海南島熱帶低地雨林2種攀緣竹的葉片功能性狀差異*
2021-02-12 05:58:02徐瑞晶劉廣路梁昌強孔祥河
林業科學 2021年12期
徐瑞晶 胡 璇 劉廣路 郭 雯 梁昌強 孔祥河
(1.國際竹藤中心 北京 100102; 2.海南省甘什嶺自然保護區管理站 三亞 572000)
植物為適應不同生境,在葉片光合特性、解剖結構和形態性狀上表現出不同的適應策略(張詠梅等, 2019; 寇萌等, 2019; 鐘巧連等, 2018)。葉片是植物進行光合作用、呼吸作用等生理代謝活動的重要器官,其變化特征最能反映植物的環境適應性(張詠梅等, 2019; Gaoetal., 2017; 王勛陵等, 1989)。熱帶季雨林植物通過調節葉片凈光合速率,積累更多干物質適應霧涼季低溫環境(栗忠飛等, 2012),常綠植物通過調節氣孔狀態,提高光合水分利用效率維持葉片四季常綠(白坤棟等, 2010)。在逆境脅迫中的植物通過調節葉片水勢、蒸騰速率和水分利用效率等光合生理因子,以適應季節更替和脅迫(鄧彭艷等, 2010; Inoueetal., 2017)。單子葉植物通過增大葉的表皮和上角質層厚度等保護組織厚度來保護葉肉組織,同時依靠泡狀細胞和較大的維管束等特殊解剖結構適應水分條件差、強光照的環境(寇萌等, 2019)。氣孔密度體現植物對環境(強光、高溫、大風等)的響應,在高光照強度或低水分利用率時葉片的氣孔密度較高(Rossattoetal., 2012); 發達的維管組織不僅利于提高水分利用效率,同時也是對葉片干旱和強風環境的適應(史剛榮等, 2007)。如內蒙古高原草地植物依靠小而密的氣孔,可快速有效地利用水分; 而青藏高原草地植物氣孔大而疏,將更多能量用于抵抗低溫脅迫(王媛媛等, 2018)。植物在長期適應環境過程中,通過內部不同功能之間的協同進化,形成能適應環境變化的形態和功能,葉功能性狀是反映這種適應策略的有效指標,如普定喀斯特地區植物有較低的葉面積和比根長度,較高的葉干物質含量和葉組織密度,這是其適應干旱貧瘠的喀斯特環境的重要生態策略(鐘巧連等, 2018)。表型可塑性是植物適應異質生境的重要生態對策,表型可塑性高的物種能適應更寬的環境變化范圍,能在不利條件下優化利用資源(Narayan, 2012; 施建敏等, 2014; 梁寬等, 2017)。
竹子葉片功能性狀及適應性相關研究已有報道,但只涉及直立竹,未涉及藤本狀的攀緣竹。直立竹主要集中在勃氏甜龍竹(Dendrocalamusbrandisii)、黃條金剛竹(Pleioblastuskongosanensisf.aureostriatus)、黃竹(D.membranaceus)等引種后的生長指標和抗性指標評價(邱爾發等, 2006; 夏海濤等, 2016; 唐紅等, 2019),以及毛竹(Phyllostachysedulis)的可塑性生長(施建敏等, 2014; 曹永慧等, 2016; 劉廣路等, 2017),還有慈竹(Bambusaemeiensis)、八月竹(Chimonobambusaszechuanensis)和短錐玉山竹(Yushaniabrevipaniculata)對海拔梯度的適應生長(蔣琴等, 2018)及淡竹(Phyllostachysglauca)在石灰巖生境的適應生長等方面(梁寬等, 2017)。
攀緣竹是竹亞科(Bambusoideae)的一個特殊類群,主要分布于熱帶、亞熱帶高溫多雨地區,以云南和海南島最為豐富。攀緣竹植株奇特,竿呈攀援狀或斜倚呈懸垂狀,類似藤本植物,可作特色園林和立體綠化用竹,具有較高的觀賞價值。攀緣竹是海南島低地雨林的重要組成部分,具有較高的科學研究價值(胡璇等, 2018; 徐瑞晶等, 2020)。無耳藤竹(Dinochloaorenuda)和響子竹(Bonialevigata)是海南島熱帶雨林的攀緣竹代表種,是熱帶低地雨林的重要組分。無耳藤竹喜陽、喜攀抵雨林冠層爭奪光資源,能較好適應高光環境,多分布于雨林林緣或林窗陽光充足處; 響子竹較為耐蔭,喜在林下攀爬,多分布在雨林林下弱光環境中。目前,有關2個竹種的研究較少,僅涉及分類學、地徑與節長分布和光合特性研究(Xia, 1996; 胡璇等, 2018; 徐瑞晶等, 2020)。本文系統研究2種攀緣竹的葉片解剖結構、氣體交換參數、單葉面積和葉片干物質含量等,可以揭示攀緣竹在熱帶雨林的生存與適應策略,為其資源保護和利用提供科學依據。
