2種白刺對鹽脅迫的代謝響應機制*
2021-03-13 03:16:38閆海冰張慧芳于兆友楊秀清
林業科學 2021年1期
閆海冰 張慧芳 馮 帆 于兆友 楊秀清
(山西農業大學林學院 太谷 030801)
土壤鹽堿化已成為全球性的資源和環境問題,全球約有 9.5億hm2的鹽堿土。中國的鹽堿土面積可達0.36億hm2(潘雄波等,2016),而這些土地已成為農林業發展的障礙用地,并嚴重影響我國的生態環境建設。研究植物的耐鹽堿機制,評價并篩選耐鹽堿植物進行鹽堿地的生物改良,是實現鹽堿地提質增效利用并實現鹽堿地長效生態修復的重要途徑(劉雨欣,2018)。
白刺屬(Nitraria)是蒺藜科(Zygophyllaceae)的多分枝生態抗逆兼果肉經濟型灌木,主要分布于我國新疆、青海、寧夏、甘肅以及內蒙古等地區的沙漠及鹽堿地上,表現出較強的耐鹽耐旱能力(楊秀艷等,2017)。目前,關于白刺耐鹽機制及其評價研究大多集中于其生長特征、滲透調節系統、活性氧代謝以及光合特性等生理生化方面(李煥勇等,2019;楊秀艷等,2017;左鳳月,2013;王文等,2012;武香等,2012;閆永慶等,2010)。基于蛋白組學方法可研究鹽脅迫下白刺蛋白表達的變化(Chengetal.,2015),然而基因和蛋白質在mRNA水平上增加或減少并不直接影響相應蛋白表達的變化(Gygietal.,1999);即使蛋白質已經被翻譯,也不可能總是能提高酶活性(Sumneretal.,2003)。代謝組學方法克服了蛋白組學方法無法完全描述生化合成的細胞內代謝系統的不足,可通過代謝物的上下調變化以及代謝物實際參與的代謝通路(途徑)來真實地表征植物耐鹽反應和發生過程(蔣丹,2019;Saito et al.,2010)。因此,基于代謝組學方法研究鹽脅迫下白刺的耐鹽反應,可以全面揭示白刺細胞內代謝狀態,獲得有關白刺代謝物的綜合信息,對于深入了解白刺對鹽脅迫響應的代謝途徑及過程非常重要(Nietal.,2015;馮帆等,2019)。
本文以唐古特白刺(N.tangutorum)和西伯利亞白刺(N.sibirica)葉片為研究對象,采用GC-TOF-MS代謝組學方法,研究鹽脅迫下2種白刺葉片代謝物及其代謝通路的變化,以揭示這2種白刺對鹽脅迫的代謝響應機制,為有效提高白刺對鹽堿地的持續生物改良及適應性研究提供基礎理論依據。
1 材料與方法
1.1 種子采集及實生苗培養
西伯利亞白刺種子和唐古特白刺種子均為取自內蒙古巴彥淖爾市磴口縣(海拔1 036 m, 40°32′N, 106°28′E)白刺產地的野生種。分別挑選大小一致、顆粒飽滿的2種白刺的種子先后于50 ℃恒溫水浴鍋浸泡24 h,0.5‰KMnO4溶液避光浸泡2 h,用蒸餾水清洗干凈備用。
