腺苷三磷酸對(duì)人外周血單個(gè)核細(xì)胞hBD-2表達(dá)的影響及其分子機(jī)制探討
2021-03-01 02:57:24張夢(mèng)潔
山東醫(yī)藥 2021年5期
張夢(mèng)潔
上海中醫(yī)藥大學(xué)附屬普陀醫(yī)院,上海200062
人β防御素2(hBD-2)是第一個(gè)被發(fā)現(xiàn)的具有 可誘導(dǎo)表達(dá)特性的防御素[1],廣泛分布于皮膚、支氣管肺部、子宮等部位,尤其以支氣管肺部表達(dá)最多。其具有良好的殺菌能力和抗病毒、抗真菌、抗腫瘤等多種生物學(xué)活性,能趨化多數(shù)免疫細(xì)胞,在固有免疫和特異性免疫中均有重要功能,因此在肺部炎癥性疾病中發(fā)揮重要作用。在自然條件下,hBD-2表達(dá)水平極低或不表達(dá);在受到外界因素刺激誘導(dǎo)后,則在各種組織黏膜細(xì)胞中表達(dá)并上調(diào),發(fā)揮其抗微生物和調(diào)節(jié)免疫的功能[2]。腺苷三磷酸(ATP)是一種輔酶,通過(guò)多種蛋白功能為機(jī)體多種合成反應(yīng)提供所需能量,增強(qiáng)機(jī)體組織及細(xì)胞的代謝活性,因而其對(duì)治療各種疾病均有較強(qiáng)的針對(duì)性。目前,ATP已被廣泛應(yīng)用于細(xì)菌、病毒、球蟲(chóng)等引起的呼吸、消化及生殖系統(tǒng)疾病的臨床輔助治療。實(shí)驗(yàn)研究發(fā)現(xiàn),ATP可以上調(diào)大鼠防御素2(mBD-2)的表達(dá),但ATP是否可以上調(diào)hBD-2的表達(dá)水平目前鮮有文獻(xiàn)報(bào)告。2016年9月—2017年7月,我們應(yīng)用ATP體外刺激人外周血單個(gè)核細(xì)胞(PBMCs),觀(guān)察其是否可誘導(dǎo)hBD-2表達(dá),并探討其可能的分子機(jī)制。
1 材料與方法
1.1 主要實(shí)驗(yàn)試劑 PBMCs分離液(Ficoll)及ATP干粉均購(gòu)自美國(guó)Sigma公司,胎牛血清購(gòu)自美國(guó)BI公司,青霉素鏈霉素溶液(雙抗)、RPMI-1640培養(yǎng)液、HBSS和PBS緩沖液均購(gòu)自美國(guó)HyClone公司,臺(tái)盼蘭購(gòu)自美國(guó)Gibco公司,人白細(xì)胞介素1β(IL-1β)和hBD-2酶聯(lián)免疫吸附(ELISA)測(cè)定試劑盒均購(gòu)自武漢Elabscience公司。
1.2 實(shí)驗(yàn)方法
1.2.1 標(biāo)本來(lái)源與收集 經(jīng)醫(yī)院倫理委員會(huì)批準(zhǔn),收集健康志愿者20名,男10例、女10例,年齡(35.2±10.5)歲,漢族,均已簽署知情同意書(shū)。無(wú)菌抽取健康志愿者外周血9 mL于EDTA管抗凝,2 h內(nèi)進(jìn)行PBMCs的分離。
