BTV-16四結構蛋白的共表達與病毒樣顆粒的制備
2021-03-10 02:41:30黃超華花群義曹琛福史衛軍阮周曦林彥星葉奕優陳金頂
中國獸醫學報 2021年1期
黃超華,花群義,曹琛福,史衛軍,阮周曦,林彥星,葉奕優,陶 虹,王 瀟,陳金頂*
(1.華南農業大學 獸醫學院,廣東 廣州 510642;2.深圳海關動植物檢驗檢疫技術中心,廣東 深圳 518045)
藍舌病(bluetongue,BT)是一種蟲媒性傳染病,屬于世界動物衛生組織(Office International des Epizooties,OIE)規定及時上報的動物疫病。BT的流行和暴發會帶來巨大的經濟損失。目前疫苗免疫是預防BT的主要有效手段[1-2]。藍舌病病毒(bluetongue virus,BTV)病毒樣顆粒(virus-like particles,VLPs)疫苗由于具有DIVA (disabled infectious single animal)性質基因工程苗的特點,因而成為BT疫苗研究的熱點之一[3-4]。該VLPs由BTV的4種結構蛋白 VP2、VP3、VP5和VP7組裝而成,其中60個VP3二聚體組裝形成直徑為55 nm的BTV病毒亞核心,其外層為VP7三聚體構成的二十面體結構,兩者共同構成BTV CLPs[5]。VP2是BTV型特異性抗原,是BTV血清型的主要決定因素,能引起血凝,誘導產生中和抗體[6]。而VP5是BTV唯一的糖基化蛋白,在VP2免疫中和反應中起到了重要的增強作用[7]。VP5與VP2一樣以三聚體形式存在,VP5三聚體最終形成球形基序與VP2三聚體形成的三腳蛋白復合體共同構成BTV VLPs的外衣層結構[3,8-10]。
流行病學調查和有關的病毒分離報道顯示,我國存在的BTV血清型非常多,其中1和16型為主要血清型。目前我國雖然在BT疫苗研究上取得了諸多進展,如滅活疫苗、DNA疫苗、重組痘病毒載體疫苗和重組腺病毒疫苗等均能產生中和抗體,對動物有一定的免疫保護作用[11-12],但在BTV VLPs疫苗方面研究較少。本試驗通過基因工程技術獲取具有4個獨立啟動子的載體,構建具有BTV-16型4種結構蛋白基因VP2、VP3、VP5和VP7的重組桿狀病毒并進行真核表達,以期為BTV VLPs疫苗的研發奠定基礎。
1 材料與方法
