E3泛素連接酶TRIM50促進口腔鱗狀細(xì)胞癌增殖及遷移侵襲的分子機制研究
2021-03-16 10:24:16趙許兵馬志軍
山西醫(yī)科大學(xué)學(xué)報 2021年2期
劉 琛,趙許兵,程 政,馬志軍
(1西安交通大學(xué)口腔醫(yī)學(xué)院口腔綜合科,西安 710004;2西安市第九醫(yī)院口腔科;*通訊作者,E-mail:Maazhijun@126.com)
口腔鱗狀細(xì)胞癌(oral squamous cell carcinoma,OSCC)是最常見的口腔頜面部惡性腫瘤,在口腔癌中占比90%,其發(fā)病率在全身腫瘤中排名第八[1]。盡管針對OSCC的放化療以及手術(shù)治療在不斷進步,但是患者的總生存率并沒有因此而得到明顯的提高[2,3]。因此,尋找可靠的生物標(biāo)志物和新的分子靶點對為OSCC患者制定并實施精準(zhǔn)的個性化的治療方案至關(guān)重要[4]。
泛素化是泛素與目標(biāo)蛋白賴氨酸殘基發(fā)生共價結(jié)合使其降解的生物學(xué)過程,此過程由3種酶完成,泛素激活酶(E1)、泛素偶聯(lián)酶(E2)和泛素連接酶(E3),每一種酶都由一組蛋白組成[5]。其中E3連接酶是泛素級聯(lián)的關(guān)鍵組成部分,通過直接與各種底物相互作用,在泛素化途徑中發(fā)揮特殊作用[6,7]。目前,根據(jù)泛素蛋白酶體系統(tǒng)(ubiquitin proteasome system,UPS)的組成成分進而開發(fā)靶標(biāo)藥物是腫瘤研究中的一個熱點,受到越來越多的關(guān)注[8]。TRIM(tripartite motif containing protein)蛋白家族近年來因其特有的結(jié)構(gòu)(3個保守的結(jié)構(gòu)域)和廣泛參與各種生物學(xué)途徑而備受關(guān)注[9]。越來越多的證據(jù)表明部分TRIM家族成員作為E3泛素連接酶介導(dǎo)泛素化降解途徑靶向調(diào)控癌相關(guān)基因,在癌癥發(fā)展的過程中起到關(guān)鍵作用[10]。
TRIM50作為TRIM蛋白家族新成員,參與調(diào)控神經(jīng)系統(tǒng)的發(fā)育以及胃消化腺的分泌等過程[11,12]。但其在腫瘤方面的相關(guān)報道甚少,特別是在口腔癌中并沒有報道。本研究旨在探究E3泛素連接酶TRIM50對OSCC生物學(xué)功能的影響,并初步探究TRIM50蛋白表達對OSCC細(xì)胞惡性表型調(diào)控的潛在機制,為明晰OSCC發(fā)病機制以及發(fā)展分子靶向治療提供理論基礎(chǔ)。
1 材料與方法
1.1 主要試劑

1.2 實驗方法