采用純沙進行幼苗培養。將0.5% KMnO4溶液殺菌后的純沙置于16 cm×14 cm營養缽中,每缽播白刺種子50粒,每品種12缽,共24缽,種植深度約4 cm,置于溫室內進行培養。培養條件設為平均溫度26 ℃,光照時長14 h,光照強度1 200 Lux,每3天每缽澆灌蒸餾水100 mL,保持純沙含水量60%。
1.2 鹽脅迫處理及取樣
待幼苗生長至25 cm左右且葉量充足時進行鹽脅迫處理。此時唐古特白刺每缽平均出苗35株,西伯利亞白刺每缽平均出苗28株。分別將每種白刺的12缽幼苗隨機分為2組,每組6缽,并統一留苗20株,試驗組每3天澆灌300 mmol·L-1NaCl溶液100 mL,對照組澆灌等量的蒸餾水,為防止鹽分流失,營養缽底部墊有塑料托盤,澆灌后滲出的溶液要重新倒回營養缽內,并定時記錄2種白刺的生長狀況。
脅迫后第7天試驗組有植株頂端嫩葉邊緣出現微黃,此時隨機剪取各缽幼苗中上部第2節完好葉片100 mg,每組6次。所取樣品用錫箔紙包好并編號,液氮浸泡7 min后迅速取出,置于-80 ℃冰箱備用。
1.3 代謝物提取及檢測
取樣本25 mg于2 mL EP管中,加入提取液300 μL、核糖醇5 μL,渦旋30 s后加入瓷珠,45 Hz研磨儀處理4 min,超聲5 min(冰水浴),4 ℃、12 000 r·min-1離心15 min,之后取200 μL上清液于1.5 mL EP管中,每個樣本各取50 μL混合成QC樣本,真空濃縮器中干燥后加入甲氧胺鹽試劑30 μL,混勻,80 ℃孵育30 min,再加入40 μL BSTFA,混勻,70 ℃孵育1.5 h,冷卻至室溫,加入5 μL FAMEs,備測。
使用配有Agilent DB-5MS毛細管柱的Agilent 7890氣相色譜-飛行時間質譜聯用儀(30 m×250 μm×0.25 μm, J&W Scientific, Folsom, CA, USA)進行GC-TOF/MS檢測。
1.4 數據處理
采用ChromaTOF軟件(V 4.3x,LECO)對質譜數據進行峰提取、基線矯正、解卷積、峰積分、峰對齊等分析(馮帆等,2019);使用LECO-Fiehn Rtx5數據庫分析質譜匹配及保留時間指數匹配進行物質定性,利用內標對代謝物數據進行歸一化處理。通過SIMCA軟件(V14.1, Sartorius Stedim Data Analytics AB, Umea, Sweden)對歸一化數據進行對數轉換、中心化格式化處理、主成分分析(PCA)和正交偏最小二乘法(OPLS-DA)分析,依據OPLS-DA模型第一主成分的VIP值>1.0、t檢驗的P<0.05來篩選差異代謝物(李煥勇,2018)。對差異代謝物進行層次聚類分析,并利用KEGG、PubChem等相關數據庫挖掘差異代謝物參與的通路,最后通過富集分析和拓撲分析篩選與代謝物差異相關性最高的關鍵通路。