1.1 主要材料pFastBac-Dual質粒購自Invitrogen公司產品;感受態細胞DH5α和DH10Bac購自Invitrogen公司;BTV-16型綿羊陽性血清和昆蟲細胞Sf9由本實驗室保存;Cellfectin?Ⅱ Reagent、SF-900Ⅲ培養基、PureLinkTMHiPure質粒純化試劑盒購自Invitrogen公司;限制性內切酶以及Taq酶、BacPAKTMBaculovirus Rapid Titer Kit購自TaKaRa公司;Rabbit Anti-SheepIgG H&L(HRP)購自Abcam公司;TMB顯色試劑盒購自上海生工公司;弗氏完全佐劑、弗氏不完全佐劑購自Sigma公司。
1.2 引物的設計與合成分別根據BTV-16 S2、L3、S6和S7基因參考序列設計并合成各自的特異性引物用于構建重組質粒的篩選和鑒定;根據F4序列設計并合成F4序列特異性引物用于載體的改造;參照Bac-to-Bac昆蟲桿狀表達系統操作說明,合成1對公用引物M13-F和M13-R,用于重組桿粒的篩選和鑒定。具體引物序列見表1。
1.3 pFastBac-Dual載體的改造為共表達BTV結構蛋白VP2、VP5、VP3和VP7,對載體pFastBac-Dual進行改造,使其具有4個多克隆位點(multiple cloning site,MSC),并且每個多克隆位點均具有各自獨立的啟動子。分析載體pFastBac-Dual的全序列,發現在載體的3 510堿基處為Bsp1407Ⅰ酶切位點且是唯一的,3 985堿基處為SnaBⅠ酶切位點且是唯一的,同時在3 510~3 985堿基處沒有任何元件,因此可作為改造區域對載體進行改造。

表1 所用引物序列
由于原載體已有2個多克隆位點,各自的啟動子分別為Polyhedrin和P10,因此本研究通過Bsp1407Ⅰ和SnaBⅠ雙酶切載體pFastBac-Dual,切除3 510~3 985堿基處的序列,隨后插入一段合成的序列F4(該序列含3 510~3 985堿基的序列和2個多克隆位點,且該2個多克隆位點具有各自獨立的啟動子),該序列共1 347 bp,其組成如圖1所示,從而完成載體的改造。改造后的載體命名為pF4,整個載體圖譜見圖2,其中MSC1和MSC2為2個新插入的多克隆位點,其啟動子分別為P10和Polyhedrin,而MSC3和MSC4為原有的2個多克隆位點,其啟動子分別為P10和Polyhedrin。
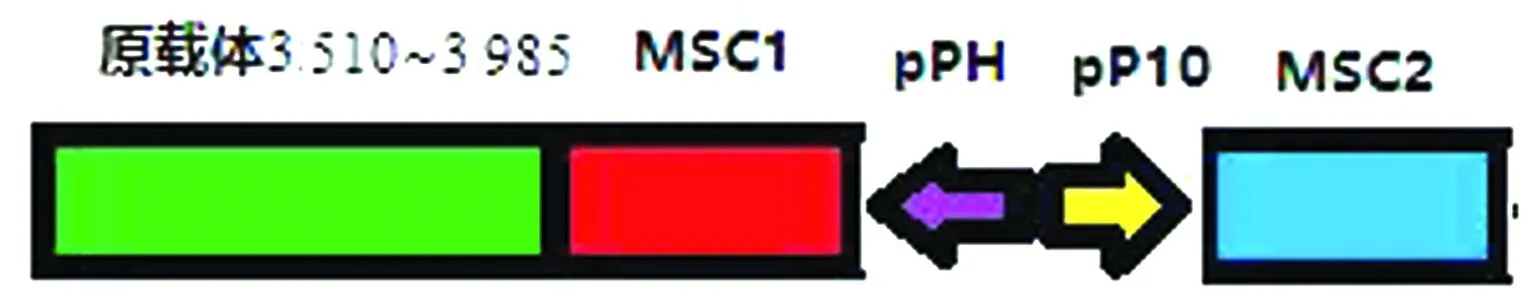