1 研究地概況
研究地點位于海南甘什嶺省級自然保護區(109°40′4″E,18°23′2″N,海拔202 m)。保護區地處三亞市與保亭縣南部交界處,屬低山丘陵地貌,坡度30°~50°,海拔50~681 m,土壤母質為花崗巖,巖層裸露率10%,土壤含砂量為20%左右。該區屬熱帶海洋季風氣候,年降水量1 200~1 800 mm,干濕季分明,雨季為6—11月,其降水占全年90%以上,旱季為12月—翌年5月。年均氣溫23.4 ℃,最低月平均氣溫19.2 ℃,最高月平均氣溫27.1 ℃(Wheat, 2020)。地帶性植被為熱帶低地雨林,喬木樹種有鐵凌(Hopeareticulata)、瓊南柿(Diospyroshowii)、華潤楠(Machiluschinensis)和青梅(Vaticamangachapoi)等,灌木有刺軸櫚(Licualaspinosa)、海南龍船花(Ixorahainanensis)、山血丹(Ardisialindleyana)等,草本有高稈珍珠茅(Scleriaterrestris)、烏毛蕨(Blechnumorientale)和華山姜(Alpiniaoblongifolia)等,層間植物有百足藤(Pothosrepens)、無耳藤竹和響子竹等(漆良華等, 2014)。
2 研究方法
2.1 試驗取樣 所調查的無耳藤竹平均地徑(2.3±0.2)cm,單節平均長(35.4±0.4)cm,分布密度為(180±51)叢·hm-2,攀爬高度可至18 m; 響子竹平均地徑(0.6±0.1)cm,單節平均長(25±0.6)cm,分布密度為(60±12)叢·hm-2,攀爬高度可至6 m。
在試驗前,每個竹種選5株長勢相近的植株進行掛牌,作為不同月份取樣測定的固定植株。分別于2019年6、9、12月和2020年3月選擇晴朗天氣的8:30—11:30和14:30—17:30期間,測定2種攀緣竹葉的光合氣體交換參數,每個竹種每次測量5株長勢相近的植株,每株選取枝條頂端第3~5片完全展開的3片健康成熟葉作為待測葉片。在葉片光合氣體交換參數測定結束后,在3片測定葉中采集1片葉,剪成1 cm×1 cm大小,分別置于70% FAA固定液中保存,帶回實驗室進行葉片石蠟切片和葉片表皮制片,用于觀察解剖結構。每個竹種采集植株上5個枝條的全部葉片,帶回實驗室測定葉片功能性狀,每次采樣5個植株。
2.2 測定方法 1) 葉片光合氣體交換參數測定 用紅藍光源模擬自然光,在無耳藤竹(1 100 μmol·m-2s-1)和響子竹(900 μmol·m-2s-1)的光飽和點附近設定光強,每片葉在飽和點光強下誘導15 min后讀數5次,測定其葉片凈光合速率(Pn)、氣孔導度(Cond)、胞間二氧化碳濃度(Ci)和蒸騰速率(Tr),并計算水分利用效率(WUE=凈光合速率/蒸騰速率)(張向峰等, 2012)。
2) 葉片解剖結構觀測 葉片石蠟切片制作參照李正理(1987)的方法,材料經軟化、脫水浸蠟、包埋、切片、番紅-固綠染色和中性樹膠封片等步驟; 而后在奧斯微PH50-3 M100光學顯微鏡下觀測和拍照,用ImageView圖像分析軟件測量各組織葉厚(LT)、上角質層厚度(USCT)、乳突厚度(MPT)、上表皮厚度(UET)、下表皮厚度(LET)、泡狀細胞數量(NBC)、泡狀細胞截面積(SABC)、單個梭形細胞截面積(SAFC)、主脈維管束截面積(SAFVB)、二級維管束直徑(DSVB)、二級維管束橫截面積(SASVB)、二級維管束間距(DBAVB)共12個指標。每個竹種在每次觀測均取30個視野統計指標值。