2 結果與分析
2.1 代謝物的PCA和OPLS-DA分析
通過對2個種24個白刺樣品的總離子流色譜圖(TIC)進行掃描和重疊(圖1),發現TIC出峰保留時間和峰面積都重疊較好。
利用SIMCA軟件對圖1中的代謝譜數據進行PCA和OPLS-DA分析,發現鹽脅迫前后,唐古特白刺代謝物在第二主成分(PC2)上區分顯著,而西伯利亞白刺代謝物在第一主成分(PC1)上區分顯著(圖2),此外,2種白刺在2種處理下的數據點在第一主成分的預測主成分得分上區分顯著(圖3),即2種白刺在鹽脅迫前后均有差異代謝物存在,且西伯利亞白刺代謝物差異更為顯著。由2種白刺的OPLS-DA置換檢驗圖(圖3)可知原模型具有良好的穩健性。

圖1 所有QC樣本的TIC重疊
2.2 差異代謝物的篩選和分析
依據OPLS-DA模型第一主成分的VIP值大于1.0、t檢驗的P值小于0.05篩選組間的差異代謝物(表1),并對這些差異代謝物進行整理分類(圖4)。300 mmol·L-1NaCl脅迫處理前后唐古特白刺共篩選出差異代謝物11種,其中3種顯著下調,8種顯著上調,包括1種脂肪酸、5種有機酸、1種醇以及1種堿基衍生物(上調濃度介于1.32~2.68倍之間);而西伯利亞白刺篩選出差異代謝物108種,其中2種顯著下調,106種顯著上調,包括51種氨基酸、22種糖、11種脂肪酸、8種有機酸、7種醇、5種生物堿以及2種維生素(上調濃度介于0.54~14.45倍之間)。以上結果表明,相比唐古特白刺,西伯利亞白刺有更多的代謝物參與鹽脅迫響應。
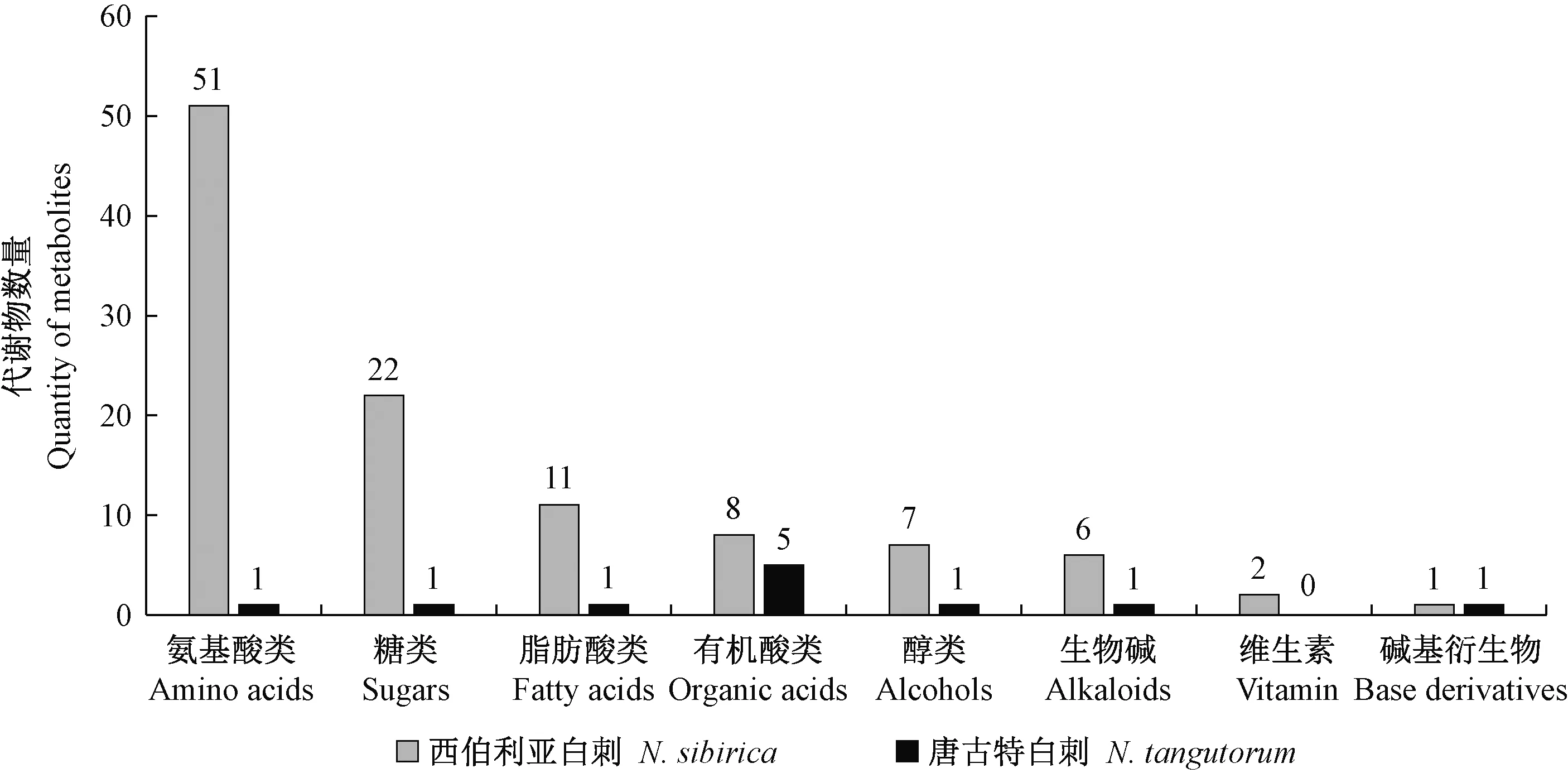
圖4 鹽脅迫后2種白刺葉片中差異代謝物的分類
2.3 差異代謝物的KEGG注釋及代謝通路分析
通過KEGG注釋將唐古特白刺的差異代謝物富集到了6條代謝通路中,而將西比利亞白刺的差異代謝物富集到了46條代謝通路中(表2)。對注釋得到的代謝通路進行富集分析和拓撲分析,進一步篩選到與唐古特白刺代謝物差異相關性最高的關鍵路徑為硫代謝、二羧酸循環及TCA循環(圖5),說明唐古特白刺主要通過加強這3條代謝通路來響應鹽脅迫。其中硫代謝最為顯著,參與此代謝通路的差異代謝物為O-琥珀酰-L-高絲氨酸,其含量顯著上調,為未脅迫時的2.68倍。