1.2.2 人PBMCs的分離 采用Ficoll-Hypaque密度梯度離心法。用無(wú)菌吸管各取3 mL血液于3只15 mL離心管中,并各加入3 mL等量HBSS液進(jìn)行1∶1稀釋?zhuān)p輕上下吹打,充分混勻;用10 mL注射器吸取搖勻的Ficoll 3 mL于15 mL離心管內(nèi),避免管壁殘留分離液(Ficoll∶稀釋前全血∶HBSS=1∶1∶1);將離心管傾斜45°,槍頭緊貼管壁將稀釋全血沿管壁緩慢加于Ficoll液面上,注意保持兩者界面分層清晰。在室溫(20℃)下,用水平離心機(jī)以離心半徑10 cm、400 g離心30 min。離心后管內(nèi)分為三層,在第一、二層交界處有一狹窄帶,呈白色云霧狀,以單個(gè)核細(xì)胞為主。用移液槍去除最上層,小心吸出PBMC層(1~2 mL),移至另一15 mL離心管中。加入6~8倍體積PBS洗滌2次,室溫下以離心半徑10 cm、400 g離心5 min,留取沉淀。用含10%胎牛血清的RPMI-1640培養(yǎng)液1 mL重懸細(xì)胞,并配成單個(gè)細(xì)胞懸液。將重懸了PBMCs的RPMI-1640培養(yǎng)液吹均勻,然后吸取少量懸液沿蓋片邊緣緩緩滴入,顯微鏡下觀(guān)察并計(jì)數(shù)。用移液器吸取10 μL細(xì)胞懸液到離心管中,再加入0.4%臺(tái)盼藍(lán)10 μL染液,于顯微鏡下計(jì)數(shù)PBMCs,活細(xì)胞百分比達(dá)95%以上,達(dá)到實(shí)驗(yàn)要求。
1.2.3 PBMCs分組與ATP干預(yù) 在超凈臺(tái)內(nèi),用含10%胎牛血清的RPMI-1640培養(yǎng)基調(diào)整細(xì)胞密度為1×106/mL;將細(xì)胞懸液接種于96孔培養(yǎng)板,每孔100 μL;將PBMCs分為六組,每組設(shè)3個(gè)復(fù)孔。空白組不處理,0、25、50、100、200 μmol/L ATP組分別加入 HBSS液和 25、50、100、200 μmol/L ATP 100 μL,于37℃、5%CO2及相對(duì)濕度為95%的培養(yǎng)箱中培養(yǎng)。分別于培養(yǎng)12、24 h后終止培養(yǎng),將各組培養(yǎng)后的細(xì)胞懸液轉(zhuǎn)移至1.5 mL無(wú)菌離心管中,標(biāo)記編號(hào)后以離心半徑10 cm、1 500 r/min低溫離心10 min,留取適量培養(yǎng)上清液置-80℃冰箱內(nèi)保存。
1.2.4 PBMCs培養(yǎng)上清液hBD-2、IL-1β檢測(cè) 收集培養(yǎng)上清液,以離心半徑10 cm、3 000 r/min離心5 min,取上清液,采用雙抗體夾心ELISA法檢測(cè)hBD-2和IL-1β,具體步驟按試劑盒要求。即加50 μL待測(cè)樣品及系列濃度的標(biāo)準(zhǔn)溶液在細(xì)胞因子單抗包被的酶標(biāo)板上,室溫孵育使細(xì)胞因子結(jié)合到酶標(biāo)板上,再加入二抗孵育結(jié)合;最后加底物液顯色30 min,用酶標(biāo)儀在450 nm波長(zhǎng)處測(cè)OD值并計(jì)算hBD-2和IL-1β含量。
