常壓室溫等離子體誘變與微生物液滴培養(yǎng)系統(tǒng)聯(lián)用篩選L-組氨酸產(chǎn)生菌
2021-03-11 07:19:46鄭穗平
中國釀造 2021年2期
關(guān)鍵詞:產(chǎn)量
鄧 磊,張 豪,鄭穗平
(華南理工大學(xué) 生物科學(xué)與工程學(xué)院,廣東 廣州 510006)
L-組氨酸(L-histidine)是含有咪唑基團的半必需堿性氨基酸,因其具有免疫調(diào)節(jié)[1]和抗氧化[2]等多種生理功效而被廣泛應(yīng)用于食品[3]、飼料[4]和醫(yī)藥[5]等領(lǐng)域。以血粉等天然蛋白質(zhì)為原料的水解提取法是目前L-組氨酸的主要生產(chǎn)方式,但由于單一分離組氨酸成本高、提取損失率高等因素導(dǎo)致該方法在組氨酸大規(guī)模工業(yè)生產(chǎn)中的應(yīng)用受到限制[6]。發(fā)酵法是最理想的氨基酸生產(chǎn)方法,通過發(fā)酵法生產(chǎn)的L-谷氨酸[7]、L-賴氨酸[8]等大宗氨基酸品種早已形成產(chǎn)業(yè)化。然而L-組氨酸是為數(shù)不多的幾種未能實現(xiàn)大規(guī)模發(fā)酵生產(chǎn)的氨基酸品種之一,因此選育L-組氨酸產(chǎn)生菌株對加快發(fā)酵法生產(chǎn)L-組氨酸具有非常重要的意義。然而由于從頭合成L-組氨酸的途徑長、代謝調(diào)控復(fù)雜,完全利用系統(tǒng)代謝工程改造菌株難度大并且L-組氨酸產(chǎn)量提高不明顯[9]。通過選育氨基酸結(jié)構(gòu)類似物抗性突變株獲得氨基酸產(chǎn)生菌株,依然是目前氨基酸發(fā)酵常規(guī)育種的有效方法[10]。
常壓室溫等離子體(atmospheric and room temperature plasma,ARTP)是一種新型的誘變技術(shù),因其具有突變率高、環(huán)境友好、操作安全等優(yōu)勢[11],已被廣泛應(yīng)用于氨基酸[12]、維生素[13]、二十二碳六烯酸(docosahexaenoic acid,DHA)[14]等高產(chǎn)菌株的選育。雖然ARTP誘變在突變庫容量、突變率等方面發(fā)揮的巨大優(yōu)勢,但突變后的菌株仍然面臨篩選工作量大的問題。近期基于微流控技術(shù)開發(fā)出的全自動高通量微生物液滴培養(yǎng)系統(tǒng)(microbialmicrodropletculturesystem,MMC)[15]具有高通量、自動培養(yǎng)、自動傳代、化學(xué)因子梯度添加、在線檢測液滴的生長以及微生物分選等功能,目前已被用于菌種壓力富集篩選[16]、菌種適應(yīng)性進(jìn)化[17]等方面的研究。
為了獲得L-組氨酸過量產(chǎn)生菌株,以野生型谷氨酸棒桿菌ATCC 14067為出發(fā)菌株,首先利用常壓室溫等離子體(ARTP)對菌株進(jìn)行誘變處理,然后通過全自動高通量微生物液滴培養(yǎng)系統(tǒng)(MMC)和平板相結(jié)合選育耐受高濃度組氨酸結(jié)構(gòu)類似物的抗性突變株。通過孔板發(fā)酵初篩以及搖瓶發(fā)酵復(fù)篩,分析各個結(jié)構(gòu)類似物抗性突變體產(chǎn)L-組氨酸的能力并確定最佳L-組氨酸產(chǎn)生菌株。菌株選育流程如下:
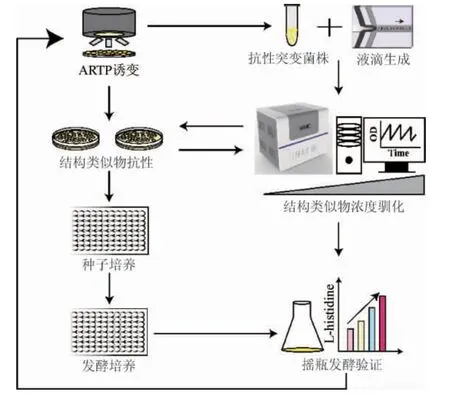
1 材料與方法
1.1 材料與試劑
1.1.1 菌株與主要試劑
野生型谷氨酸棒桿菌(Corynebacterium glutamicum)ATCC 14067:美國菌株保藏協(xié)會購買并保存于實驗室。