1.2.1 細(xì)胞培養(yǎng) OSCC細(xì)胞系SCC9、SCC25以及Cal27購于美國菌種保藏中心,HOK和HSC3細(xì)胞購于廣州賽庫生物公司。細(xì)胞完全培養(yǎng)基按DMEM培養(yǎng)基 ∶胎牛血清 ∶青鏈霉素=90 ∶9 ∶1的體積比例配制,細(xì)胞培養(yǎng)瓶放置在37 ℃、體積分?jǐn)?shù)為5% CO2的恒溫培養(yǎng)箱中培養(yǎng)。當(dāng)OSCC細(xì)胞生長至密度為約80%時,用0.25%的胰蛋白酶消化并傳代,收集生長狀態(tài)良好的細(xì)胞進行后續(xù)的細(xì)胞轉(zhuǎn)染、細(xì)胞增殖、克隆形成以及遷移侵襲等實驗。
1.2.2 過表達載體構(gòu)建 過表達目的蛋白的載體為pcDNA3.1(+),由CMV啟動子驅(qū)動目的基因的表達。TRIM50-Flag cDNA克隆的上游引物序列為5′-GCGAAGCTTATGGCTTGGCAGGTGAGCCTG-3′,其5′端包含了保護堿基GCG以及HindⅢ限制性內(nèi)切酶的酶切位點;其下游引物序列為5′-CGGGATCCCTTGTCATCGTCGTCCTTGTAGTCCAGCTTG-GTGGGCTGCTCGG-3′,其5′端包含了BamHⅠ限制性內(nèi)切酶的酶切位點。Rb-HA cDNA克隆的上游引物序列為5′-GCGAAGCTTGCGGAAAGGCGTCA-TGCCGCC-3′,下游引物的序列為5′-CGGGATCCAGCGTAATCTGGAACATCGTATGGGTATTTC TCTTCCTTGTTTGAGGTATCC-3′。其上游及下游引物的5′端分別包含HindⅢ以及BamHⅠ酶切位點。Ub-Flag cDNA克隆上游引物序列為5′-GCGAAGC-TTATGCAGATCTTCGTGAAAACCC-3′,下游引物序列為5′-CGGGATCCCTTGTCATCGTCG TCCTTGTAGTCGCCACCCCTCAGGCGCAGG-3′。全長cDNA克隆PCR的反應(yīng)條件為:94 ℃預(yù)變性5 min;94 ℃變性30 s,58 ℃退火30 s,72 ℃延伸按每個千道爾頓1 min進行,循環(huán)33次。
1.2.3 細(xì)胞轉(zhuǎn)染 按1×105的數(shù)量將OSCC細(xì)胞接種于6孔板中。待細(xì)胞融合至密度約為60%時,按照LipofectamineTM2000試劑盒說明書,轉(zhuǎn)染TRIM50 siRNAs以及上述過表達載體和空白載體pcDNA3.1(+)至OSCC細(xì)胞。轉(zhuǎn)染6 h后,更換新鮮培養(yǎng)基并繼續(xù)培養(yǎng)48 h,收集目的細(xì)胞用于后續(xù)實驗。本實驗所用干擾片段均由吉瑪基因公司(上海)設(shè)計并合成,具體序列見表1。

表1 干擾片段序列