圖1 合成序列F4組成
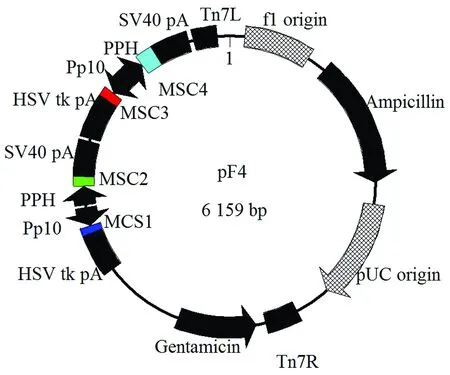
圖2 pF4圖譜
1.4 pF4-VP2-VP5-VP3-VP7的構建通過NCBI 基因庫比對找到與BTV-16的4個結構蛋白VP2、VP5、VP3和VP7同源性較高的基因序列(AB686226.1、KF664138.1、KP339226.1和JX007-928.1),并以此為參考序列,通過上海生工公司合成VP2、VP5、VP3和VP7基因序列從而獲得重組質粒pUC-VP2(VP2序列的5′和3′端分別有酶切位點EcoRⅠ和SalⅠ)、pUC-VP5(VP5序列的5′和3′端分別有酶切位點XhoⅠ和SphⅠ)、pUC-VP3(VP3序列的5′和3′端分別有酶切位點NcoⅠ和 NheⅠ)和pUC-VP7(VP7序列的5′和3′端分別有酶切位點AatIⅠ和Eco81Ⅰ)。按照常規的分子克隆技術,即雙酶切、T4連接、轉化感受態細胞、菌落PCR和雙酶切鑒定等方法,依次將VP2、VP5、VP3和VP7基因插入到改造后的載體pF4的4個多克隆位點,其中VP2插入到MSC4,VP5插入到MSC3,VP3插入到MSC1,VP7插入到MSC2,從而構建重組質粒pF4-VP2-VP5-VP3-VP7,質粒圖譜見圖3。
1.5 重組桿粒Bacmid-VP2-VP5-VP3-VP7的提取通過熱轉化的方式將重組質粒pF4-VP2-VP5-VP3-VP7轉化入感受態細胞DH10Bac中,平鋪于含卡那霉素、四環素、IPTG和Bluo-gal的LB平板上,37 ℃培養48 h;挑取單個白色菌落劃線于新的上述平板并繼續培養48 h。挑取單個白色菌落,進行菌落PCR鑒定,確定VP2-VP5-VP3-VP7是否轉座成功。鑒定結果為陽性的,過夜大量培養,并用PureLinkTMHiPure質粒純化試劑盒大量提取純化重組桿粒Bacmid-VP2-VP5-VP3-VP7。
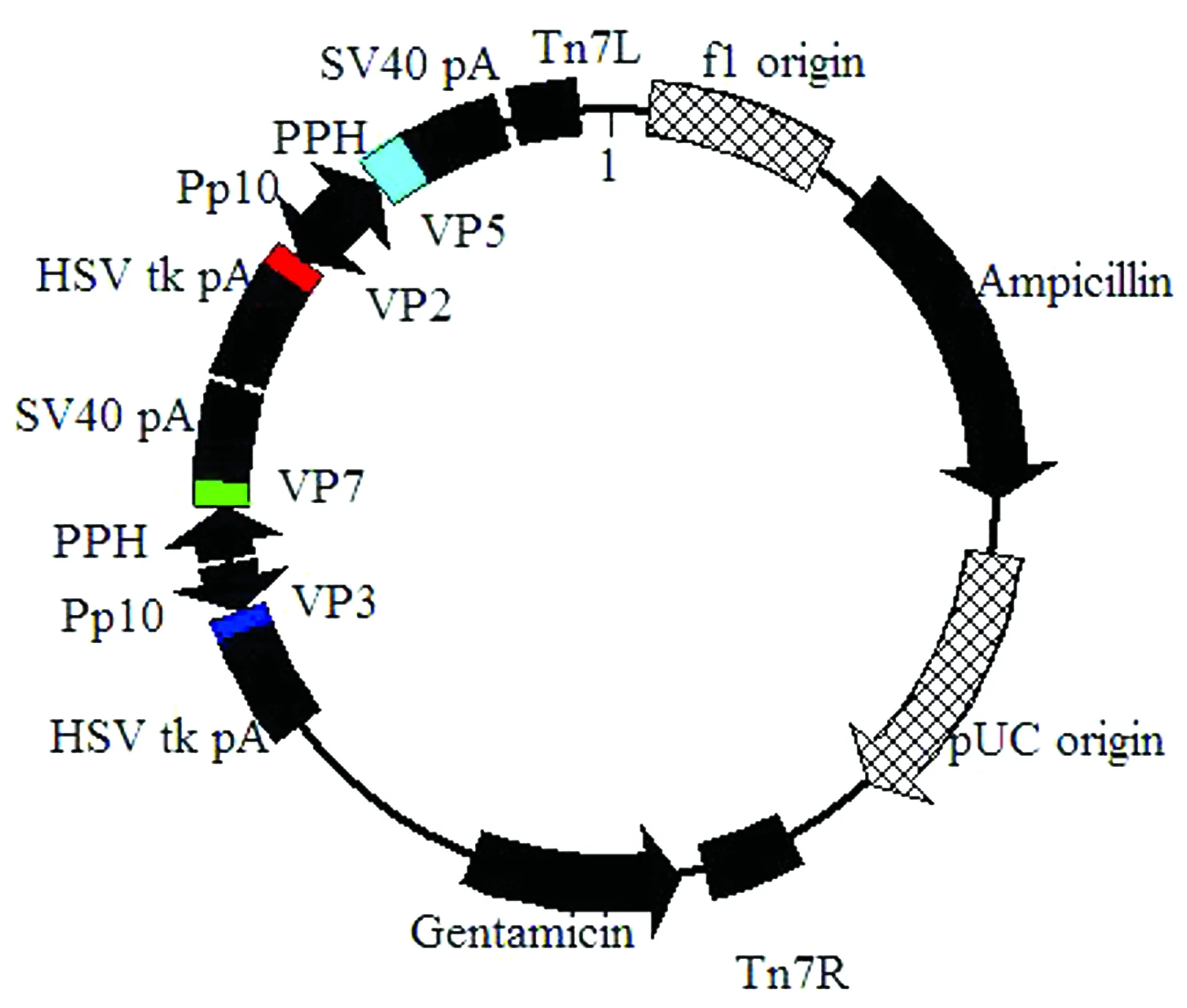
圖3 pF4-VP2-VP5-VP3-VP7圖譜
