編碼大鼠ENPP1蛋白的cDNA克隆及重組腺相關病毒構建
2021-03-02 01:35:38狄楓吳秀娟沈水娟郭波堯
溫州醫科大學學報 2021年1期
狄楓,吳秀娟,沈水娟,郭波堯
紹興市人民醫院,浙江 紹興 312000,1.呼吸科;2.腎內科
血管鈣化是動脈粥樣硬化、糖尿病血管病變、慢性腎臟病(chronic kidney dielace,CKD)等多種疾病的共同病理生理過程。在腎小球濾過率降低的CKD患者中,血管鈣化發生率可達89%[1]。CKD患者的血管鈣化并不是簡單的鈣磷被動沉積過程,而是血管平滑肌細胞(vascular smooth muscle cells,VSMC)在CKD環境中經歷凋亡和囊泡形成,并向成骨樣細胞表型轉換,進而誘導鈣化蛋白表達,基質形成并吸收鈣磷沉積的主動過程[2]。在這一過程中,血管鈣化是鈣化促進因素和抑制因素不平衡的結果。我們的前期研究發現,維生素D3 聯合尼古丁(vitamin D3 and nicotine,VDN)血管鈣化模型的大鼠在停止藥物干預后,血管鈣化可發生自行消退,在這一過程中,外核苷酸焦磷酸酶/磷酸二酯酶1(ectonucleotide pyrophosphatase/phosphodiesterase 1,ENPP1)基因高表達[3]。為此,本研究通過采用克隆編碼ENPP1蛋白cDNA并構建其重組腺相關病毒(adeno-associated virus vector,AVV),以便為后續研究其在血管鈣化抑制中的作用及基因治療提供技術條件。
1 材料和方法
1.1 試劑和儀器 主要試劑:DH5α感受態細胞(Takara,日本);高純度質粒小量中提試劑盒(北京天根生化有限公司);DNA引物合成(上海捷瑞公司);Qiagen大規模質粒抽提試劑盒(Qiagen,德 國);DNA內切酶和T4 DNA連接酶(DNA marker,上海賽默飛);DNA凝膠回收試劑盒(Omega,美國);PCR產物純化試劑盒(Axygen,美國);Pav-fh AAV9載體(Vigenebio,美國)。主要儀器:全波長酶標儀、超微量高精度分光光度計、落地式培養搖床(美國Thermo公司);超速離心機(日本Hitachi);pH計(德國Mettler toledo公司);倒置顯微鏡(日本 Olympus);實時熒光定量擴增儀(美國Bio-Rad);熒光顯微鏡(日本Nikon);CO2培養箱(日本Sanyo); 超凈工作臺、生物安全柜(蘇州安泰空氣技術有限公司)。
