 萬東芳, 萬永山, 朱紅軍, 等. 皮膚軟組織感染患者金黃色葡萄球菌感染狀況研究 [J]. 中華醫(yī)院感染學雜志, 2019, 29(1): 32-35.
[5] 陳雪琴. 589株社區(qū)獲得性皮膚軟組織感染病原菌分布及耐藥性分析 [D]. 遵義: 遵義醫(yī)科大學, 2019.
[6] 廖國建, 彭希希, 田俊, 等. 達托霉素耐藥分子機制研究進展 [J]. 生物工程學報, 2018, 34(6): 832-838.
[7] Hu H C, Kao K C, Chiu L C,. Clinical outcomes and molecular typing of heterogenous vancomycin- intermediatebacteremia in patients in intensive care units [J]., 2015, 15: 444.
[8] Chung D R, Lee C, Kang Y R,. Genotype-specific prevalence of heterogeneous vancomycin-intermediatein Asian countries [J]., 2015, 46(3): 338-341.
[9] 陳玲玲, 梁彥濤, 卜志剛, 等. 黑龍江省香鱗毛蕨六個天然分布地種群結(jié)構分析 [J]. 北方園藝, 2016, 40(24): 70-76.
[10] 朱沖沖, 彭冰, 曾祖平, 等. 香鱗毛蕨的化學成分及藥理作用研究進展 [J]. 中國藥房, 2017, 28(10): 1418-1423.
[11] 康廷國. 中藥鑒定學 [M]. 北京: 中國中醫(yī)藥出版社, 2003: 115.
[12] 謝強, 林玉桓, 苗淑萍, 等. 香芹酚對大腸桿菌和金黃色葡萄球菌細胞膜的影響 [J]. 食品工業(yè)科技, 2014, 35(23): 54-58.
[13] 樊莉, 孫鳳軍, 枉前, 等. 亞抑菌濃度抗菌藥物對多重耐藥鮑曼不動桿菌生物膜形成的影響 [J]. 中國藥房, 2018, 29(22): 3129-3134.
[14] 李曉娟. 香鱗毛蕨精油和間苯三酚類成分提取分離及其抑菌活性研究 [D]. 哈爾濱: 東北林業(yè)大學, 2012.
[15] 鄭躍, 席云, 賴維, 等. 皮膚軟組織感染致病菌的菌種構成和藥敏變化趨勢分析 [J]. 皮膚性病診療學雜志, 2014, 21(1): 29-33.
[16] 王敏, 原明明, 李樹君, 等. 成人細菌性皮膚病的病原菌分布與耐藥性研究 [J]. 中國病原生物學雜志, 2020, 15(7): 846-849.
[17] 梁玉婷, 宋國強, 林楚怡, 等. 香鱗毛蕨化學成分及其體外抗淺部真菌活性研究 [J]. 天然產(chǎn)物研究與開發(fā), 2019, 31(10): 1758-1763.
[18] 周本宏, 李妍, 周夢宇, 等. 消炎抗菌片的抗菌活性及對福氏志賀氏菌的抗菌機制初步探討 [J]. 中國醫(yī)院藥學雜志, 2018, 38(23): 2429-2433.
[19] 劉蓉, 唐方, 凌海慧, 等. 紫蘇葉油對結(jié)腸平滑肌細胞鈣離子和膜電位的作用 [J]. 時珍國醫(yī)國藥, 2019, 30(6): 1302-1304.
[20] Pereira S F, Henriques A O, Pinho M G,. Evidence for a dual role of PBP1in the cell division and cell separation of[J]., 2009, 72(4): 895-904.
[21] Plata K B, Riosa S, Singh C R,. Targeting of PBP1by β-lactams determines recA/SOS response activation in heterogeneous MRSA clinical strains [J]., 2013, 8(4): e61083.
[22] 王洪彬, 朱利霞, 于秀劍, 等. 細菌生物被膜研究進展 [J]. 動物醫(yī)學進展, 2019, 40(9): 74-79.
Antibacterial activity and mechanism of flavaspidic acid BB on pathogenic bacteria of skin and soft tissue infection
YIN Chuan-ping1, CHEN Wen-hao1, MO Zi-tong1, WU Xiao-shi1, TANG Chun-ping1, 2, SHEN Zhi-bin1, 2, 3
1. School of Traditional Chinese Medicines, Guangdong Pharmaceutical University, Guangzhou 511400, China 2. Guangdong Provincial Engineering Center of Topical Precise Drug Delivery System, Guangzhou 510006, China 3. Guangdong Provincial Cosmetics Engineering Technology Research Center, Guangzhou 510006, China
To study the antibacterial activity of flavaspidic acid BB on pathogenic bacteria related to skin and soft tissue infections, and explore the antibacterial mechanism against.The microdilution method was used to determine the minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of flavaspidic acid BB against pathogenic bacteria related to skin and soft tissue infections; Clinical strain of methicillin-resistant(MRSA) andstandard strain ATCC29213, which were sensitive to flavaspidic acid BB, were used as test bacteria to explore the influence of flavaspidic acid BB on growth vitality and the morphological structure of tested bacteria. Effect of flavaspidic acid BB on permeability of cell wall was detected by microplate reader; Effect of flavaspidic acid BB on cell membrane potential was detected by fluorescence spectrophotometer; Effect of flavaspidic acid BB on expression of penicillin-binding protein 1 () mRNA was detected by qRT-PCR.MIC of flavaspidic acid BB against ATCC29213 was 10.0—20.0 μg/mL, MBC was 10.0—40.0 μg/mL, MIC values of MRSA clinical strains were lower than cefazolin. Compared with control group, growth of ATCC29213 and MRSA in flavaspidic acid BB group were inhibited, morphological structure of bacterial cell was changed, and cell membrane potential was reduced (< 0.05), expression ofmRNA was down-regulated by 1/2MIC flavaspidic acid BB and up-regulated by 2MIC flavaspidic acid BB (< 0.01).The flavaspidic acid BB exhibits good antibacterial activity against MRSA clinical strains, of which antibacterial effect is related to the destruction of bacterial cell structure, lowering the cell membrane potential and affecting the expression ofmRNA.
flavaspidic acid BB; methicillin-resistant; cell membrane potential; antibacterial activity; penicillin-binding protein 1
R285.5
A
0253 - 2670(2021)06 - 1662 - 06
10.7501/j.issn.0253-2670.2021.06.014
2020-10-25
國家重點研發(fā)計劃“中醫(yī)藥現(xiàn)代化”重點專項項目(2018YFC1707100);廣東省科技廳應用型研發(fā)專項(2015B020234009)
殷川平(1995—),男,碩士研究生,研究方向為天然藥物化學。E-mail: 2274609080@qq.com
沈志濱(1964—),女,教授,博士,研究方向為中藥藥效物質(zhì)基礎及新藥研發(fā)。E-mail: szb8113@126.com
[責任編輯 李亞楠]