1.3 統(tǒng)計(jì)學(xué)方法 采用SPSS22.0統(tǒng)計(jì)軟件。所有計(jì)量數(shù)據(jù)以-x±s表示,多組間比較行單因素方差分析,組間兩兩比較行LSD-t檢驗(yàn),組內(nèi)不同時(shí)間點(diǎn)的比較行重復(fù)測(cè)量方差分析,相關(guān)關(guān)系采用Person相關(guān)性分析。P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
2.1 各組PBMCs培養(yǎng)上清液中hBD-2水平比較 見(jiàn)表1。
2.2 各組PBMCs培養(yǎng)上清液中IL-1β水平比較 見(jiàn)表2。
2.3 ATP作用后PBMCs培養(yǎng)上清液中hBD-2與IL-1β水平的關(guān)系 經(jīng)ATP作用12、24 h時(shí),PBMCs培養(yǎng)上清液中hBD-2與IL-1β水平均呈正相關(guān)(r分別為0.494、0.725,P均<0.01)。
3 討論
hBD-2是防御素家族中非常重要的成員,主要表達(dá)于皮膚、支氣管等各種黏膜上皮細(xì)胞,外周血中的中性粒細(xì)胞和單核細(xì)胞在致炎因子刺激下也可以表達(dá) hBD-2[3-4]。在高濃度時(shí),hBD-2 對(duì)病原微生物具有直接殺傷作用;而低濃度時(shí),hBD-2就只有趨化性,它可趨化募集樹(shù)突狀細(xì)胞、T淋巴細(xì)胞、白細(xì)胞、肥大細(xì)胞和巨噬細(xì)胞等,使這些細(xì)胞聚集在感染部位、誘導(dǎo)分化成熟、促進(jìn)細(xì)胞因子產(chǎn)生;而這些細(xì)胞及細(xì)胞因子反過(guò)來(lái)又影響hBD-2的誘導(dǎo)表達(dá),從而對(duì)機(jī)體的免疫狀態(tài)產(chǎn)生影響。hBD-2也可通過(guò)激活G蛋白偶聯(lián)受體和磷脂酶C途徑還能與肥大細(xì)胞表面兩種特異性受體結(jié)合,從而誘導(dǎo)肥大細(xì)胞聚集到炎癥部位并活化、脫顆粒、釋放組胺,參與速發(fā)型超敏反應(yīng)[5]。研究顯示,hBD-2是Toll樣受體(TLR)的內(nèi)源性配體,通過(guò)TLR2、TLR4、CC趨化因子受體6(CCR6)等受體介導(dǎo),在IL-1相關(guān)蛋白激酶(IRAK)和腫瘤壞死因子受體相關(guān)因子6(TRAF6)等信號(hào)轉(zhuǎn)導(dǎo)分子的協(xié)同作用下,觸發(fā)細(xì)胞內(nèi)信號(hào)級(jí)聯(lián)反應(yīng),激活NF-κB,活化T淋巴細(xì)胞,觸發(fā)強(qiáng)有力的特異性免疫應(yīng)答[6]。YANG等[7]發(fā)現(xiàn),hBD通過(guò)CCR6途徑誘導(dǎo)的未成熟樹(shù)突狀細(xì)胞(iDC)的遷移,在hBD-2的刺激下通過(guò)CCR6促進(jìn)iDC遷移,促進(jìn)抗原提呈作用,激活T細(xì)胞,引發(fā)免疫應(yīng)答。
表1 各組PBMCs培養(yǎng)上清液中hBD-2水平比較()
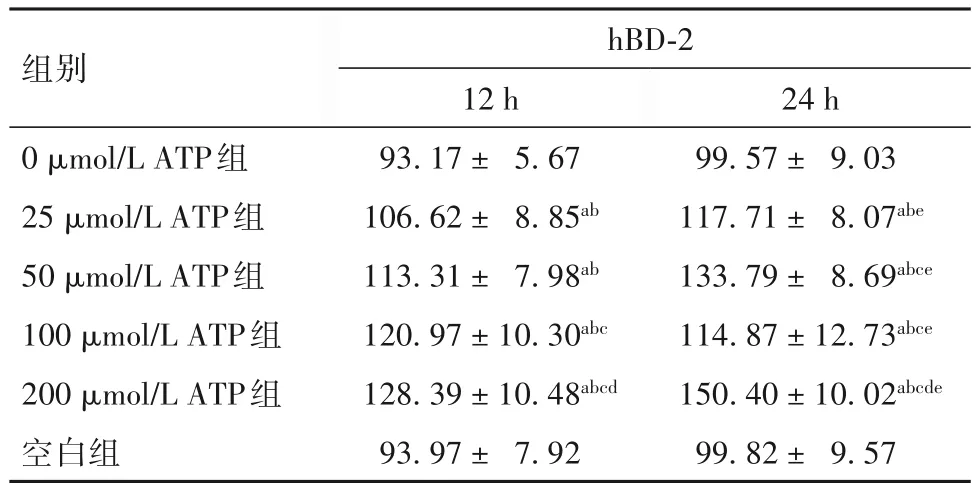
表1 各組PBMCs培養(yǎng)上清液中hBD-2水平比較()
注:與空白組比較,aP<0.01;與0 μmol/L ATP組比較,bP<0.01;與 25 μmol/L ATP組比較,cP<0.01;與 50 μmol/L ATP組比較,dP<0.01;與同組培養(yǎng)12 h比較,eP<0.01。