1.2 方法
1.2.1 克隆大鼠ENPP1基因大片段:提取8周齡雄性SD大鼠的血管中膜平滑肌組織。確定大鼠目的基因ENPP1,NM053535.1;序列全長2 721 bp。設計引物:正向GCGTGAATTCGCCACCATGGAGCGCGACG;反向 GCGTACGCGTGTCTTCTTGGCTGAAGATTGGT。PCR引物由上海生工生物工程股份有限公司合成。模板:大鼠血管中膜平滑肌原代細胞cDNA。收到引物后瞬時離心,將引物干粉加水稀釋成100 μmol/L的母液,再轉入1.5 mL離心管,稀釋10倍成為PCR工作液。應用RT-PCR試劑盒反轉錄擴增ENPP1基因。RT-PCR反應條件為:首先42 ℃ 30 min,98 ℃變性3 min進行反轉錄,隨后98 ℃變性20 s,64 ℃退火30 s,72 ℃延伸90 s,循環35 次,再72 ℃延伸5 min,進行PCR擴增。擴增產物經1.0%的瓊脂糖凝膠電泳初步鑒定片段大小,膠回收PCR產物,利用DNA純化試劑盒將PCR產物進行純化。
1.2.2 構建克隆質粒:將純化PCR產物BamH I和EcoR I酶切后定向連接到pCDH-CMV-MCS-EF1-copGFPT2A-Puro(CD513B-1)載體中,構建pCDH-ENPP1重組質粒,將連接產物轉化感受態細胞E.coli DH5α構建克隆質粒。步驟如下:將連接產物與感受態細胞混勻后冰浴30 min,42 ℃熱激90 s,即置冰上 5 min,加入預熱至室溫的400 μL LB培養基,37 ℃ 恒溫搖床200 r/min培養1 h,離心棄上清液后用移液器混勻后均勻涂布于含氨芐青霉素抗性的LB平板上,37 ℃倒置恒溫培養箱培養過夜。挑取單菌落于37 ℃振蕩培養過夜后提取質粒,限制性內切酶鑒定后送上海生工生物工程股份有限公司進行DNA測序。
1.2.3 重組pAAV/ENPP1質粒的構建:用EcoR I/Xho I分別對腺相關病毒空載體pAAV-MCS和pCDHENPP1重組質粒進行雙酶切,反應結束后用1%瓊脂糖凝膠電泳檢測酶切目的條帶大小,膠回收目的基因酶切片段,T4 DNA連接酶將目的片段與載體片段22 ℃連接2 h,連接產物轉化大腸桿菌DH5α感受態細胞。將制備好的DH5α感受態置于冰浴中,待DH5α感受態細胞融化后,取1 μL連接產物于20 μL DH5α感受態細胞中,混勻后冰浴中靜置30 min。放入 42 ℃水浴鍋中40 s然后快速移至冰浴中,靜置 2 min。向離心管中加入200 μL無菌LB培養基混勻后置于搖床中37 ℃,200 r/min,振搖1 h。涂布到含氨芐青霉素抗性的固體培養基平皿中,37 ℃培養箱中培養過夜進行篩選。將pAAV-MCS空載體轉化大腸桿菌轉染VSMC作為陰性對照。