1.2.4 采用蛋白質(zhì)免疫印跡法檢測TRIM50和Rb蛋白表達量 收集轉(zhuǎn)染后的各組OSCC細(xì)胞,加入RIPA裂解液,置于冰上充分裂解30 min以提取細(xì)胞的總蛋白。用BCA蛋白試劑盒測定各組蛋白濃度后,取適量蛋白溶液,95 ℃加熱15 min使蛋白質(zhì)變性。每泳道上樣20 μg蛋白,之后進行十二烷基硫酸鈉-聚丙烯酰胺凝膠(dodecyl sulfate sodium salt-polyacrylamide gel electrophoresis,SDS-PAGE)電泳(100 V,90 min),將分離的蛋白通過半干轉(zhuǎn)法(100 V,45 min)轉(zhuǎn)移至聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF),轉(zhuǎn)膜后置于5%脫脂奶粉溶液中封閉2 h,再分別加入抗TRIM50(ab272586,abcam,1 ∶1 000)、β-actin(BS6007M,Bioworld,1 ∶5 000)、Rb(ab24,abcam,1 ∶1 000)、Flag(M185-3L,MBL,1 ∶2 500)和HA(M180-3S,MBL,1 ∶2 000)抗體,4 ℃孵育過夜。加入辣根過氧化物酶(horseradish peroxidase,HRP)標(biāo)記的二抗(1 ∶1 000),37 ℃下孵育1 h。洗膜后,滴加增強化學(xué)發(fā)光液進行避光顯影。
1.2.5 采用逆轉(zhuǎn)錄聚合酶鏈反應(yīng)法檢測TRIM50和Rb基因表達量 使用TRIzol裂解液裂解實驗細(xì)胞并提取總RNA,然后通過逆轉(zhuǎn)錄試劑盒將RNA逆轉(zhuǎn)錄為cDNA。再以cDNA為模板,通過聚合酶鏈?zhǔn)椒磻?yīng)(polymerase chain reaction,PCR)試劑盒進行PCR擴增。TRIM50上游引物:5′-CGCCAAACTGGTGAACAAC-3′;下游引物:5′-TGGTCCTCATTGCCGAAC-3′。Rb上游引物:5′-AGTTCGCTTGTATTACCG-3′;下游引物:5′-ATTTCAATGGCTTCTGG-3′。內(nèi)參基因GAPDH上游引物:5′-CAA-GGTCATCCATGACAACTTTG-3′;下游引物:5′-GT-CCACCACCCTGTTGCTGTAG-3′。擴增結(jié)束后,進行瓊脂糖凝膠電泳。
1.2.6 采用細(xì)胞計數(shù)法檢測細(xì)胞增殖能力 用靶向TRIM50的siRNAs或過表達質(zhì)粒轉(zhuǎn)染OSCC細(xì)胞(1×104)48 h后,將各組細(xì)胞接種于24孔培養(yǎng)板中培養(yǎng)。分別在細(xì)胞培養(yǎng)24,48,72,96,120 h后進行細(xì)胞計數(shù),每次實驗重復(fù)3次,統(tǒng)計各組的細(xì)胞數(shù),并繪制細(xì)胞生長曲線圖。