L-組氨酸(L-histidine,L-His)、D-組氨酸(D-His)、對氨基苯磺酸(純度99.5%)、玉米漿:上海阿拉丁生化科技股份有限公司;3-氨基-1,2,4-三氮唑(3-amino-1,2,4-triazole,AMT)(純度96%)、3-(N-嗎啉基)丙磺酸(3-morpholinopropanesulfonic acid,MOPS)(純度99.5%):上海麥克林生化科技股份有限公司;腦心浸液(brain heart infusion,BHI):美國BD公司;其他試劑均為國產(chǎn)分析純。
1.1.2 培養(yǎng)基
腦心浸液培養(yǎng)基:腦心浸液37 g/L,固體培養(yǎng)基中加入瓊脂20 g/L,調(diào)pH 7.0,121 ℃滅菌20 min。
選擇培養(yǎng)基:BHI固體培養(yǎng)基滅菌冷卻至60 ℃左右后,添加過濾除菌的不同質(zhì)量濃度的L-組氨酸類似物3-氨基-1,2,4-三氮唑(AMT)和(或)D-組氨酸(D-His),制成組氨酸類似物抗性平板。
種子培養(yǎng)基:(NH4)2SO45 g/L,玉米漿35 g/L,K2HPO41 g/L,MgSO4·7H2O 0.5 g/L,調(diào)pH 至7.0,121 ℃滅菌20 min;葡萄糖25 g/L單獨滅菌。
發(fā)酵培養(yǎng)基:(NH4)2SO420 g/L,KH2PO41 g/L,K2HPO41 g/L,MgSO4·7H2O 0.25 g/L,3-(N-嗎啉基)丙磺酸(MOPS)42 g/L,調(diào)pH 7.0,121 ℃滅菌20 min;葡萄糖40 g/L,尿素5 g/L分別單獨滅菌;FeSO4·7H2O 10 mg/L,MnSO4·H2O 10 mg/L,CaCl210 mg/L,ZnSO4·7H2O 1 mg/L,NiCl2·6H2O 0.02 mg/L,CuSO40.2 mg/L,生物素0.2 mg/L,原兒茶酸0.03 mg/L,硫胺素HCl 0.1 mg/L(上述微量元素、生物素、硫胺素HCl、原兒茶酸配制母液并過濾除菌后按上述質(zhì)量濃度加入)。
1.2 儀器與設(shè)備
UV-2802S型分光光度計:上海尤尼柯儀器有限公司;ARTP-M誘變系統(tǒng)、MMC-B微生物液滴細(xì)胞培養(yǎng)儀:天木生物科技有限公司;5810R臺式離心機:德國Eppendorf公司;恒溫培養(yǎng)搖床:上海知楚儀器有限公司;DHP-9052A電熱恒溫培養(yǎng)箱:紹興市景邁儀器設(shè)備有限公司;L550湘儀離心機:濟南千司生物技術(shù)有限公司;48深孔板:上海甘薇生物科技有限公司;48淺孔板:美國康寧公司。
1.3 方法
1.3.1 ARTP誘變
將活化的菌液轉(zhuǎn)接于種子培養(yǎng)基,控制起始OD600nm值為0.2~0.3,待培養(yǎng)OD600nm值為0.8~1.0時離心收集菌體。用無菌生理鹽水洗滌3次,再將菌體稀釋至OD600nm值為0.6~0.8。取10 μL菌液均勻涂抹在載片上,隨后載片放入ARTP儀器樣品板中,與等離子炬噴嘴對齊并暴露于輻射下。ARTP誘變參數(shù):載片與氣流端口距離2 mm;射頻功率100 W;氣流量10 SLM;溫度25~35 ℃。實驗中各個組ARTP處理為0、30~270 s,處理過后的載片用1 mL 的0.85%無菌生理鹽水洗下菌體并稀釋適當(dāng)倍數(shù)后,取100 μL均勻涂抹于BHI培養(yǎng)基固體平板上,每組設(shè)置3個平行,30 ℃培養(yǎng)3~4 d后進(jìn)行單菌落計數(shù),并計算致死率,其計算公式如下:

式中:A為未經(jīng)ARTP誘變處理對照菌的總菌落數(shù),CFU/mL;B為經(jīng)ARTP誘變處理后對應(yīng)的總菌落數(shù),CFU/mL。