1.6 構建重組桿狀病毒rBac-VP2-VP5-VP3-VP7用轉染試劑盒Cellfectin?Ⅱ Reagent將重組桿粒Bacmid-VP2-VP5-VP3-VP7轉染Sf9細胞,設空白對照,持續觀察細胞病變情況。在細胞病變明顯時,收集細胞培養上清,即P1代重組桿狀病毒rBac-VP2-VP5-VP3-VP7,并繼續傳代以提高重組桿狀病毒的滴度。使用TaKaRa公司的BacPAKTMBaculovirus Rapid Titer Kit測定重組桿狀病毒的滴度。
1.7 BTV VLPs的純化將無血清培養的Sf9細胞轉瓶至75 cm2細胞培養瓶中(60%~80%),貼壁1 h 后,接入適量的重組桿狀病毒,72 h時棄細胞培養液,加入1 mL細胞裂解液冰上裂解15 min,并反復凍融3次使細胞充分裂解,12 000 r/min,4 ℃離心10 min收集細胞裂解上清。按照前述方法大量接毒、裂解細胞、離心收集細胞裂解上清,并用Millipore超濾管進行離心濃縮。采用蔗糖梯度離心純化濃縮后的細胞裂解上清中的BTV VLPs,蔗糖梯度離心條件參考相關文獻[13]。
1.8 4種結構蛋白的抗原性鑒定取少量純化產物進行SDS-PAEG和Western blot分析,其中Western blot 分析時,以Protein-Free T20 (PBS) Blocking Buffer室溫封閉1 h;洗滌后加入綿羊BTV-16陽性血清(1∶500稀釋),37 ℃孵育1 h;洗滌后加入Rabbit Anti-Sheep IgG H&L(HRP)(1∶5 000稀釋),37℃孵育45 min,洗滌后顯色。
1.9 BTV VLPs電鏡觀察取少量蔗糖梯度離心純化的樣品,采用醋酸鈾負染法對樣品進行負染并固定在載網上,在TEM電鏡下觀察VLPs形態。
2 結果
2.1 pFastBac-Dual載體的改造由上海生工公司合成F4序列,獲得重組質粒pUC-F4。用Bsp1407 Ⅰ和SnaBⅠ雙酶切載體pFastBac-Dual和重組質粒pUC-F4,獲得具有黏性末端的pFastBac-Dual和F4片段。通過連接、轉化、鑒定(菌落PCR、雙酶切和測序),構建了改造后的載體pF4。用F4序列特異性引物進行菌落PCR可得到1 347 bp左右的目的條帶,以Bsp-1407Ⅰ和SnaBⅠ雙酶切提取的質粒可得到4 853 bp 大小的原載體條帶和1 347 bp大小的目的條帶,結果見圖4。并對改造后的載體pF4進行全基因測序,確認載體pF4包含4個克隆位點和4個獨立的啟動子,且全序列未發生突變,所有元件完整。
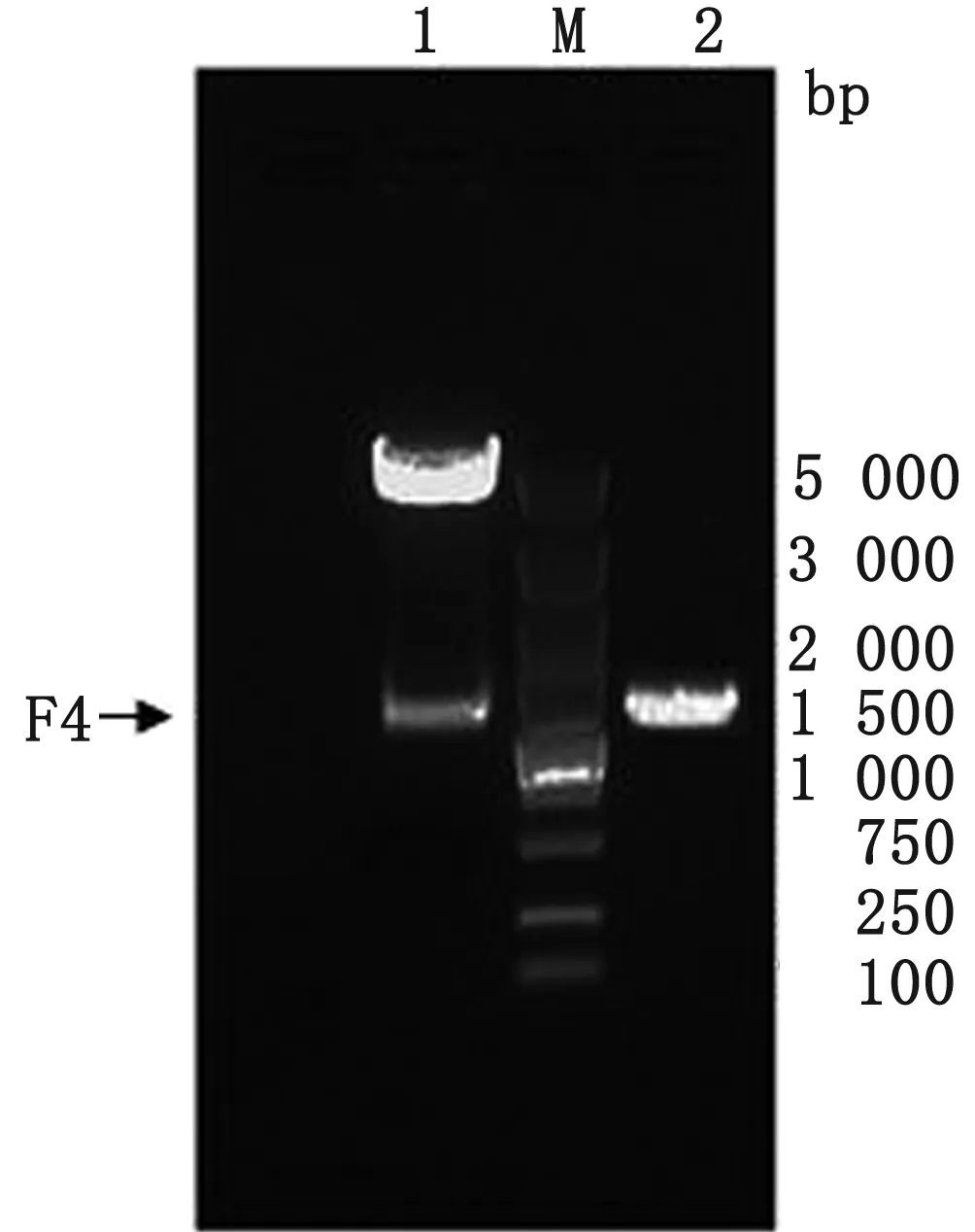