葉表皮切片制作用過氧化氫-冰醋酸離析法(唐光大等, 2010)。將葉片放入裝有1∶1的冰醋酸與30%過氧化氫混合液的消煮瓶,置于60 ℃水浴溫箱中27 h,待葉肉組織與上、下表皮開始分離時,把離析后的材料取出移入盛有蒸餾水的容器中,用毛筆分離上、下表皮,將洗凈后的葉表皮放入染色皿中,在1%番紅50%酒精溶液內染色5 min后用中性樹膠封片。切片在奧斯微PH50-3 M100光學顯微鏡下觀測和拍照,用ImageView圖像分析軟件測量各組織的氣孔長度(SL)、氣孔寬度(SW)、單個氣孔器面積(AS),并計算氣孔密度(SD=視野內氣孔個數/視野面積)和氣孔器面積百分比(AP=單個氣孔器面積×氣孔密度×100%)(李冬林等, 2019)。每個竹種在每次觀測均取30個視野統計指標值。
3) 單葉面積、葉片干物質含量和比葉面積測定 采用Epson perfection V19通過對葉片掃描測定單葉面積。將成熟葉剪去葉柄,浸泡在清水中12 h,用吸水紙吸去葉表水分,稱飽和鮮質量,然后105 ℃下殺青、80 ℃烘干至恒質量,稱取干質量。干物質含量(LDMC)=干質量(g)/飽和鮮質量(kg); 比葉面積(SLA)=葉面積(m2)/干質量(kg)(郭雯等, 2019)。
4) 葉片指標可塑性指數計算 葉片指標可塑性指數=(某指標的最大值-最小值)/最大值。可塑性指數范圍在0~1之間,數值越大表明可塑性越大,說明該指標對環境適應性越強(Valladaresetal.,2000)。
2.3 數據處理 采用SPSS 20.0對葉片光合氣體交換參數、解剖結構指標和形態性狀進行多重比較(SNK法),竹種間的指標差異顯著性分析使用獨立樣本t檢驗,使用Excel 2010和Sigma plot 12.5數據處理軟件對所測數據進行統計分析和繪圖。
3 結果與分析
3.1 葉片光合氣體交換參數月份變化 2種攀緣竹在不同月份的光合氣體交換參數結果(圖1)表明,無耳藤竹的凈光合速率、氣孔導度、胞間二氧化碳濃度和蒸騰速率在6月最高,12月最低; 水分利用效率則相反; 反映出無耳藤竹的光合作用能力在雨季(6、9月)高于旱季(12、3月)。響子竹凈光合速率、氣孔導度、胞間二氧化碳濃度和蒸騰速率在9月最高,3月和6月較低; 水分利用效率在3月最高,9月最低。對2個竹種,除12月的胞間二氧化碳濃度外,無耳藤竹的凈光合速率、氣孔導度、胞間二氧化碳濃度和蒸騰速率在各月均高于響子竹,2竹種各指標參數在6月和3月之間的差異顯著(P<0.05)。無耳藤竹的水分利用效率在6月和3月均顯著低于響子竹(P<0.05),在9月和12月高于響子竹,但差異不顯著(P>0.05)。以上結果反映了無耳藤竹的葉片光合能力強于響子竹,它們的水分利用效率在各月表現不同。
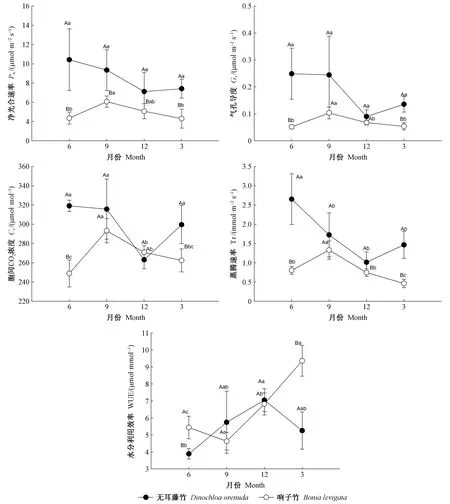
圖1 2種攀緣竹葉片的光合氣體交換參數月份動態Fig. 1 Monthly dynamics of photosynthetic gas exchange parameters in the leaves of two climbing bamboo species 同月份不同大寫字母表示2物種在0.05水平存在顯著差異,而同物種不同小寫字母表示月份間在0.05水平存在顯著差異(P<0.05)。 下同。 Different capital letters in the same month indicate the significant difference between species at 0.05,while different lowercase letters in the same species indicate the significant difference among months at 0.05(P<0.05). The same below.