篩選到與西伯利亞白刺代謝物差異相關性最高的關鍵路徑為纈氨酸、亮氨酸和異亮氨酸的生物合成、C5支鏈二元酸代謝、泛酸和CoA生物合成、苯丙氨酸代謝、硫代謝、絡氨酸代謝,屬于氨基酸生物合成的代謝通路數量多達13條,即氨基酸代謝在西伯利亞白刺響應鹽脅迫過程中發揮主要作用(圖5)。其中,纈氨酸、亮氨酸和異亮氨酸的生物合成途徑差異性最為顯著,參與此代謝通路的差異代謝物分別為檸檬酸、L-亮氨酸、3-蘋果酸異丙酯、L-纈氨酸、L-異亮氨酸以及丙酮酸,其含量均顯著上調,為未脅迫時的1.21~3.12倍。

表2 鹽脅迫后2種白刺差異代謝物映射的KEGG通路

圖5 唐古特白刺(上)和西伯利亞白刺(下)的代謝通路分析
3 討論
植物對逆境脅迫的響應是一個復雜的過程,其中最重要的是代謝的改變,即植物通過調節代謝網絡以誘導產生一系列特殊代謝物,從而達到對生物、非生物脅迫的防御作用(郭鳳丹等,2017)。
3.1 鹽脅迫下2種白刺代謝物的變化
逆境脅迫時,植物通過啟動一系列細胞、分子以及生理生化水平上的調節,以維持細胞代謝平衡(倪建偉等,2014)。代謝物作為細胞調控過程的終產物,是聯系基因型和表型的紐帶,其種類和數量的變化被視為植物對基因或環境變化的最終響應(Zhangetal., 2019;Fernieetal., 2008)。本研究中,鹽脅迫下西伯利亞白刺鑒定出差異代謝物108種,其中106種顯著上調,而唐古特白刺僅鑒定出11種,其中8種顯著上調,即相比唐古特白刺,西伯利亞白刺存在更多的代謝物質參與鹽脅迫調控。就鹽脅迫后2種白刺的代謝差異物相比,代謝物種類差距較大的主要集中在氨基酸、糖以及脂肪酸,而生物堿、堿基衍生物以及維生素等代謝物的種類均比較少,且含量上調、下調不顯著。
鹽脅迫后西伯利亞白刺葉片中51種氨基酸含量顯著上調,而唐古特白刺葉片中1種上調。氨基酸作為蛋白質的組成單位,是能量代謝物質,也是許多生物體內重要含氮化合物的前體(徐春英,2008),其在植物體內的積累可作為一種耐鹽指示物進而反映植物的耐鹽性(燕輝等,2012)。同時,在植物響應非生物脅迫中,氨基酸通過參與植物生理代謝,調節相關基因表達和關鍵酶的活性以提高植物對逆境的抗性(邢芳芳等,2018;楊冬爽,2017)。因此,與唐古特白刺相比,西伯利亞白刺在響應鹽脅迫時氨基酸類代謝物的變化更為顯著,可為其抵御鹽脅迫提供更多的物質和能量(段二龍等,2019)。
可溶性糖參與植物對逆境脅迫的響應(武香等,2012)。與鹽敏感玉米雜交種(Logo)相比,鹽脅迫下耐鹽玉米(Zeamays)雜交種(SR08)通過代謝物葡萄糖、果糖和蔗糖的積累以提高其耐鹽適應性(Richteretal.,2015)。本研究中,鹽脅迫后西伯利亞白刺葉片中22種糖類代謝物顯著上調,這些糖類代謝物在應答鹽脅迫時既可以作為滲透調節物質減緩滲透脅迫,也可作為合成有機溶質的碳架和能量來源,為白刺抵抗鹽脅迫提供重要能量(馮帆等,2019),同時,糖降解過程中的許多代謝產物也是合成氨基酸的前體物質,如絲氨酸、丙氨酸、纈氨酸等。但是,鹽脅迫后唐古特白刺葉片中唯一的糖類代謝物乳糖顯著下調,其他可溶性糖的含量無明顯變化,這表明,可溶性糖可能不參與唐古特白刺對鹽脅迫的應答。