1.2.4 重組pAAV/ENPP1質粒的篩選與鑒定:經含氨芐青霉素抗性篩選,挑取單菌落于37 ℃振蕩培養過夜后提取質粒,挑取單菌落培養后提取質粒通過PCR和EcoR I/Xho I酶切鑒定陽性克隆,測序驗證,測序結果與Gen Bank中所報道目的基因序列進行比對分析。
1.2.5 病毒包裝:構建好的病毒載體和輔助質粒用Qiagen質粒抽提試劑盒進行大量抽提,保證濃度大于1 μg/μL,A260/A280 1.7~1.8,用以包裝病毒。準備HEK-293T細胞,提前1 d將HEK-293T細胞按照5.0×105細胞/孔的密度接種到孔板中,包裝時細胞密度85%~90%且細胞分布均勻,狀態良好。轉染前1~2 h給細胞換液為無血清的DMEM培養基(1% HEPES和1% P/S)。轉染:配制包裝mix,轉染試劑、包裝質粒、載體質粒、輔助質粒的比例為15:2:2:1。室溫靜置30 min。將靜置后的液體加到HEK-293T細胞中搖晃混勻。將細胞置于37 ℃,5% CO2培養箱中培養72 h后收集病毒。收集病毒:將細胞吹起,與培養基一并收到50 mL離心管中。離心分離細胞沉淀及上清液。將培養基上清液轉移到新管中,PGE8000沉淀過夜。離心去上清液,將細胞沉淀用PBS+0.001% PF68重懸。凍融1次后加入5 mmol/L NaCl 1 mL,渦旋混勻,將重懸液振蕩混勻后超聲至不黏稠。將超聲后的液體3 500×g離心30 min,收集上清液。純化:配置不同濃度的碘克沙醇,密度梯度離心。超速離心純化病毒。濃縮:收集病毒,置于超濾管中對病毒進行濃縮。吸10 μL病毒液進行滴度檢測及特異性檢測。吸取病毒液用96 孔板轉染HEK-293T細胞24 h。病毒滴度測定:腺相關病毒用蛋白酶K(5 μg/μL)處理破除病毒外殼,q-PCR法檢測病毒滴度。設立5個梯度稀釋濃度的質粒標準品的拷貝數,將超純水作為陰性對照。根據標準曲線獲得樣品拷貝數。病毒顆粒數(個/mL)=與標準品相對值×10 000。
1.2.6 重組腺相關病毒轉染VSMC:將目的細胞VSMC接種于96孔板,加入梯度稀釋病毒原液,轉染VSMC觀察細胞形態以摸索最佳轉染復數(MOI)。取正常生長的P4代VSMC,消化接種于6孔板,每孔2×105個細胞,待細胞密度達到70%~80%時進行病毒轉染,將最佳MOI病毒液加入VSMC,混勻,37 ℃小體積轉染96 h。