3.2 葉片解剖結構月份變化 2種攀緣竹的葉片由表皮、葉肉和葉脈3部分組成(圖2),它們的解剖結構組成基本相似。竹葉表皮分上表皮和下表皮,均含有1層細胞,上表皮細胞較厚,下表皮細胞較薄; 表皮細胞由長、短2種細胞組成,上表皮有少量氣孔分布,下表皮氣孔分布較多,氣孔的每個保衛細胞外側各有一個副衛細胞,氣孔器分布在脈間區域并與葉脈平行; 在上表皮的2個維管束之間分布有2~6個排列在一起、橫切面呈扇形的泡狀細胞,表皮上層有少量角質層; 下表皮分布有乳突、氣孔器和粗毛,無耳藤竹著生粗毛較多,響子竹較少有或沒有。2種攀緣竹的葉肉細胞沒有柵欄和海綿組織分化,為等面葉,葉肉分布有大量自上而下的單向指狀內褶疊的臂細胞,在維管束兩側有2個透明的梭形細胞,梭形細胞隨維管束增大而變小。2種攀緣竹的主脈由多個維管束組成,二級脈只有1個維管束,維管束鞘細胞1~3層,均沒有形成花環結構,為C3植物。

圖2 2種攀緣竹葉片解剖特征Fig. 2 Anatomical characteristics in the leaves of two climbing bamboo species a.無耳藤竹葉片橫切面; b.響子竹葉片橫切面; c.無耳藤竹上表皮; d.響子竹上表皮; e.無耳藤竹下表皮; f.響子竹下表皮。UC: 上角質層; UE: 上表皮細胞; LE: 下表皮細胞; EH: 表皮毛; MP: 乳突; BC: 泡狀細胞; BS: 維管束鞘; V: 導管; Ph: 韌皮部; Xy: 木質部; St: 氣孔; LC: 長細胞; SC: 短細胞; FC:梭形細胞。