1.3.2 誘變菌株篩選
初篩:根據(jù)菌落生長的大小、圓整和豐度,從組氨酸結(jié)構(gòu)類似物抗性平板上隨機挑選單菌落進(jìn)行孔板篩選。種子培養(yǎng)使用48孔淺孔板,裝液量400 μL,于30 ℃、800 r/min條件下?lián)u床培養(yǎng)24 h;發(fā)酵培養(yǎng)使用48孔深孔板,裝液量800 μL,接種量為10%,每株菌3個平行,于30 ℃、650 r/min條件下?lián)u床培養(yǎng)72 h后,發(fā)酵結(jié)束后用孔板離心機收集上清液進(jìn)行組氨酸含量測定。
復(fù)篩:將孔板篩選得到的組氨酸生產(chǎn)菌株,進(jìn)行搖瓶復(fù)篩驗證。種子培養(yǎng)使用250 mL搖瓶,裝液量25 mL,于30 ℃、250 r/min的搖床培養(yǎng)24 h;發(fā)酵培養(yǎng)使用500 mL搖瓶,裝液量30 mL,接種量為10%,每株菌三個平行,于30 ℃、250 r/min條件下?lián)u床培養(yǎng)72 h后,發(fā)酵結(jié)束后測定發(fā)酵上清液組氨酸含量。
1.3.3 AMT抗性菌株馴化
將從組氨酸結(jié)構(gòu)類似物AMT抗性平板篩選得到的組氨酸生產(chǎn)菌株,接種于含有一定質(zhì)量濃度組氨酸結(jié)構(gòu)類似物AMT的BHI培養(yǎng)基中,30 ℃、250 r/min振蕩培養(yǎng)24 h。培養(yǎng)結(jié)束后,用新滅菌的BHI培養(yǎng)基將菌液稀釋至OD600nm值為0.8~1.2,之后注入液滴微流控培養(yǎng)儀專用進(jìn)樣瓶(液滴生成進(jìn)樣瓶),同時配制一個含低質(zhì)量濃度AMT(2 g/L)的BHI培養(yǎng)基進(jìn)樣瓶和一個含高質(zhì)量濃度AMT(10 g/L)的BHI培養(yǎng)基進(jìn)樣瓶。通過MMC設(shè)備使菌株在含組氨酸類似物的培養(yǎng)基中培養(yǎng)、傳代并伴隨不同質(zhì)量濃度的AMT添加,最后根據(jù)菌株馴化生長情況選擇相應(yīng)液滴,進(jìn)行收集。
1.3.4L-組氨酸含量的測定
采用Pauly比色法測定發(fā)酵液中組氨酸的含量[18]。L-組氨酸標(biāo)準(zhǔn)曲線的繪制:將配制好的質(zhì)量濃度為0.1 g/L的組氨酸標(biāo)準(zhǔn)溶液分別準(zhǔn)確吸取0、0.1 mL、0.2 mL、0.3 mL、0.4 mL、0.5 mL于10 mL比色管中,加入0.2 mL 1%對氨基苯磺溶液和0.2 mL 5%亞硝酸鈉溶液,混勻后25 ℃水浴反應(yīng)15 min;反應(yīng)結(jié)束后迅速加入10%碳酸鈉溶液0.6 mL,振蕩10 s后,加入2 mL體積分?jǐn)?shù)20%的乙醇溶液,混勻后蓋上瓶塞,室溫條件下靜置5 min;之后定容至10 mL。在本實驗條件下利用分光光度計分析反應(yīng)產(chǎn)物最大吸收波長以及對應(yīng)的吸光度值。
1.3.5 細(xì)胞生物量測定
將發(fā)酵液稀釋20倍,在波長600 nm處測量吸光度值。
1.3.6 突變菌株遺傳穩(wěn)定性分析
將篩選出的組氨酸生產(chǎn)菌株,接種于BHI液體培養(yǎng)基進(jìn)行活化,然后將其轉(zhuǎn)移至種子培養(yǎng)基中并連續(xù)轉(zhuǎn)移7次,之后進(jìn)行搖瓶發(fā)酵,測試突變菌株組氨酸生產(chǎn)能力以及生長狀況。
2 結(jié)果與分析
2.1 L-組氨酸標(biāo)準(zhǔn)曲線的確定