1.2.7 Western blot檢測VSMC ENPP1蛋白的表達:蛋白樣品制備:使用細胞裂解液,對VSMC進行裂解。然后BCA法測定蛋白濃度。測定562 nm處的吸光度值,根據標準曲線計算樣品的蛋白濃度。聚丙烯酰胺凝膠電泳制膠,電泳30 min。轉膜:將PVDF膜在甲醇中浸泡約30 s,再在轉移緩沖液浸泡10 min,與濾紙、凝膠一起放入緩沖液中200 mA穩流電轉移。封閉:PVDF膜去離子水洗滌后浸入封閉液,置搖床上室溫封閉1 h。孵育一抗:取出已封閉的PVDF膜,浸于1×TBST緩沖液中,洗滌5 min,然后移入含有一抗的孵育盒中,4 ℃孵育過夜。TBST緩沖液洗膜。 孵育二抗:將PVDF膜轉移到含二抗的抗體孵育盒孵育1 h后洗膜。ECL化學發光顯影:將PVDF膜置于保鮮膜上,取適量ECL試劑盒中等體積的A液和B液混合,混勻后加在膜的表面,移入凝膠成像分析儀中,化學光敏模式曝光顯影。
2 結果
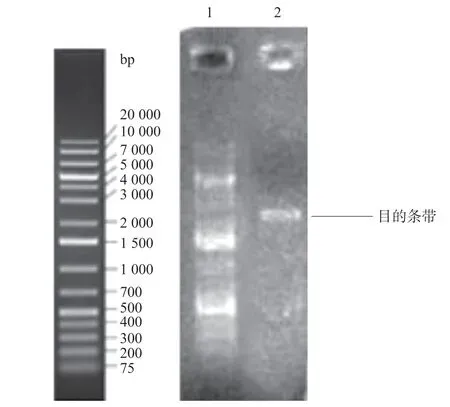
圖1 目的基因ENPP1 PCR產物的純化

圖2 酶切重組質粒載體
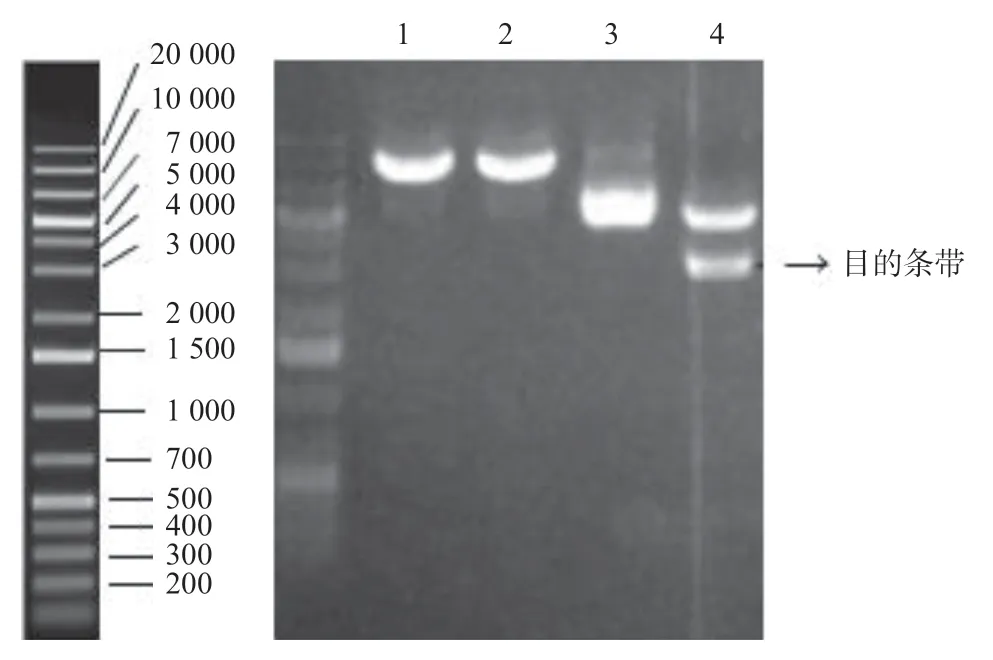
圖3 菌落PCR鑒定陽性克隆
2.1 大鼠ENPP1基因大片段的克隆與鑒定 在RTPCR反應中首先對PCR的退火溫度進行梯度篩選,發現在退火溫度為64 ℃時可以擴增出較特異的DNA片段,所得PCR產物大小約2 720 bp,與目的片段大小一致(見圖1)。隨后將該PCR產物與pCDH-CMV-MCSEF1-copGFP-T2A-Puro載體進行連接,酶切結果顯示目的片段成功克隆至pCDH載體中(見圖2),重組質粒載體轉化大腸桿菌DH5α感受態細胞,培養后挑取單 菌落提取質粒進行菌落PCR鑒定陽性克隆(見圖3)。2.2 重組pAAV/ENPP1質粒的構建與鑒定 經過EcoR I/Xho I酶切、連接、轉化和氨芐青霉素抗性篩選,LB平板上有數十個可疑克隆生長,提取質粒后經EcoR I/Xho I酶切和PCR擴增鑒定,共獲得3個陽性重組質粒pAAV/ENPP1。測序結果顯示,3個質粒與基因庫BLAST比對(見圖4)為序列一致的大鼠ENPP1全長cDNA,同源性100%,E值達8×10-98,且開放讀碼框完全正確(見圖5),表明大鼠VSMC ENPP1重組腺相關病毒載體構建成功。