a. Leaf transverse section of D. orenuda; b. Leaf transverse section of B. levigata; c. Upper epidermal thickness of D. orenuda; d.Upper epidermal thickness of B. levigata; e.Lower epidermal thickness of D. orenuda; f. Lower epidermal thickness of B.levigata. UC: Upper stratum corneum; UE: Upper epiderma; LE: Lower epiderma; EH: Epidermal hair; MP: Mastoid process; BC: Bulliform cell; BS: Vascular bundle sheath; V: Vessels; Ph: Phloem; Xy: Xylem; St:Stomata; LC: Long cell; SC: Short cell; FC: Fusoid cell.
2種攀緣竹的葉厚、泡狀細胞數量、泡狀細胞截面積、二級維管束直徑、二級維管束截面積、二級維管束間距和氣孔器面積占比均為雨季月份高于旱季月份(表1); 此外,單個梭形細胞截面積、氣孔長度、單個氣孔器面積在6月或9月達到最高,響子竹的上角質層厚度旱季高于雨季。2竹種的乳突厚度、下表皮厚度、氣孔寬度和氣孔密度最大值在各月表現不同。在各月的17個解剖結構指標中,2竹種的二級維管束直徑、二級維管束截面積、氣孔寬度和單個氣孔器面積4個指標和無耳藤竹的泡狀細胞數量、氣孔長度以及響子竹的上角質層厚度、氣孔器面積占比的最大值與最小值的差異均達顯著水平(P<0.05),說明這些指標對月份變化較敏感。以上結果表明攀緣竹大部分解剖結構指標值在雨季較大,旱季較小; 二級維管束直徑、二級維管束截面積、氣孔寬度和單個氣孔器面積4個指標較為敏感,其余12個指標的月份變化較小,較為穩定。