1.2.7 采用細(xì)胞克隆集落形成法檢測細(xì)胞克隆形成能力 同樣將所示的質(zhì)粒或siRNAs轉(zhuǎn)染至OSCC細(xì)胞48 h后,將其置于6孔培養(yǎng)板中,用含10%胎牛血清的DMEM培養(yǎng)基培養(yǎng)14 d。之后用無水甲醇固定細(xì)胞,然后用結(jié)晶紫溶液染色。在顯微鏡下統(tǒng)計≥30 μm的細(xì)胞集落數(shù),每次實驗重復(fù)3次。
1.2.8 采用Transwell法檢測細(xì)胞遷移侵襲能力 Transwell小室實驗用于體外細(xì)胞遷移和侵襲的測定,用siRNAs或是質(zhì)粒轉(zhuǎn)染OSCC細(xì)胞。之后,將無血清培養(yǎng)基重懸1×104轉(zhuǎn)染后的細(xì)胞并接種到上室進行細(xì)胞遷移測定。對細(xì)胞侵襲的測定,我們在上室覆蓋基質(zhì)膠再接種相應(yīng)數(shù)量的轉(zhuǎn)染細(xì)胞。遷移和侵襲后的細(xì)胞用無水甲醇固定,通過5%結(jié)晶紫溶液染色。在顯微鏡下從每個下室膜中采集圖像,統(tǒng)計發(fā)生遷移侵襲的細(xì)胞數(shù)量。
1.2.9 采用免疫共沉淀法驗證TRIM50蛋白與Rb蛋白的相互作用 將TRIM50-Flag和Rb-HA過表達質(zhì)粒轉(zhuǎn)染至HSC3細(xì)胞48 h,再用10 μmol/L MG132處理細(xì)胞6 h,然后使用抗Flag或HA抗體孵育轉(zhuǎn)染細(xì)胞的蛋白裂解液,4 ℃過夜,使其充分與目的蛋白結(jié)合,免疫復(fù)合物用蛋白A/G瓊脂糖珠進行捕獲,之后再通過SDS-PAGE電泳分離。然后對電泳后PAGE凝膠進行銀染質(zhì)譜分析,或者通過蛋白質(zhì)免疫印跡法檢測免疫共沉淀復(fù)合物中的蛋白。
1.2.10 采用放線菌酮實驗檢測Rb蛋白降解半衰期的變化 將TRIM50-Flag過表達質(zhì)粒轉(zhuǎn)染至HSC3細(xì)胞,并設(shè)置空載組作為對照,轉(zhuǎn)染48 h后,提取細(xì)胞蛋白記為0 h,然后在培養(yǎng)基中加入20 μg/ml CHX,分別在加入2,4,6,8 h后提取細(xì)胞蛋白。之后,通過蛋白質(zhì)免疫印跡法檢測CHX處理后的細(xì)胞內(nèi)蛋白降解的情況。
1.2.11 采用體外泛素化實驗檢測Rb蛋白的泛素化水平 將TRIM50-Flag、Rb-HA以及Ub-Flag過表達質(zhì)粒共轉(zhuǎn)染HSC3細(xì)胞48 h,MG132(10 μmol/L)處理6 h,使用抗HA抗體免疫沉淀Rb-HA蛋白免疫復(fù)合物,通過蛋白質(zhì)免疫印跡法檢測Rb-HA蛋白的泛素化水平。
1.3 統(tǒng)計學(xué)方法
采用SPSS 18統(tǒng)計學(xué)軟件處理分析實驗數(shù)據(jù),實驗結(jié)果通過GraphPad Prism 5繪圖。所有實驗均獨立重復(fù)至少3次,兩組數(shù)據(jù)采用獨立樣本t檢驗,多組間比較采用方差分析,兩兩比較采用LSD-t檢驗。當(dāng)P<0.05時,視為差異有統(tǒng)計學(xué)意義。