重組pAAV/ENPP1質粒用HEK-293T細胞包裝。重組腺相關病毒滴度測定:q-PCR檢測病毒滴度,質粒標準品的拷貝數,5個梯度稀釋濃度分別為4.51×1011、3.73×1010、3.26× 109、5.57×108、4.31×107,得出標準曲線(見圖6),根據標準曲線計算標準品相對值。最終獲得ENPP1基因腺相關病毒滴度為2.79×1014v.g./mL。
2.3 rAAV/ENPP1轉染VSMC后ENPP1表達情況 病毒轉染組ENPP1蛋白表達較空白對照組高(見圖7)。
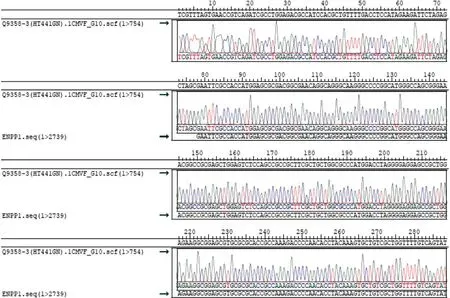
圖4 目的基因序列測序圖
3 討論
ENPP1屬于ENPP酶家族,是一種具有核苷酸焦磷酸酶和磷酸二酯酶活性的II型跨膜糖蛋白。ENPP1基因突變在人類可導致一種罕見的嚴重常染色體隱性遺傳疾病——嬰兒泛發性動脈鈣化(generalized arterial calcification in infancy,GACI),特點是發生廣泛中型肌性動脈鈣化,病情進展迅速,多數患兒因心衰死于嬰兒期[4]。敲除大鼠ENPP1基因可制作GACI動物模型,而給ENPP1基因敲除大鼠皮下注射ENPP1融合蛋白(小鼠ENPP1細胞外結構域融合人IgG1 Fc段)可明顯減輕GACI模型小鼠的血管鈣化,降低死亡率,以及心肌梗死及后遺癥發生率[5],提示其在抑制血管鈣化中具有非常重要的作用。
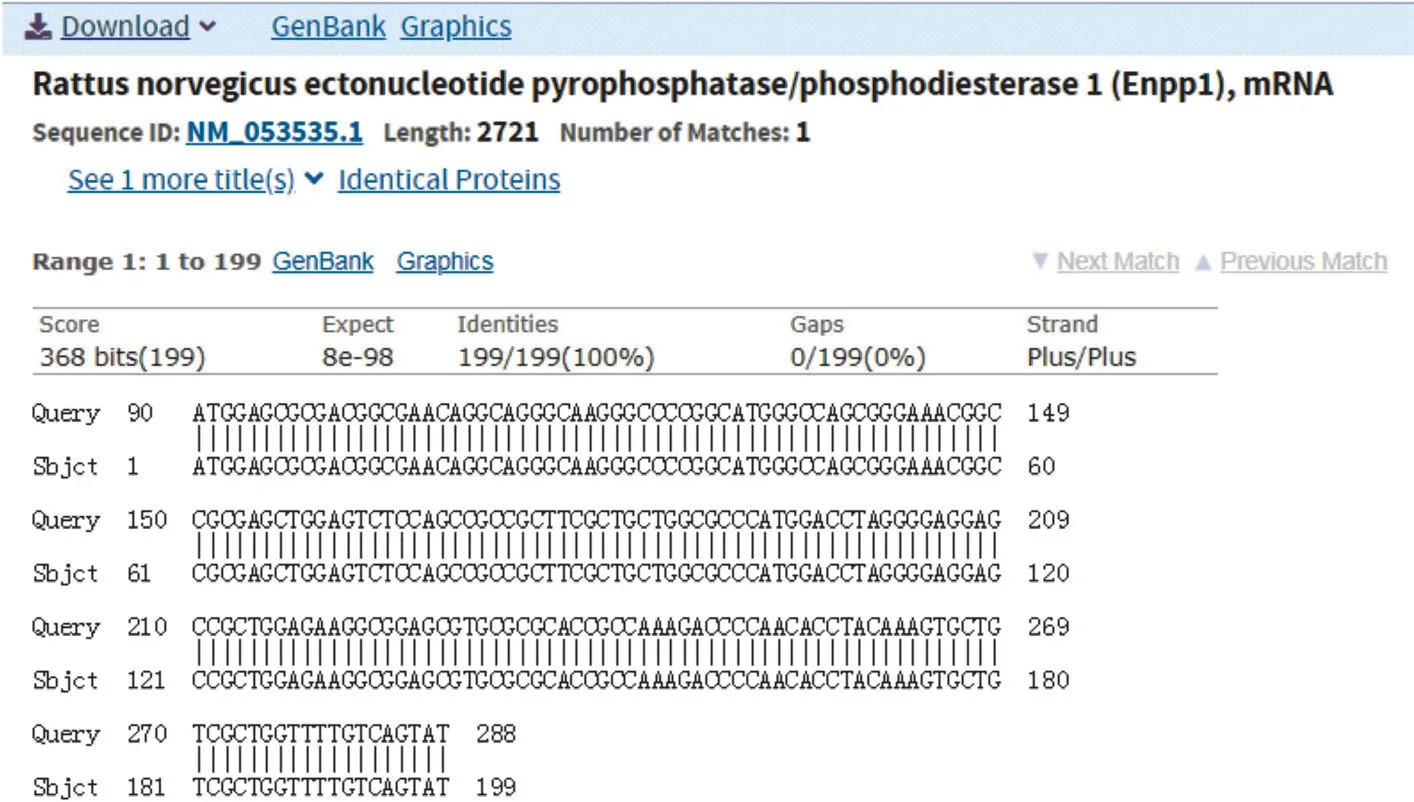
圖5 基因序列BLSAT比對
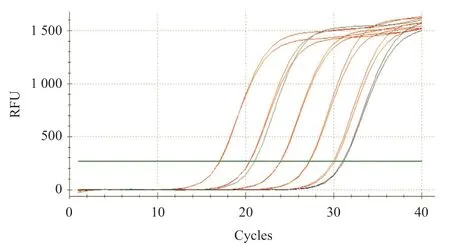
圖6 ENPP1病毒液qPCR擴增曲線圖
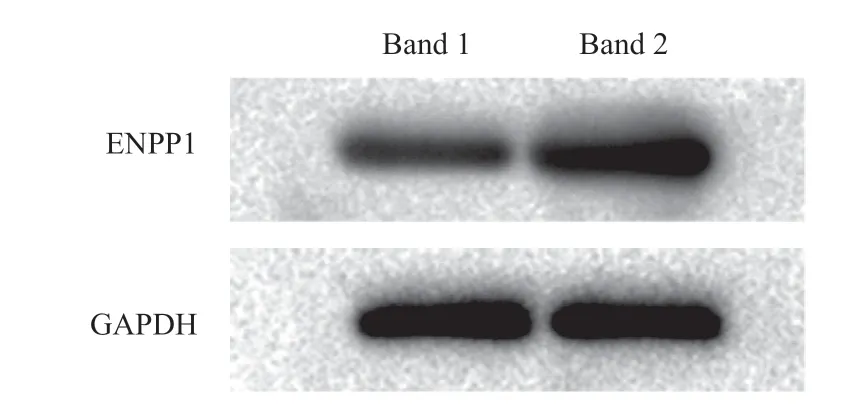
圖7 ENPP1 Western blot條帶圖
ENPP1廣泛參與核苷酸循壞、焦磷酸水平調控、磷脂信號調控、胰島素受體調控及胞外激酶活性。ENPP1可通過水解胞外ATP中的5,磷酸二酯鍵生成AMP和焦磷酸(PPi),后者可防止非晶態磷酸鈣的成核,還能抑制晶體向羥基磷灰石方向生長,并通過與羥基磷灰石表面結合形成不規則晶體而抑制晶體生長,有效預防病理性組織鈣化。ENPP1Q等位基因與主動脈弓鈣化增加有關[6]。有研究認為ENPP1可作為評價非糖尿病患者冠心病的生物學標志物[7]。因此我們認為ENPP1在慢性腎臟病、心血管病所致的血管鈣化中也起著重要作用,是一個礦化調節的重要基因。目前,臨床上對于血管鈣化尚無特異性的治療方法,或許可通過ENPP1基因治療對血管鈣化進行有效救治。