M.DL5000 DNA Marker;1.Bsp1407Ⅰ和SnaBⅠ雙酶切; 2.菌落PCR
2.2 重組質粒pF4-VP2-VP5-VP3-VP7的構建按照常規的分子克隆技術,依次將VP2、VP5、VP3和VP7插入到載體pF4中,從而構建了重組質粒pF4-VP2-VP5-VP3-VP7。構建鑒定結果見圖5。
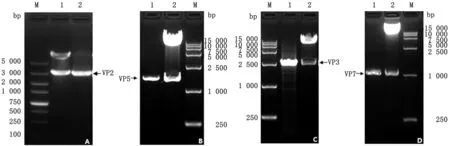
A.重組質粒pF4-VP2的鑒定結果(M.DL5000 DNA Marker;1.雙酶切;2.菌落PCR);B.重組質粒pF4-VP2-VP5的鑒定結果(M.DL15000 DNA Marker;1.菌落PCR;2.雙酶切);C.重組質粒pF4-VP2-VP5-VP3的鑒定結果(M.DL15000 DNA Marker;1.菌落PCR;2.雙酶切);D.重組質粒pF4-VP2-VP5-VP3-VP7的鑒定結果(M.DL15000 DNA Marker;1.菌落PCR;2.雙酶切)
2.3 重組桿粒Bacmid-VP2-VP5-VP3-VP7的提取將重組質粒pF4-VP2-VP5-VP3-VP7轉化入DH10Bac發生轉座,通過藍白斑和菌落PCR篩選出轉座成功的菌落,并用質粒純化試劑盒大量提取純化重組桿粒Bacmid-VP2-VP5-VP3-VP7。分別以VP2特異性上下游引物、VP3特異性上下游引物、VP5特異性上下游引物和VP7特異性上下游引物進菌落PCR,可以獲得2 900,2 700,1 500,1 000 bp的目的條帶分別對應VP2、VP3、VP5和VP7;為進一步確認是否轉座成功,分別以VP2-F+M13-R、M13F+VP2-R引物組合進行PCR鑒定,電泳結果顯示分別可以獲得3 600,12 000 bp的目的條帶,與預期的結果相符,結果見圖6。
2.4 重組桿狀病毒rBac-VP2-VP5-VP3-VP7的獲得用轉染試劑盒將重組桿粒Bacmid-VP2-VP5-VP3-VP7轉染Sf9細胞,轉染后48~72 h可以觀察到明顯的細胞病變:與正常Sf9細胞相比,細胞數量變少,體積變大且發生臌脹、變得容易脫落,并有少量細胞破碎,見圖7。轉染后超過72 h,Sf9細胞大量破碎。