從2個竹種對比來看(表1),無耳藤竹的葉厚、氣孔寬度、單個氣孔器面積、氣孔器面積占比在各月均高于響子竹,乳突厚度、上表皮厚度、二級維管束間距、氣孔密度均低于響子竹。此外,無耳藤竹的上角質層厚度、泡狀細胞數量、泡狀細胞截面積、氣孔長度在雨季各月高于響子竹,而下表皮厚度在雨季各月低于響子竹。2種攀緣竹的葉厚、乳突厚度、氣孔寬度、單個氣孔器面積、氣孔密度和氣孔器面積占比差異顯著(P<0.05),說明葉厚、乳突和氣孔是2個竹種葉片解剖結構差異的關鍵指標。
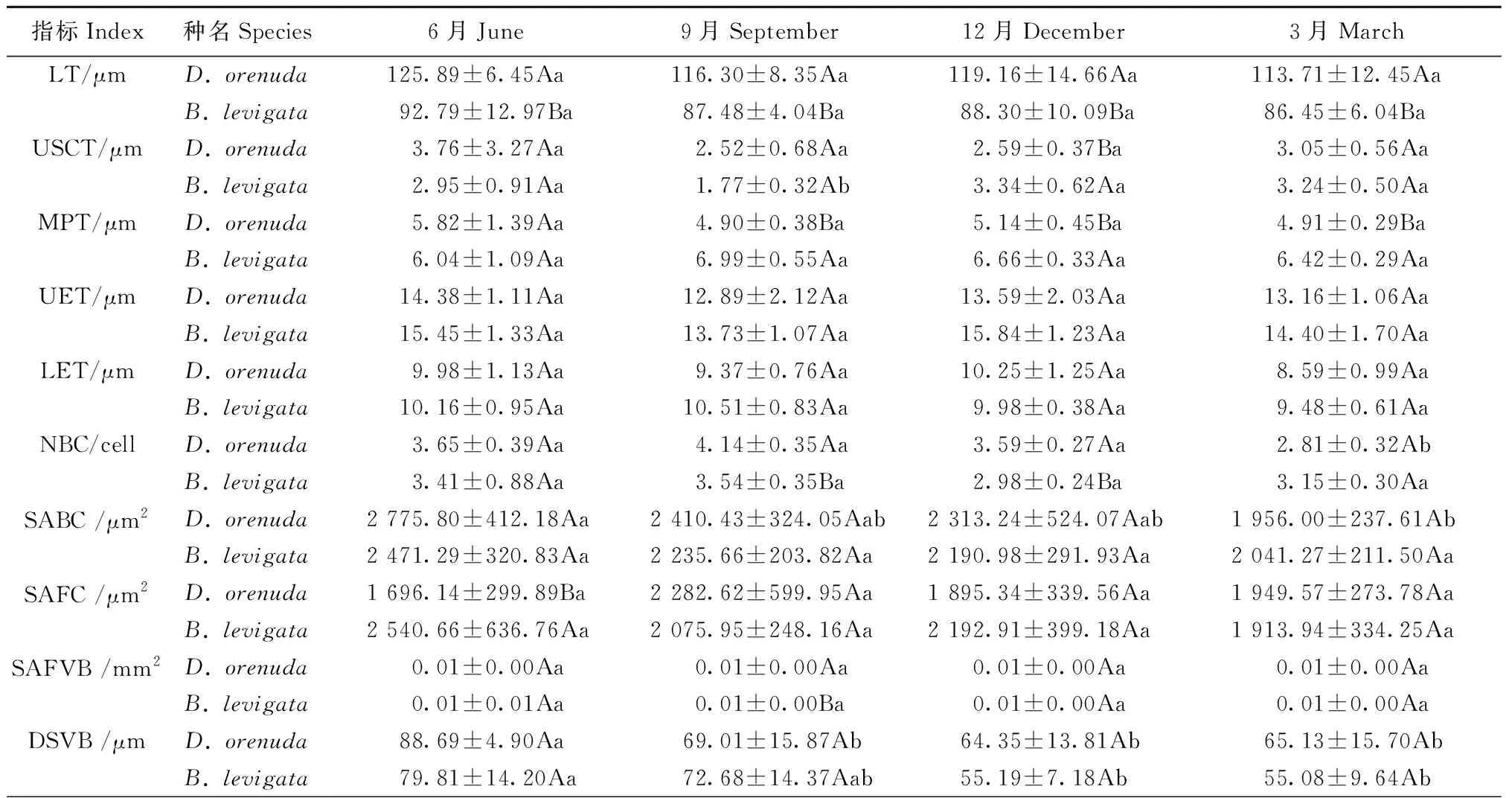
表1 2種攀緣竹葉片解剖結構月份動態①Tab.1 Monthly dynamics of anatomical characteristics in the leaves of two climbing bamboo species
3.3 葉片形態性狀的月份變化 各月的葉片形態性狀測定結果存在差異(圖3)。無耳藤竹的單葉面積在9月最高,6月最低; 響子竹則在6月最高,3月最低,最高和最低值之間的差異顯著(P<0.05)。葉片干物質含量在6月最低,最高值在2個竹種表現不同,2個竹種的最高值和最低值之間差異顯著(P<0.05)。2個竹種的比葉面積在6月和3月較高,9月和12月較低,最高和最低值之間差異顯著(P<0.05)。以上結果表明攀緣竹的葉片形態性狀存在明顯的季節差異。
在2個竹種之間,響子竹的單葉面積和干物質含量(6月除外)高于無耳藤竹(圖3)。無耳藤竹的葉子干物質含量在雨季高于響子竹,在旱季則相反。除6月的單葉面積、12月和3月的葉片干物質含量外,2個竹種的各形態性狀指標差異不顯著(P>0.05)。以上結果表明2個竹種的葉片形態性狀指標存在差異,但差異不明顯。