此外,病毒載體的選擇也是基因治療的關鍵。近年來,國內外對轉基因治療血管性疾病進行了深入研究。在這些實驗中,常選擇腺病毒載體,其優點是轉染效率高,缺點是轉染的基因表達時間較為短暫,不適合血管鈣化這種慢性疾病的防治。與腺病毒相比,AAV攜帶的外源基因可以長時間穩定表達,而且它引起的免疫反應輕微。AAV屬于細小病毒家族,是一類無包膜的復制缺陷病毒,可以通過基因工程技術將DNA傳遞給靶細胞,是一個非常穩定的載體,能夠承受廣泛的溫度和pH變化,幾乎沒有喪失活性[8],已被證明是最有希望的人類基因治療的病毒載體之一[9]。重組AAV(rAAV)是人工改造的AAV,AAV中與復制和包裝相關的erp和cap基因被替換為ITRs間的基因,使rAAV具有4.1~4.9 kb的包裝容量。rAAV由多個質粒組成,包括cis質粒、輔助質粒、erp/cap質粒,它們之間不具有同源性序列,故rAAV不具有復制能力,復制過程依賴于宿主細胞的聚合酶,因此避免通過重組而恢復出野生型病毒,宿主的免疫源性被降到最低。rAAV進入細胞后,作為附加的DNA能穩定存在,因此能長期表達外源基因,基因表達可持續數周至數月。rAAV是基因傳遞和表達的一個重要工具,本研究應用rAAV成功包裝出具有2 721 bp的ENPP1大容量基因片段。目前AAV有12種血清型,不同的血清型有不同的衣殼蛋白結構、序列和組織特異性,因而其識別與結合細胞表面受體也相應有很大的差別。根據不同血清型轉染細胞類型和轉染效率,AAV-9轉染原代細胞具有獨特的能力,AAV-1和AAV-9在向骨骼肌和心肌傳遞基因方面表現得非常有效[9],AAV-2/9作為載體可在不引起炎癥的情況下有效地傳遞冠狀動脈和外周動脈平滑肌細胞中的基因[10]。本實驗選用AAV-9血清型作為載體轉染VSMC,Western blot顯示轉染目的基因后目的蛋白ENPP1高表達,提示AAV-9血清型可作為研究VSMC基因表達的載體。
總之,本研究選用AAV載體AAV-9 作為研究ENPP1的載體,在獲得編碼大鼠VSMC ENPP1的全長cDNA后,通過分子生物學技術將其裝載至AAV載體rAAV-MCS中,最終通過酶切鑒定和DNA測序證實成功構建了序列信息完全正確的ENPP1重組AAV表達載體。