2 結(jié)果
2.1 TRIM50蛋白在不同的OSCC細(xì)胞系中的表達量
蛋白質(zhì)印跡結(jié)果顯示,TRIM50蛋白的表達水平在SCC9和SCC25細(xì)胞中較低,在Cal27與HOK細(xì)胞中較高(見圖1)。

圖1 TRIM50蛋白在不同OSCC細(xì)胞系中的表達
2.2 過表達TRIM50促進OSCC細(xì)胞增殖能力
蛋白質(zhì)印跡結(jié)果顯示,TRIM50-Flag-pcDNA3.1(+)轉(zhuǎn)染的SCC9以及SCC25細(xì)胞后成功過表達TRIM50蛋白(見圖2A)。與空白對照組相比,過表達TRIM50組OSCC細(xì)胞增殖活力明顯增強,差異具有統(tǒng)計學(xué)意義(P<0.01,見圖2B)。

圖2 過表達TRIM50對OSCC細(xì)胞的增殖能力的調(diào)控作用
2.3 過表達TRIM50促進OSCC細(xì)胞克隆形成能力
與空白對照組相比,過表達TRIM50組OSCC細(xì)胞的克隆形成集落數(shù)明顯增多,差異具有統(tǒng)計學(xué)意義(P<0.05,見圖3)。
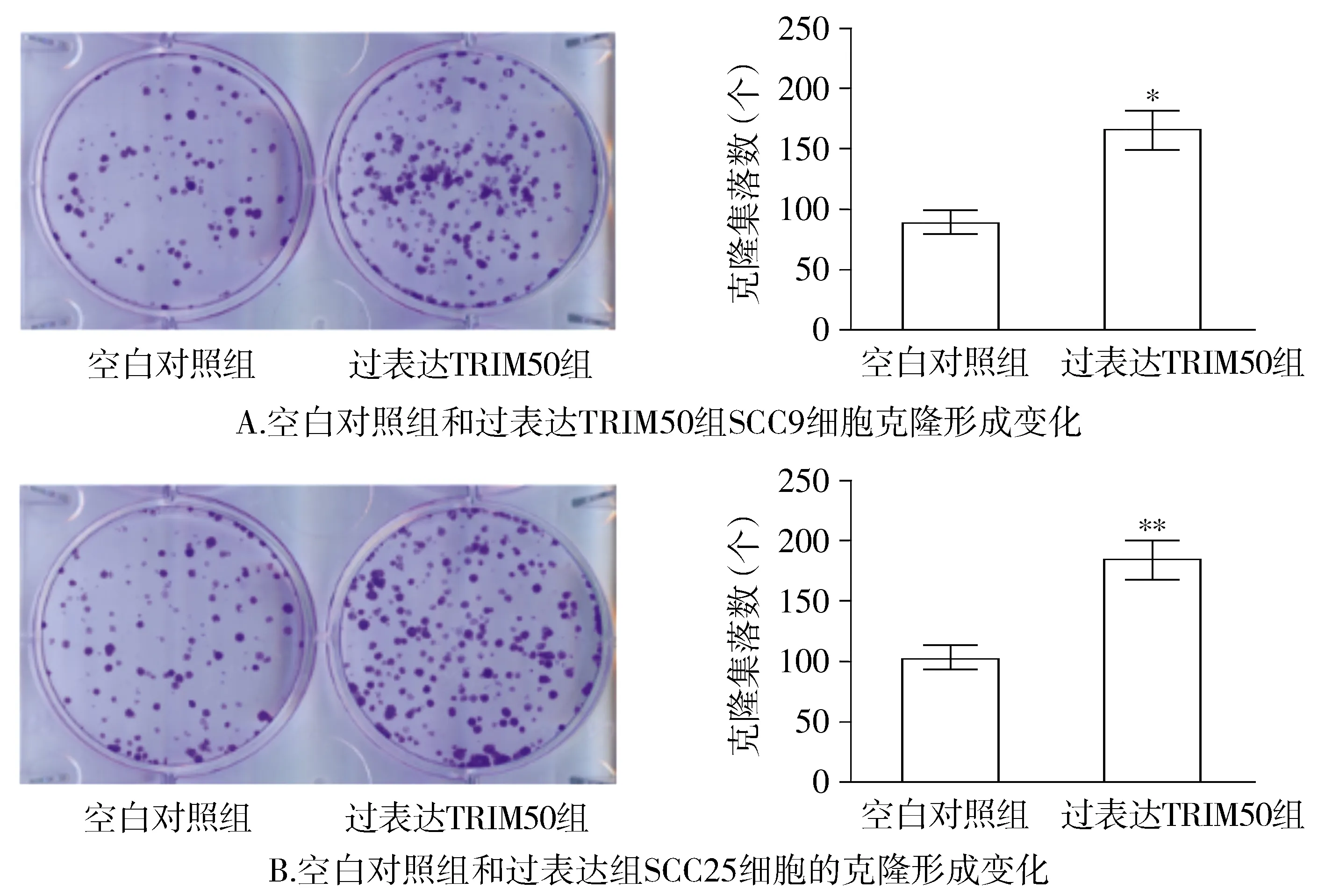
與空白對照組比較,*P<0.05,**P<0.01
2.4 過表達TRIM50促進OSCC細(xì)胞遷移侵襲能力
與空白對照組相比,過表達TRIM50組的OSCC細(xì)胞遷移侵襲的數(shù)量顯著升高(P<0.01,見圖4)。

與空白對照組比較,*P<0.05,**P<0.01
2.5 沉默TRIM50抑制OSCC細(xì)胞增殖能力
蛋白質(zhì)印跡結(jié)果顯示,TRIM50 siRNA干擾片段轉(zhuǎn)染后的Cal27以及HOK細(xì)胞成功下調(diào)TRIM50蛋白的表達(見圖5A)。與陰性對照組相比,沉默TRIM50組OSCC細(xì)胞的生長能力顯著受到抑制(P<0.01,見圖5B)。
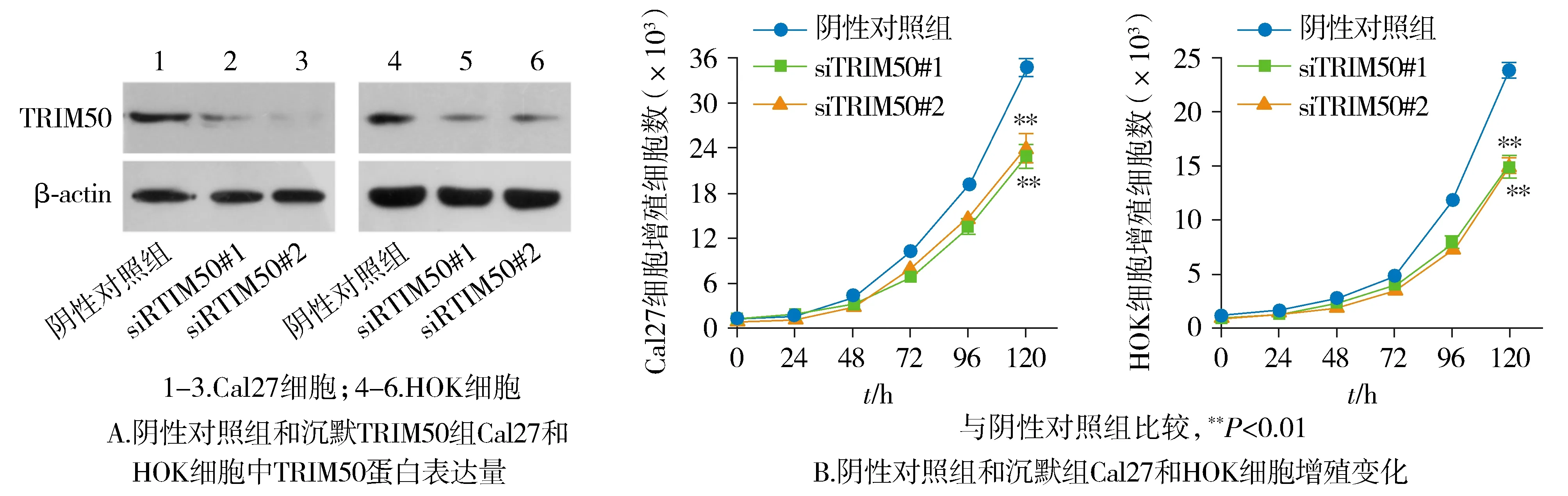
圖5 TRIM50沉默對OSCC細(xì)胞的增殖能力的影響
2.6 沉默TRIM50抑制OSCC細(xì)胞克隆形成能力
與陰性對照組相比,沉默TRIM50組OSCC細(xì)胞克隆形成能力顯著下調(diào)(P<0.01,見圖6)。

與陰性對照組比較,**P<0.01
2.7 沉默TRIM50抑制OSCC細(xì)胞遷移侵襲能力
與陰性對照組相比,沉默TRIM50組OSCC細(xì)胞遷移侵襲的能力受到顯著抑制(P<0.01,見圖7)。
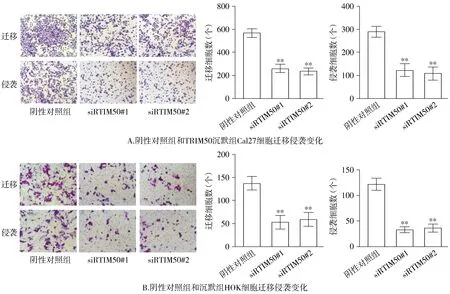
與陰性對照組比較,**P<0.01
2.8 TRIM50與抑癌蛋白Rb相互作用