2.5 BTV VLPs的純化及SDS-PAGE和Western blot 分析通過蔗糖梯度離心純化得到BTV VLPs,取少量純化產物進行SDS-PAGE(圖8)和Western blot(圖9)分析,結果均出現4條分別為110,100,60,38 kDa左右的主要條帶,所對應的蛋白分別為VP2、VP3、VP5和VP7,說明本研究構建的重組桿狀病毒可以在Sf9細胞中共表達BTV-16的4種結構蛋白并且具有免疫原性。
2.6 電鏡觀察結果取少量純化產物通過醋酸鈾負染固定于載網上,在透射電鏡下觀察其形態。結果顯示,純化的樣品中存在數量較多的中空顆粒,大小形態略有不同,其中也有部分中空顆粒直徑為75~85 nm,顆粒側壁較厚,且表面的突起不明顯,其形態、大小與文獻報道的BTV VLPs一致。圖10為透射電鏡下不同視野觀察到的具有典型形態的BTV-16的VLPs,并有少量中空顆粒直徑為60~70 nm,顆粒表面有非常明顯的突起,其形態、大小均與文獻報道的桿狀病毒表達系統獲得的BTV CLPs一致。
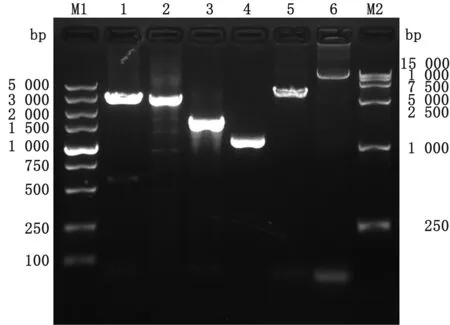
M1.DL5000 DNA Marker;1.引物組合(VP2-F+VP2-R)菌落PCR結果;2.引物組合(VP3-F+VP3-R)菌落PCR結果;3.引物組合(VP5-F+VP5-R)菌落PCR結果;4.引物組合(VP7-F+VP7-R)菌落PCR結果;5.引物組合(VP2-F+M13-R)菌落PCR結果;6.引物組合(M13-F+VP2-R)菌落PCR結果;M2.DL15000 DNA Marker
A.正常Sf9細胞;B.重組桿粒Bacmid-VP2-VP5-VP3-VP7轉染Sf9細胞
3 討論
VLPs的制備均是通過構建的表達載體在原核細胞或真核細胞中共表達多個病毒結構蛋白,使結構蛋白在體外實現自組裝、形成具有原病毒類似形態結構的中空顆粒。目前,用于制備VLPs的表達體系包括昆蟲桿狀病毒/昆蟲細胞表達體系(Bac-to-Bac)、植物細胞表達體系、哺乳動物細胞表達體系以及原核表達體系。其中昆蟲桿狀病毒/昆蟲細胞表達體系最為常用,只有少數病毒VLPs在原核表達體系中構建成功。FRENCH等[14]和王健偉等[15]分別通過昆蟲桿狀病毒/昆蟲細胞表達體系(Bac-to-Bac)成功制備了BTV-10、BTV-17和BTV-13 的VLPs,而THUENEMANN等[13,16]在植物細胞中共表達BTV-8 VP2、VP5、VP3和VP7這4種結構蛋白,成功制備出BTV-8 VLPs[13,16]。