通過在300~800 nm波長條件下掃描Pauly試劑以及不同濃度的L-組氨酸Pauly顯色產(chǎn)物,結(jié)果見圖1。由圖1a可知,當(dāng)反應(yīng)中有L-組氨酸參與時,在波長500 nm處存在吸收峰;由圖1b可知,在此波長條件下隨著反應(yīng)的L-組氨酸質(zhì)量濃度增加,對應(yīng)的吸光度值也在增加;由圖1c可知,以L-組氨酸質(zhì)量濃度(x)為橫坐標(biāo),對應(yīng)波長500 nm處吸光度值(y)為縱坐標(biāo),繪制L-組氨酸的標(biāo)準(zhǔn)曲線方程為y=13.678x+0.043,相關(guān)系數(shù)R2=0.996 4,L-組氨酸檢測質(zhì)量濃度在0.01~0.05 g/L范圍內(nèi)線性關(guān)系良好,可用于發(fā)酵液中L-組氨酸含量的測定。

圖1 Pauly試劑(a)和L-組氨酸(b)的顯色產(chǎn)物吸光度值檢測結(jié)果及L-組氨酸標(biāo)準(zhǔn)曲線(c)Fig.1 Result absorbance detection of the color product of Pauly reagent (a) and L-histidine (b) and standard curve of L-histidine (c)
2.2 ARTP誘變致死曲線的測定
按照方法1.3.1,對出發(fā)菌野生型谷氨酸棒桿菌ATCC 14067進(jìn)行ARTP誘變,按照ARTP處理時間為變量,測定菌株致死曲線。如圖2所示,當(dāng)ARTP照射180 s時,菌株致死率可達(dá)83.05%;進(jìn)一步將ARTP處理時間增至210 s時,菌株致死率高達(dá)93.26%;而ATRP處理240 s后菌株致死率基本穩(wěn)定在98.11%。根據(jù)之前的研究報道[19],當(dāng)菌株致死率約為90%時菌株正突變率最高,但由于本實驗ARTP照射時間超過240 s后,幾乎沒有菌體存活下來,因此確定野生型谷氨酸棒桿菌ATCC 14067最佳的誘變時間210 s。

圖2 谷氨酸棒桿菌ATCC 14067 常壓室溫等離子誘變致死曲線Fig.2 Lethal rate curve of Corynebacterium glutamicum ATCC 14067 mutated by ARTP
2.3 L-組氨酸產(chǎn)生菌的選育
2.3.1 AMT抗性突變株的篩選
將野生型谷氨酸棒桿菌ATCC 14067利用ARTP處理后,分別稀釋涂布于含5種不同質(zhì)量濃度的AMT抗性平板上。從抗性平板上一共挑取264個單菌落,通過孔板發(fā)酵72 h后檢測組氨酸產(chǎn)量,結(jié)果如表1所示。由表1可知,5.0 g/L和4.0 g/L AMT 抗性平板上的菌株明顯比1.0 g/L、2.0 g/L、3 g/L AMT抗性平板的菌株的正突變率要高,這說明從抗性較高的突變菌株中,篩選獲得組氨酸積累菌株的可能性更大。

表1 不同質(zhì)量濃度的3-氨基-1,2,4-三氮唑抗性突變株篩選結(jié)果Table 1 Screening results of resistant mutants by different concentrations of 3-amino-1,2,4-triazole
從117株正突變菌株中,挑選出產(chǎn)量最高的6株菌株(編號為Cg-A1~Cg-A6)與出發(fā)菌株谷氨酸棒桿菌ATCC 14067(編號為Cg-A0)一起進(jìn)行搖瓶發(fā)酵進(jìn)一步驗證組氨酸產(chǎn)量以及檢測對應(yīng)的細(xì)胞生物量,結(jié)果如圖3所示。由圖3可知,菌株Cg-A1組氨酸產(chǎn)量最高可達(dá)(110.39±2.09)mg/L,比出發(fā)菌株提高約493%,但菌株Cg-A1生長OD600nm值較出發(fā)菌株卻下降了38.11%,下降幅度較為明顯。