根據(jù)TRIM50蛋白免疫免疫共沉淀蛋白印跡法結(jié)果顯示,與空白對照組相比,過表達TRIM50組的Rb蛋白存在于TRIM50蛋白復(fù)合物中(見圖8A),過表達Rb組的TRIM50蛋白也存在于Rb蛋白復(fù)合物中(見圖8B),二者共沉淀。

圖8 TRIM50蛋白與抑癌蛋白Rb相互作用
2.9 TRIM50抑制Rb蛋白的表達
蛋白質(zhì)印跡以及逆轉(zhuǎn)錄PCR結(jié)果顯示,與空白對照組相比,過表達TRIM50組Rb蛋白的表達水平下調(diào),Rb mRNA的表達水平?jīng)]有變化(見圖9A)。與陰性對照組相比,而沉默TRIM50組Rb蛋白的表達量上調(diào),Rb mRNA的表達沒有變化(見圖9B)。
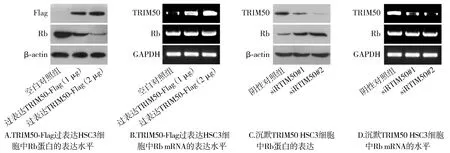
圖9 TRIM50對Rb表達水平的調(diào)控
2.10 TRIM50促進Rb的泛素化降解從而下調(diào)Rb蛋白的表達
相比于空白對照,TRIM50過表達縮短了Rb蛋白降解的半衰期(P<0.05,見圖10A,B)。泛素化實驗結(jié)果顯示,在蛋白酶體抑制劑MG132存在時,相比于空白對照組,TRIM50過表達組Rb多聚泛素化水平增多(見圖10C)。
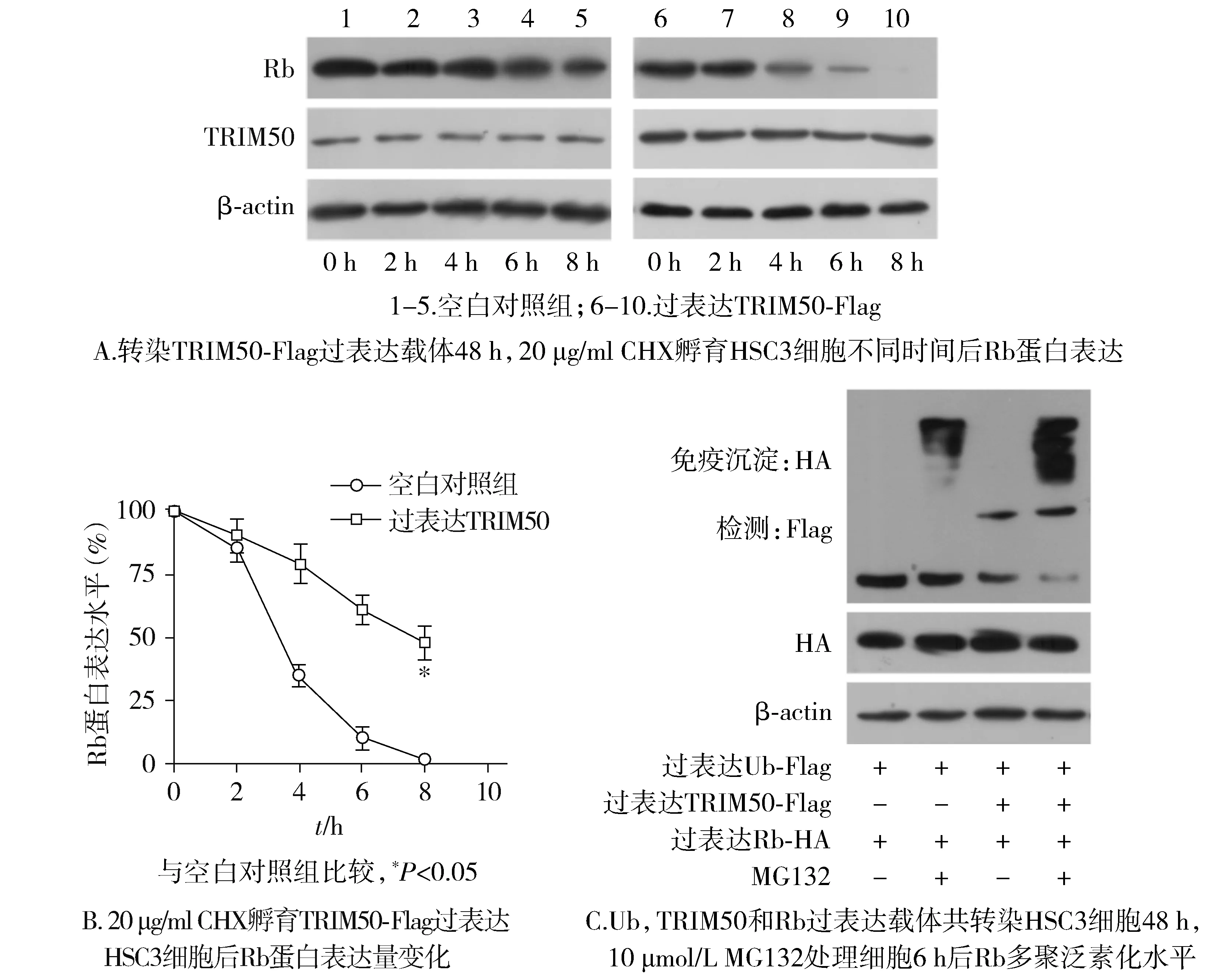
圖10 過表達TRIM50對Rb半衰期以及泛素化水平的影響
2.11 TRIM50表達逆轉(zhuǎn)Rb過表達對HSC3細(xì)胞增殖的抑制作用
蛋白質(zhì)印跡結(jié)果顯示,TRIM50-Flag-pcDNA3.1(+)和Rb-HA-pcDNA3.1(+)共轉(zhuǎn)染的HSC細(xì)胞成功過表達Rb和TRIM50蛋白(見圖11A)。細(xì)胞生長實驗結(jié)果顯示,相比于Rb過表達組,TRIM50與Rb共過表達組HSC細(xì)胞增殖能力增強(P<0.01,見圖11B),但相比于空白對照組,TRIM50與Rb共過表達組HSC細(xì)胞增殖能力沒有變化(P>0.05,見圖11B)。

圖11 TRIM50表達逆轉(zhuǎn)Rb過表達對HSC3細(xì)胞增殖的抑制作用
2.12 TRIM50過表達逆轉(zhuǎn)Rb過表達對HSC3細(xì)胞克隆形成的抑制作用
細(xì)胞克隆形成實驗結(jié)果顯示,相比于Rb過表達組,TRIM50與Rb共過表達組的HSC3細(xì)胞克隆形成能力增強(P<0.01,見圖12);但相比于空白對照組,TRIM50與Rb共過表達組HSC3細(xì)胞克隆形成能力沒有明顯變化(P>0.05,見圖12)。

圖12 TRIM50過表達逆轉(zhuǎn)Rb過表達對HSC3細(xì)胞克隆形成的抑制作用
2.13 TRIM50過表達逆轉(zhuǎn)Rb過表達對HSC3細(xì)胞遷移侵襲的抑制作用
細(xì)胞遷移侵襲實驗結(jié)果顯示,相比于Rb過表達組,TRIM50與Rb共過表達組HSC3細(xì)胞遷移侵襲能力增強,差異具有統(tǒng)計學(xué)意義(P<0.01,見圖13);但相比于空白對照組,TRIM50與Rb共過表達組HSC3細(xì)胞遷移侵襲能力沒有變化(P>0.05,見圖13)。