脂肪酸是生物體貯藏能量的主要來源以及細胞膜脂的重要組成成分(馮帆等,2019),參與植物對多種生物、非生物脅迫的響應,通過調控植物體基礎免疫、效應子誘導抗性及系統抗性等多種防御途徑以提高植物對逆境脅迫的抗性(劉文獻等,2014)。生物膜是逆境脅迫中受損傷的主要位點,植物體通過脂肪酸脫飽和維持膜的流動性以適應逆境(楊冬爽,2017;年洪娟等,2012)。Yang等(2017)研究發現,響應鹽脅迫時,耐鹽野生大豆(Glycinesoja)的脂肪酸代謝物含量明顯高于普通野生大豆。本研究中,鹽脅迫后西伯利亞白刺葉片中顯著上調的脂肪酸代謝物共11種,其中,亞麻酸還是植物合成抗逆信號分茉莉酸的前體物質(Xue,2006);而唐古特白刺葉片中脂肪酸代謝物僅亞油酸,說明與唐古特白刺相比較,脂肪酸在西伯利亞白刺的耐鹽調控中發揮著更重要的作用。
3.2 鹽脅迫下2種白刺代謝通路的變化
本研究中,鹽脅迫后西伯利亞白刺的差異代謝物主要富集到氨基酸代謝(相關通路多達13條),且代謝途徑的差異性以纈氨酸、亮氨酸和異亮氨酸的生物合成最顯著,表明西伯利亞白刺可能通過氨基酸代謝迅速合成親和性滲透物質以調節細胞的滲透勢,維持細胞正常的生長和發育(徐春英,2008)。此外,氨基酸在細胞內可經生物合成形成蛋白質,也可進行分解代謝產生ATP,或轉化為糖和脂肪,為抵御鹽脅迫提供能量。鹽脅迫后唐古特白刺并未發現有差異代謝物被富集到氨基酸代謝中,即唐古特白刺在響應鹽脅迫時氨基酸代謝并不顯著。
植物可通過硫代謝的加強以提高其對逆境脅迫的適應性(包雨卓,2017)。在活性氧清除系統中作用最突出的含硫化合物是谷胱甘肽(GSH),GSH是硫代謝的重要產物,也是硫代謝的一種重要的信號物質,GSH的增多有助于維持植物正常的碳和氮代謝(高吉霞,2010)。在唐古特白刺中,與鹽脅迫代謝物差異相關性最高的關鍵路徑為硫代謝,且以參與該途徑的O-琥珀酰-L-高絲氨酸含量上調最為顯著;同時,西伯利亞白刺硫代謝途徑差異性也較顯著,參與該途徑的硫酸鹽和L-半胱氨酸含量也顯著上調,說明硫代謝可能是2種白刺用于抵抗鹽脅迫的有效途徑之一。