圖3 菌株Cg-A0和Cg-A1~A6搖瓶發(fā)酵的OD600nm值和L-組氨酸產(chǎn)量Fig.3 OD600nmvalue and L-histidine yields of Cg-A0 and Cg-A1-A6 after shake-flask fermentation
2.3.2 AMT抗性菌株穩(wěn)定表型的篩選
以菌株Cg-A1作為出發(fā)菌株,按照方法1.3.3對其進(jìn)行AMT濃度耐受性馴化,結(jié)果如圖4所示。在傳代時間恒定條件下,由圖4a可知,隨著培養(yǎng)基中AMT的濃度在2~10 g/L范圍內(nèi)增加,菌株整體生長呈現(xiàn)下降趨勢,并且當(dāng)AMT質(zhì)量濃度為10g/L時,OD600nm值下降至5左右。因此需要在AMT10g/L條件下連續(xù)馴化多代,結(jié)果見圖4b。由圖4b可知,液滴在不含AMT的條件下傳代培養(yǎng)3代,OD600nm值能夠恢復(fù)并且穩(wěn)定在12~14。然后將AMT質(zhì)量濃度固定為10 g/L,在前兩代,液滴生長OD600nm值整體有一個明顯的下降,但之后液滴整體生長呈現(xiàn)穩(wěn)定的趨勢,此時表明菌株能夠穩(wěn)定耐受10 g/L AMT。根據(jù)生長狀況提取馴化OD600nm值最高的6個液滴進(jìn)行搖瓶發(fā)酵檢測L-組氨酸產(chǎn)量情況,由圖5可知,馴化之后的菌株(編號為Cg-B1~Cg-B6)較馴化之前的菌株Cg-A1,L-組氨酸產(chǎn)量都有明顯的提高,其中菌株Cg-B1的組氨酸產(chǎn)量最高可達(dá)(0.23±0.013)g/L,同時該菌株的生長OD600nm值與野生型菌株Cg-14067的OD600nm值相接近。

圖4 組氨酸類似物3-氨基-1,2,4-三氮唑不同含量條件下菌體生長曲線Fig.4 Growth curves of the strain under different concentration of histidine analog 3-amino-1,2,4-triazole

圖5 馴化后菌株搖瓶發(fā)酵OD600nm值和L-組氨酸產(chǎn)量Fig.5 OD600nmvalue and L-histidine yields of strain after domestication by shake-flask fermentation
2.3.3 AMT和D-His雙重抗性突變株的篩選
D-組氨酸(D-His)與組氨酸結(jié)構(gòu)相似度極高,已被用作組氨酸結(jié)構(gòu)類似物抗性突變株的表型篩選[20]。為了篩選到組氨酸產(chǎn)量進(jìn)一步提高的菌株,以菌株Cg-B1作為出發(fā)菌,采用ARTP迭代誘變的處理方式(即:將每一輪篩選得到組氨酸產(chǎn)量最高的菌株作為下一輪誘變的起始菌株),并且于兩種組氨酸類似物AMT和D-His在平板上AMT質(zhì)量濃度固定為10 g/L、D-His的質(zhì)量濃度依次從2 g/L遞增至8 g/L。從雙重抗性平板上的挑取單菌落,通過48孔板發(fā)酵72 h后檢測發(fā)酵上清液中L-組氨酸含量,各輪檢測結(jié)果如圖6所示。由圖6可知,最終從耐受組氨酸結(jié)構(gòu)類似物AMT 10 g/L和D-His 8 g/L表型中篩選到L-組氨酸產(chǎn)量最高的菌株Cg-F4,該菌株積累L-組氨酸產(chǎn)量可達(dá)(0.561±0.016)g/L。