與其余三組比較,**P<0.01
3 討論
根據(jù)腫瘤統(tǒng)計數(shù)據(jù)顯示,OSCC是頭頸部最常見的腫瘤之一,全球每年有超過30萬的新確診患者[13,14]。由于OSCC具有高侵襲性轉(zhuǎn)移生物學(xué)行為,即使是早期癌也容易出現(xiàn)淋巴結(jié)轉(zhuǎn)移,致死率較高[15]。同時,患者術(shù)后常有復(fù)發(fā),預(yù)后不盡人意,其5年生存率基本保持不變[3]。因此,尋找新的關(guān)鍵靶點分子對OSCC治療具有重要意義。UPS負(fù)責(zé)降解細(xì)胞中80%-90%的已知蛋白[16,17]。人類UPS的特異性主要由大約617個E3泛素連接酶決定,參與細(xì)胞周期調(diào)控和腫瘤發(fā)生的一些蛋白(如p53、p27和cyclins)被E3連接酶酶特異性調(diào)控[18-20]。由此可見,E3泛素連接酶腫瘤發(fā)生發(fā)展中的發(fā)揮著重要作用。
TRIM50最初在Williams-Beuren綜合征中被鑒定為E3泛素連接酶[12]。另有報道表明,TRIM50與SNAIL相互作用,參與調(diào)控其降解抑制肝癌的發(fā)生發(fā)展[21]。到目前為止,有關(guān)TRIM50功能的報道非常有限,其生物學(xué)功能也遠(yuǎn)未闡明。TRIM50在癌變中的作用并不是很明確,尤其是在OSCC中,鑒于E3連接酶在疾病中的活性,我們預(yù)計它可能參與調(diào)控OSCC的發(fā)展。在本研究中,我們在OSCC細(xì)胞中進行TRIM50過表達,并檢測細(xì)胞增殖、克隆形成、遷移侵襲等功能的變化。結(jié)果發(fā)現(xiàn),過表達TRIM50能夠增強OSCC細(xì)胞的增殖、克隆形成以及遷移侵襲能力。另外,TRIM50沉默后相關(guān)的功能實驗結(jié)果顯示,沉默TRIM50抑制了OSCC細(xì)胞的增殖、克隆形成以及遷移侵襲能力。因此,綜合TRIM50過表達和沉默兩方面的功能實驗結(jié)果表明,E3泛素化連接酶TRIM50在OSCC惡性表型維持中呈現(xiàn)出正向調(diào)控的功能,我們首次證明了TRIM50作為一個促癌基因調(diào)控OSCC的發(fā)生發(fā)展。
為了深入研究TRIM50促進OSCC細(xì)胞增殖和轉(zhuǎn)移的分子機制,我們通過免疫共沉淀和質(zhì)譜分析的方法來鑒定與其相互作用的蛋白,以發(fā)現(xiàn)E3連接酶TRIM50作用的底物。根據(jù)質(zhì)譜分析的結(jié)果,我們通過免疫共沉淀蛋白印跡法證實,TRIM50蛋白與Rb蛋白相互作用,說明Rb蛋白可能是E3連接酶TRIM50的潛在底物。