images/BZ_15_234_402_1193_520.png99.57± 9.03 117.71± 8.07abe 133.79± 8.69abce 114.87±12.73abce 150.40±10.02abcde 99.82± 9.57 0 μmol/L ATP組25 μmol/L ATP組50 μmol/L ATP組100 μmol/L ATP組200 μmol/L ATP組空白組93.17± 5.67 106.62± 8.85ab 113.31± 7.98ab 120.97±10.30abc 128.39±10.48abcd 93.97± 7.92
表2 各組PBMCs培養(yǎng)上清液中IL-1β水平比較()
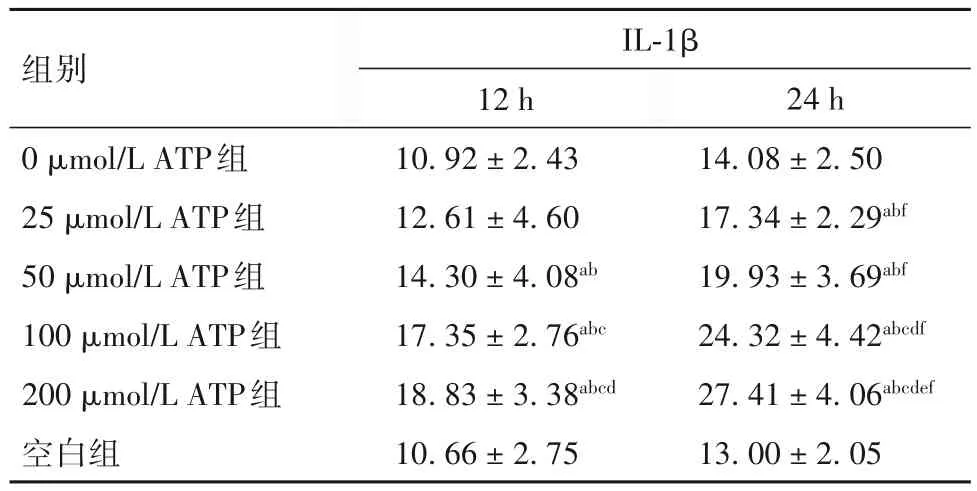
表2 各組PBMCs培養(yǎng)上清液中IL-1β水平比較()
注:與空白組比較,aP<0.01;與0 μmol/L ATP組比較,bP<0.01;與 25 μmol/L ATP組比較,cP<0.01;與 50 μmol/L ATP組比較,dP<0.01;與100 μmol/L ATP組比較,eP<0.01;與同組培養(yǎng)12 h比較,fP<0.01。
24 h 14.08±2.50 17.34±2.29abf 19.93±3.69abf 24.32±4.42abcdf 27.41±4.06abcdef 13.00±2.05組別0 μmol/L ATP組25 μmol/L ATP組50 μmol/L ATP組100 μmol/L ATP組200 μmol/L ATP組空白組IL-1β 12 h 10.92±2.43 12.61±4.60 14.30±4.08ab 17.35±2.76abc 18.83±3.38abcd 10.66±2.75
有研究發(fā)現(xiàn),ATP還具有細(xì)胞間信息傳遞功能[8],可提高細(xì)胞內(nèi)鈣離子水平,促進(jìn)鈣離子參與調(diào)節(jié)細(xì)胞的多種生物學(xué)功能。鈣非特異性調(diào)節(jié)劑ATP能通過(guò)與特殊受體(P2-嘌呤受體)結(jié)合,促進(jìn)磷酸肌醇激酶的水解,從而引起鈣動(dòng)員、胞內(nèi)鈣升高,導(dǎo)致細(xì)胞的特殊反應(yīng);這些反應(yīng)依據(jù)細(xì)胞類(lèi)型不同而異,并且與細(xì)胞的功能有關(guān)[9]。hBD-2在自然條件下表達(dá)水平極低或不表達(dá),當(dāng)受到刺激誘導(dǎo)后則在各種組織黏膜細(xì)胞中表達(dá)并上調(diào),發(fā)揮其抗微生物和調(diào)節(jié)免疫的功能[2]。hBD-2的可誘導(dǎo)表達(dá)特性,為利用某種人工方式調(diào)控其表達(dá)提供了理論依據(jù)。PBMCs為免疫活性細(xì)胞的集合體,包括T淋巴細(xì)胞、B淋巴細(xì)胞、NK細(xì)胞、單核細(xì)胞等免疫活性細(xì)胞,尤其T淋巴細(xì)胞,在機(jī)體免疫反應(yīng)中扮演重要作用。hBD-2能趨化多數(shù)免疫細(xì)胞,因而在固有免疫和特異性免疫中均起著重要作用。