他們共同的特點均是通過構建雙表達重組病毒(VP2+VP5組合和VP3+VP7組合),然后讓這種病毒共感染真核細胞,實現4種蛋白在同一個細胞中共表達從而自組裝形成VLPs。由于共感染幾率相對較低,無法確保構成VLPs的4種蛋白能在同一個細胞中共表達,從而影響BTV VLPs形成的效率。為此。本研究對載體pFastBac-dual進行改造,使其具有4個多克隆位點(MSC)。雖然存在降低單個蛋白表達水平的可能,但通過這一方式能夠確保4種蛋白在同一細胞中共表達,從而有利于4種蛋白實現自組裝形成VLPs。
M.蛋白Marker;1.Sf9細胞蛋白;2.純化的BTV VLPs
M.蛋白Marker;1.純化的BTV VLPs;2.正常Sf9細胞對照
圖10 電鏡不同視野下觀察到的VLPs(40 000×)
通過透射電鏡觀察BTV VLPs的大小、形態和結構,發現存在多種中空顆粒形態共存的現象,有無突起、直徑在50 nm左右的中空顆粒(SCLPs),表面有非常明顯的突起;直徑為60~70 nm的中空顆粒(CLPs),外側壁明顯增厚(25 nm左右),表面突起不明顯;還有直徑為75~85 nm的中空顆粒(VLPs),以及少量形態和直徑介于CLPs和VLPs之間的中空顆粒。這一結果與王健偉等[15]在昆蟲細胞表達系統制備VLPs和THUENEMANN等[13,16]在植物細胞表達系統制備VLPs的結果相一致。這一現象揭示了BTV VLPs的結構和組裝過程,即首先VP3自組裝成亞核心顆粒(直徑最小、側壁最薄),VP7在其外層形成突起結構(CLPs的表面突起),VP2和VP5共同構成CLPs的外層結構(側壁增厚、直徑最大,同時VP2和VP5蛋白在最外層從而遮掩住了VP7形成的突起結構)。而THUENEMANN等[13,16]利用CPMV-HT 和相應的pEAQ載體系統(non-replicating system)在植物細胞中制備BTV VLPs,結果發現降低VP3表達水平可提高植物細胞中VLPs組裝效率。由于本研究所用的Bac-to-Bac系統屬于可復制體系,無法控制VP3的表達水平,因此并未對其結果進行驗證。
本研究成功地構建了可共表達BTV-16的4種結構蛋白VP2、VP5、VP3和VP7的昆蟲桿狀病毒,并制備出結構完整的BTV-16 VLPs,為下一步BTV-16 VLPs免疫效果評價以及相關疫苗的研制打下基礎。