圖6 組氨酸結(jié)構(gòu)類似物3-氨基-1,2,4-三氮唑和D-His抗性突變株初篩選結(jié)果Fig.6 Screening results of histidine analogs 3-amino-1,2,4-triazole and D-His resistant mutants
2.4 誘變菌株的遺傳穩(wěn)定性
按照方法1.3.6,對突變株Cg-F4 的生長和組氨酸生產(chǎn)的遺傳穩(wěn)定性進(jìn)行考察,結(jié)果如圖7所示。由圖7可知,菌株Cg-F4傳代7次之后,分別發(fā)酵72 h后L-組氨酸可穩(wěn)定在0.55 g/L左右,OD600nm值都在9左右,表明突變菌株Cg-F4菌株具有良好的遺傳穩(wěn)定性。
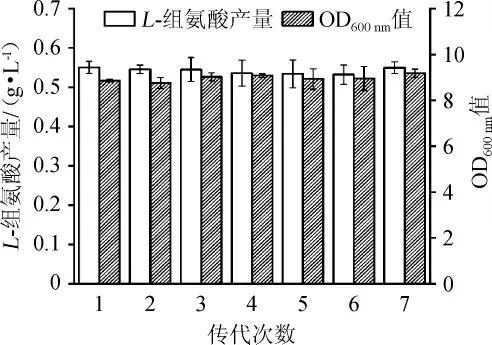
圖7 突變菌株Cg-F4的遺傳穩(wěn)定性分析結(jié)果Fig.7 Analysis results of genetic stability for mutant Cg-F4
2.5 討論
L-組氨酸的生物合成受其自身的反饋抑制和反饋阻遏,消除這種反饋調(diào)節(jié)是開發(fā)組氨酸產(chǎn)生菌株的重要一步[21]。選育組氨酸結(jié)構(gòu)類似物抗性突變株解除L-組氨酸自身的反饋調(diào)節(jié),使菌株積累L-組氨酸[22]。本研究通過ARTP誘變技術(shù)與MMC技術(shù)相結(jié)合可以高效選育穩(wěn)定表型的組氨酸結(jié)構(gòu)類似物抗性突變株,同時利用L-組氨酸與Pauly試劑反應(yīng)生成顏色化合物這種特性,實現(xiàn)了L-組氨酸的快速測定,提高了L-組氨酸產(chǎn)生菌的篩選效率。目前使用了兩種組氨酸結(jié)構(gòu)類似物作為篩選壓力,篩選到L-組氨酸產(chǎn)生菌Cg-F4,為了進(jìn)一步提高菌株產(chǎn)L-組氨酸的能力,可以在下兩個方面進(jìn)行研究:進(jìn)一步提高目前組氨酸結(jié)構(gòu)類似物的濃度或者引入其他種類組氨酸結(jié)構(gòu)類似物抗性標(biāo)記,更進(jìn)一步解除了組氨酸自身的反饋調(diào)節(jié),使產(chǎn)量不斷提升。發(fā)酵條件及工藝控制的優(yōu)化對進(jìn)一步提高L-組氨酸的積累至關(guān)重要,因此,在選育高產(chǎn)菌種后,還應(yīng)進(jìn)行發(fā)酵條件及工藝控制的優(yōu)化,包括發(fā)酵培養(yǎng)基的組成、pH值、溫度、溶氧量等。
3 結(jié)論
本研究首次將ARTP誘變技術(shù)與MMC技術(shù)相結(jié)合定向選育組氨酸結(jié)構(gòu)類似物抗性突變株。通過孔板發(fā)酵初篩以及搖瓶發(fā)酵復(fù)篩,最終從耐受兩種組氨酸結(jié)構(gòu)類似物抗性突變株中篩選獲得菌株Cg-F4 其L-組氨酸產(chǎn)量可達(dá)(0.561±0.016)g/L。菌株Cg-F4經(jīng)過7次連續(xù)傳代實驗,證明該突變菌株具有良好的遺傳穩(wěn)定性。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當(dāng)代水產(chǎn)(2021年10期)2021-12-05 16:31:48
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
今日農(nóng)業(yè)(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中國果業(yè)信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