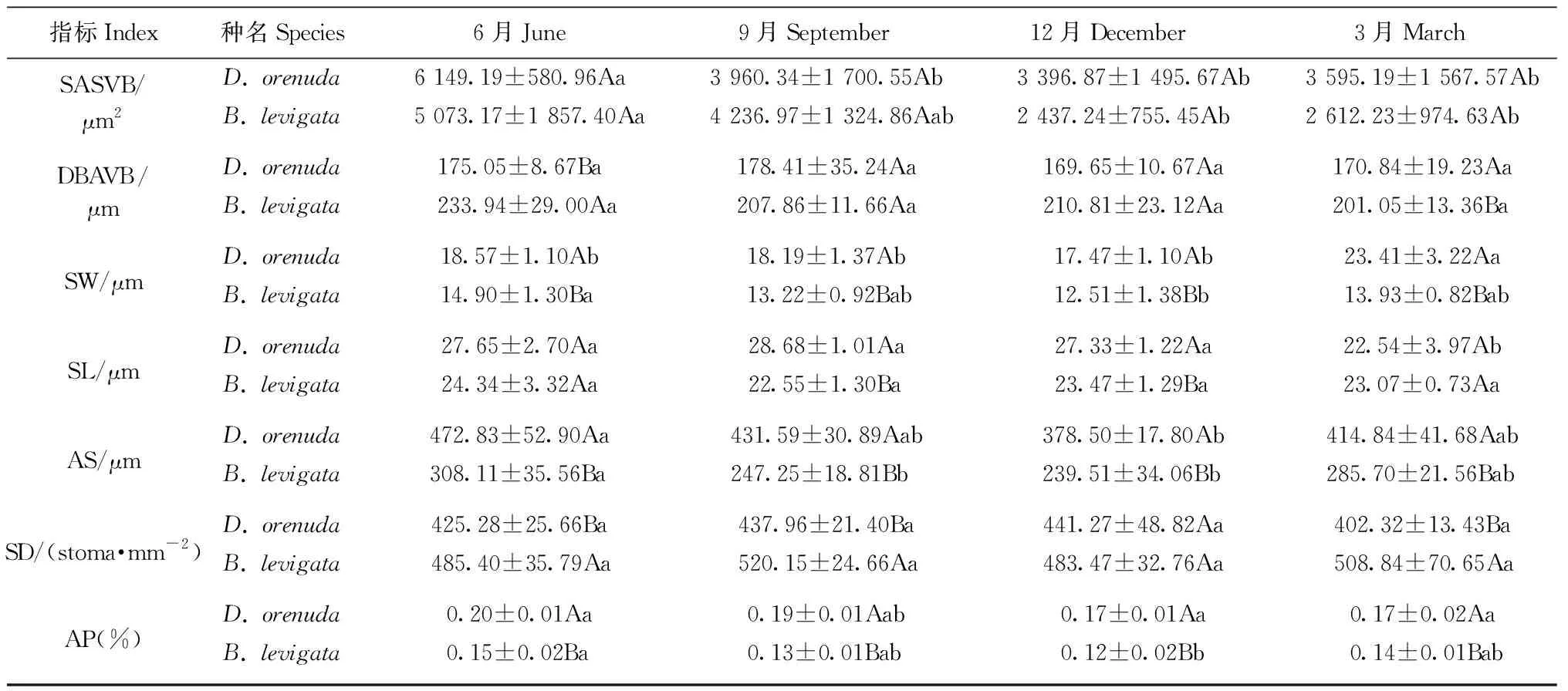
續表1 Continued
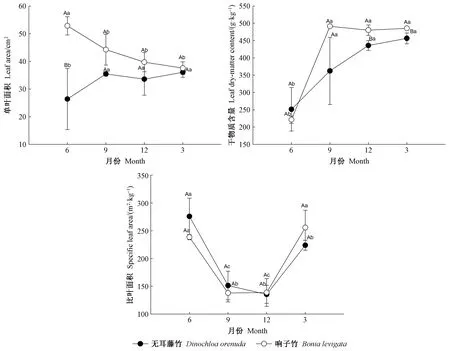
圖3 2種攀緣竹葉片形態性狀月份變化Fig. 3 Monthly dynamics of morphological traits in the leaves of two climbing bamboo species
3.4 葉片指標的可塑性 攀緣竹各月的葉片指標可塑性對比表明(表2),無耳藤竹的各指標平均可塑性在9月最高(0.34),12月最低(0.27),響子竹則是6月最高(0.28),9月最低(0.22)。無耳藤竹的各月平均可塑性指標均高于響子竹,全年的各指標平均可塑性(0.53)也高于響子竹(0.49),反映了無耳藤竹的環境適應能力高于響子竹。2竹種的氣孔導度、蒸騰速率、上角質層厚度、主脈截面積和二級維管束截面積的可塑性較大,表明這些指標具有較大環境適應性。胞間二氧化碳濃度、葉厚、上表皮厚度、下表皮厚度、單個氣孔器面積、氣孔密度的可塑性均較低,表明這些指標具有較高的穩定性。
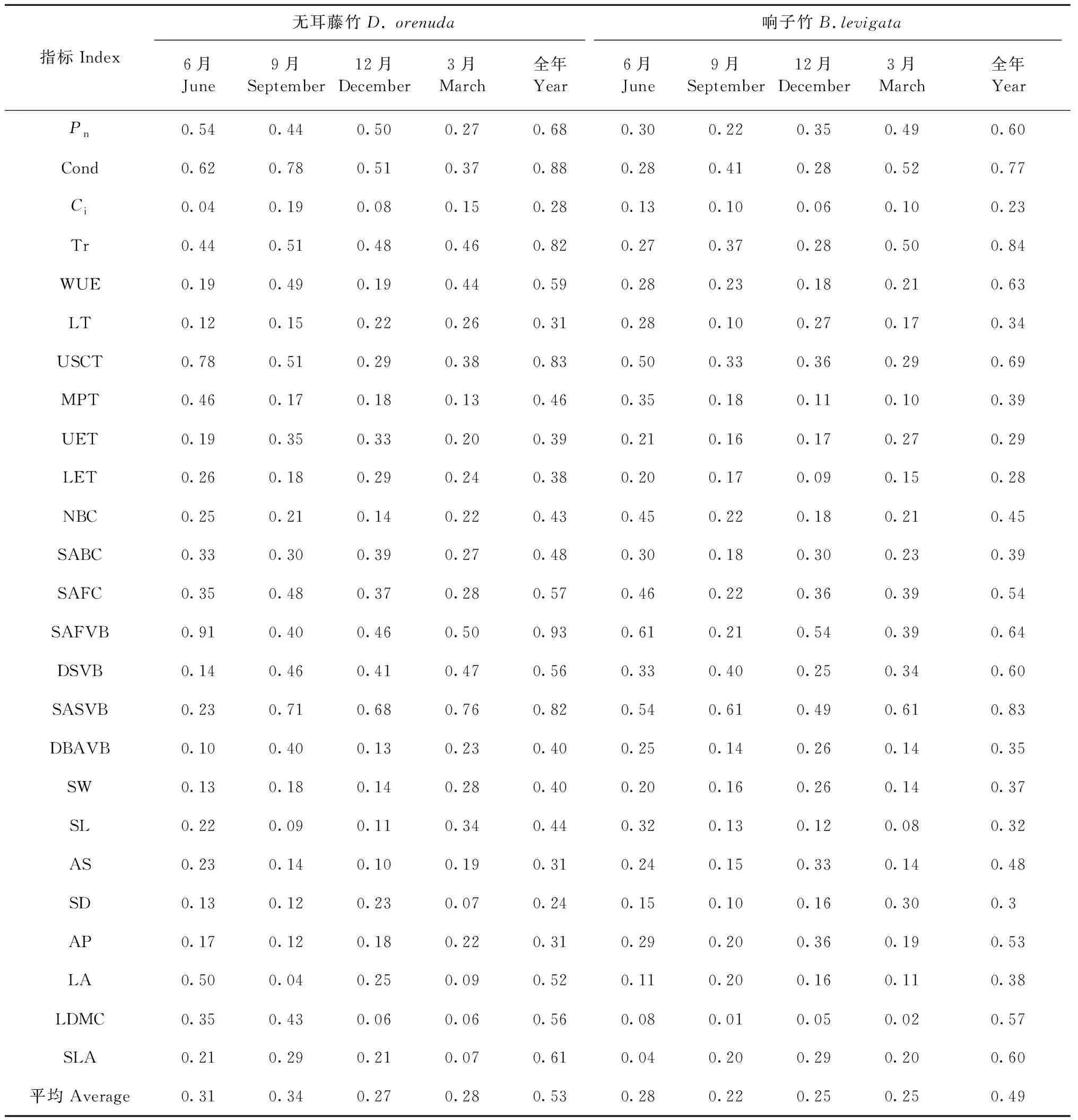
表2 2種攀緣竹葉片可塑性指數①Tab.2 Plasticity index in the leaves of two climbing bamboo species
3 討論
3.1 2種攀援竹葉片功能性狀的月份動態 2種攀援竹葉片的凈光合速率、氣孔導度、胞間二氧化碳濃度和蒸騰速率在雨季高于旱季,與云南季雨林的網脈核果木(Drypetesperreticulata)和福建紅樹林的秋茄(Kandeliaobovata)等樹種的葉片光合特性研究結果(栗忠飛等, 2012; 黃麗, 2013)一致; 水分利用效率和葉片光合能力表現與前期開展的攀緣竹光合特性研究結果(徐瑞晶等, 2020)一致,說明2竹種的光合作用年際間較穩定,這是2種攀緣竹長期適應熱帶低地雨林生境的結果。