為探索人工促進(jìn)hBD-2表達(dá)上調(diào)的方法,以進(jìn)行體內(nèi)研究且能應(yīng)用于臨床,并進(jìn)一步探討hBD-2與機(jī)體免疫狀態(tài)的關(guān)系。本研究先提取人PBMCs,用培養(yǎng)基調(diào)節(jié)至一定濃度后接種于培養(yǎng)板中,分別加入不同濃度的ATP液于培養(yǎng)箱中培養(yǎng),分別于12、24 h收集細(xì)胞培養(yǎng)上清液,用ELISA法檢測(cè)其上清中的hBD-2及IL-1β。結(jié)果發(fā)現(xiàn),與空白組和0 μmol/L ATP組比較,經(jīng)不同濃度ATP組處理后,PBMCs細(xì)胞培養(yǎng)上清中hBD-2及IL-1β表達(dá)水平出現(xiàn)不同程度的上調(diào),差異有統(tǒng)計(jì)學(xué)意義;且隨著ATP濃度適當(dāng)?shù)脑黾蛹白饔脮r(shí)間的延長(zhǎng),細(xì)胞培養(yǎng)上清液中hBD-2及IL-1β的表達(dá)水平可進(jìn)一步升高。本研究結(jié)果進(jìn)一步證明了ATP可以誘導(dǎo)人體內(nèi)hBD-2和IL-1β表達(dá),并可通過(guò)人體免疫細(xì)胞發(fā)揮作用,在增強(qiáng)機(jī)體免疫方面起極為重要的作用。本研究結(jié)果提示,經(jīng)ATP處理后,PBMCs培養(yǎng)上清液中hBD-2與IL-1β的表達(dá)水平呈正相關(guān),這與SUN等[10]發(fā)現(xiàn)大鼠肺組織中mBD-2與IL-1β表達(dá)呈正相關(guān)這一觀(guān)點(diǎn)相符。目前的研究表明,hBD-2的誘導(dǎo)表達(dá)可能是通過(guò)多種信號(hào)途徑實(shí)現(xiàn)的,核轉(zhuǎn)錄因子κB(NF-κB)信號(hào)通路被外來(lái)刺激信號(hào)激活可能是hBD-2基因激活的機(jī)制之一,hBD-2的表達(dá)是通過(guò)激活NF-κB、在IL-1β及TNF-α參與下實(shí)現(xiàn)的[11]。也有研究表明,ATP是促進(jìn)巨噬細(xì)胞中IL-1β成熟與分泌的重要內(nèi)源性刺激物[12-13]。結(jié)合本研究可以推測(cè),ATP作用于PBMCs后使細(xì)胞內(nèi)鈣離子濃度增高,導(dǎo)致NF-κB信號(hào)通路激活,從而誘導(dǎo)細(xì)胞因子如IL-1β和hBD-2表達(dá),使細(xì)胞內(nèi)IL-1β和hBD-2的分泌增加;而hBD-2反過(guò)來(lái)趨化免疫細(xì)胞,促進(jìn)產(chǎn)生細(xì)胞因子如IL-1β,這些細(xì)胞因子反過(guò)來(lái)又影響hBD-2的誘導(dǎo)表達(dá),兩者相互影響;但ATP也有可能通過(guò)直接刺激PBMCs分泌IL-1β,進(jìn)而介導(dǎo)hBD-2的表達(dá)上調(diào)。當(dāng)然,激活hBD-2基因表達(dá)還存在其他機(jī)制,如IL-6、JAK/STAT途徑等。