眾所周知,視網(wǎng)膜母細(xì)胞瘤腫瘤抑制蛋白Rb是細(xì)胞生長周期和的關(guān)鍵調(diào)節(jié)因子,調(diào)控相關(guān)基因的轉(zhuǎn)錄和表達從而導(dǎo)致細(xì)胞周期在G1期停滯[22]。為了進一步明確TRIM50與Rb的關(guān)系,我們探究了TRIM50的表達對Rb mRNA、蛋白表達水平的影響。結(jié)果發(fā)現(xiàn),TRIM50負(fù)向調(diào)控Rb蛋白的表達量卻不影響其mRNA的表達水平,說明TRIM50在轉(zhuǎn)錄后水平負(fù)向調(diào)控Rb蛋白的表達。因為TRIM50是E3泛素連接酶,因此我們推測TRIM50可能是通過促進了Rb的泛素化降解從而抑制OSCC細(xì)胞中的Rb蛋白的表達。為了確定TRIM50是否與Rb的泛素化降解有關(guān),我們進一步研究了TRIM50對Rb蛋白半衰期和泛素化的影響。放線菌酮實驗結(jié)果顯示,TRIM50過表達縮短了Rb蛋白降解的半衰期。泛素化實驗結(jié)果顯示,過表達TRIM50增加了Rb蛋白多聚泛素化的水平。綜上所述,我們的結(jié)果表明了TRIM50通過促進Rb的泛素化降解從而下調(diào)OSCC細(xì)胞中的Rb蛋白的表達量。為了證實TRIM50通過調(diào)控Rb蛋白的表達量從而抑制OSCC細(xì)胞的增殖和轉(zhuǎn)移,我們進行了TRIM50和Rb共表達組合功能實驗,結(jié)果發(fā)現(xiàn),過表達TRIM50能逆轉(zhuǎn)過表達Rb對OSCC細(xì)胞惡性生物學(xué)功能的抑制作用。因此可以說明,Rb是TRIM50下游的靶標(biāo)分子,被其所介導(dǎo)的泛素化降解途徑所調(diào)控,從而影響OSCC的生物學(xué)功能,我們初步闡明了TRIM50促進OSCC的增殖和轉(zhuǎn)移的分子機制。
綜上所述,我們揭示了TRIM50與OSCC細(xì)胞增殖與轉(zhuǎn)移之間的關(guān)系,發(fā)現(xiàn)E3泛素化連接酶TRIM50在OSCC中作為一個腫瘤驅(qū)動因子調(diào)控細(xì)胞的惡性表型,從而促進了OSCC的發(fā)生發(fā)展。TRIM50通過促進OSCC細(xì)胞中抑癌蛋白Rb的泛素化和降解來間接促進其激進的生物學(xué)行為。因此,靶向TRIM50-Rb通路可能是治療OSCC的有效手段。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
發(fā)明與創(chuàng)新(2022年30期)2022-10-03 08:40:56
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
人大建設(shè)(2018年6期)2018-08-16 07:23:10
文理導(dǎo)航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