TCA循環是糖、脂肪和氨基酸三大營養物質的最終代謝通路和代謝聯系樞紐,該循環代謝的中間產物又可作為許多生物合成途徑的起點(李明達等,2010),在植物對逆境脅迫的響應中扮演著重要角色。本研究中,鹽脅迫后唐古特白刺顯著上調的代謝物檸檬酸參與TCA循環,脅迫后西伯利亞白刺葉片中參與TCA循環的檸檬酸和丙酮酸也顯著上調。檸檬酸可以直接參與機體能量、結構和酶促反應,在TCA循環中可被完全氧化生成ATP,具有高能量利用率(崔媛等,2019)。丙酮酸是糖酵解途徑的最終產物,可以在細胞漿中還原成乳酸供能,還可通過乙酰CoA和TCA循環完成葡萄糖的有氧氧化供能過程,實現植物體內糖、脂肪和氨基酸間的相互轉化(路文靜,2011)。檸檬酸和丙酮酸的顯著上調表明鹽脅迫后2種白刺的TCA循環有所增強,即TCA循環在2種白刺抵御鹽脅迫調控中發揮了重要的作用。
植物響應鹽脅迫時,新陳代謝較為復雜,涉及氨基酸類、糖類及有機酸類等代謝物及相關代謝通路的變化(Chandnaetal.,2013),代謝物的含量反映了與之相關基因的表達量,也表明鹽脅迫對這些代謝途徑相關基因的表達具有調控作用(趙佳利等,2019)。Yang等(2017)研究表明,耐鹽野生大豆主要通過氨基酸代謝與有機酸代謝來響應鹽脅迫,而普通野生大豆主要通過TCA循環來響應鹽脅迫。本研究中2種白刺在鹽脅迫下代謝物與代謝通路均發生變化,且表現出不同的代謝響應策略,與唐古特白刺相比,西伯利亞白刺在響應鹽脅迫時參與的代謝物種類更多,途經更廣,代謝響應更顯著。本研究前期試驗(馮鑫煒等,2015)表明,300 mmol·L-1NaCl脅迫時唐古特白刺平均生物量達2.97 g,根冠比為0.21,葉片電解質滲透率25.27%,MDA含量為8.63 μmol·L-1FW,SOD活性為357.80 U·g-1;西伯利亞白刺平均生物量達5.39 g,根冠比為0.33,葉片電解質滲透率20.08%,MDA含量為6.58 μmol·L-1FW,SOD活性為408.47 U·g-1。西伯利亞白刺從生長和生理方面表現出了優于唐古特白刺的耐鹽性,這與左鳳月(2013)、武香(2012)等的研究結果相吻合。本研究中2種白刺對鹽脅迫的代謝響應與生長、生理響應的一致性進一步驗證了西伯利亞白刺具有比唐古特白刺較好的耐鹽性。
4 結論
唐古特白刺與西伯利亞白刺對鹽脅迫有著不同的代謝響應機制。唐古特白刺通過以有機酸為主的11種代謝物參與鹽脅迫響應,并通過加強以硫代謝為主的6條代謝通路來應答鹽脅迫。西伯利亞白刺通過以氨基酸、糖、脂肪酸為主的108種代謝物參與鹽脅迫響應,并通過加強以氨基酸代謝為主的46條代謝通路來應答鹽脅迫。西伯利亞白刺參與鹽脅迫調控的代謝物較唐古特白刺種類更多,代謝途徑更廣,對鹽脅迫的代謝響應更顯著。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