有研究認(rèn)為,其他部位的上皮細(xì)胞誘導(dǎo)hBD-2表達(dá)的信號(hào)轉(zhuǎn)導(dǎo)機(jī)制與氣道上皮細(xì)胞可能不同[14-15]。例如 KRISANAPRAKORNKIT 等[16]發(fā)現(xiàn),通過(guò)絲裂原素活化蛋白激酶通路可以實(shí)現(xiàn)齒齦上皮細(xì)胞hBD-2的表達(dá),而不是NF-κB激活通路。MOON等[17]用前炎癥細(xì)胞因子IL-1α刺激中耳上皮細(xì)胞表達(dá)hBD-2,發(fā)現(xiàn)這是通過(guò)Raf-MEK1/2-ERK依賴(lài)性信號(hào)通路發(fā)生的;但MCDERMOTT等[18]在角膜上皮細(xì)胞用IL-1β刺激hBD-2的表達(dá)時(shí),發(fā)現(xiàn)此過(guò)程既有NF-κB信號(hào)通路,也有絲裂原蛋白激酶信號(hào)通路。由此可見(jiàn),各種刺激因素對(duì)hBD-2的誘導(dǎo)表達(dá)可能是通過(guò)多種信號(hào)途徑實(shí)現(xiàn)的。ATP刺激PBMCs誘導(dǎo)hBD-2表達(dá)的具體機(jī)制和途徑,尚需進(jìn)一步研究。ATP作為臨床常用藥物,每日使用劑量可達(dá)100~200 mg。本研究提示,25 μmol/L ATP即可誘導(dǎo)hBD-2表達(dá)上調(diào),這與臨床上ATP使用劑量相比是極其微量的,這為肺部感染患者臨床上使用小劑量ATP來(lái)誘導(dǎo)機(jī)體hBD-2的表達(dá)并輔助抗感染提供了依據(jù)。
猜你喜歡
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評(píng)價(jià)·高一版(2020年6期)2020-11-02 02:45:24
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年12期)2019-05-21 02:55:32
中國(guó)生殖健康(2019年3期)2019-02-01 06:12:26
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
海軍航空大學(xué)學(xué)報(bào)(2015年3期)2015-11-11 17:20:00
中國(guó)火炬(2010年8期)2010-07-25 11:34:30

- 山東醫(yī)藥的其它文章
- HPV E6/E7 mRNA檢測(cè)用于絕經(jīng)后未明確診斷意義的不典型鱗狀上皮細(xì)胞分流篩查的價(jià)值
- 不同濃度醫(yī)用三氧自體血回輸治療急性痛風(fēng)性關(guān)節(jié)炎效果比較
- 沙利度胺對(duì)中晚期食管癌患者調(diào)強(qiáng)放化療效果及血清miR-451、miR-21水平的影響
- 心痛定聯(lián)合拉貝洛爾治療妊娠期高血壓疾病效果及患者血清IGF-1、HSP70水平變化
- 右美托咪定對(duì)體外循環(huán)下心臟瓣膜置換術(shù)患者多臟器損傷及細(xì)胞免疫功能的影響
- 降脂通絡(luò)軟膠囊聯(lián)合大劑量辛伐他汀對(duì)老年穩(wěn)定性冠心病患者脂代謝指標(biāo)的影響