攀緣竹的葉厚、乳突厚度、氣孔寬度、氣孔密度等解剖結構指標值在雨季大于旱季,反映了攀緣竹葉片在雨季通過增大以上組織結構以適應雨季快速生長。同時,二級維管束直徑、二級維管束截面積和單個氣孔器面積在雨季顯著高于旱季(P<0.05),可有效增強竹葉的物質運輸和光合氣體交換能力,促進攀緣竹在雨季快速生長。
葉片性狀的季節變化可反映植物的環境適應能力,不同指標間的表現是植物生長策略權衡的結果(薛立等, 2012)。2竹種的比葉面積在6月和3月較高,9月和12月較低,主要原因是竹葉的雨季光合作用增強,葉片積累了大量的有機物,其干質量增加,表現出具有較低的比葉面積; 而進入旱季后,葉片光合能力下降,降低了積累有機物的能力,因此比葉面積處于較高水平。2竹種葉片的干物質含量的月份變化趨勢與其光合作用有機物積累的季節變化相關。2竹種的單葉面積變化特征與它們的葉片生長發育規律有關,其新生葉片的單葉面積在前期較小,在發育到一定階段后基本保持不變。
3.2 2種攀緣竹的葉片適應策略 植物葉片可塑性越大,其種群分布越廣泛,生態適應性越強(蔣琴等, 2018)。作者野外調查中發現,無耳藤竹在熱帶雨林分布更廣泛,響子竹種群分布范圍較小,這與藤竹葉片可塑性有關。海南島甘什嶺熱帶低地雨林區,雨季降雨量占全年的90%以上,雨季和旱季更替是該區重要的環境變化。本研究中,2竹種的蒸騰速率、角質層厚度、主脈截面積和二級維管束截面積的可塑性均較高,它們是水分利用的重要結構指標,通過這些指標的調節利于適應雨季旱季更替引起的環境變化。無耳藤竹的乳突厚度、上表皮厚度、二級維管束間距、氣孔密度均低于響子竹。此外,無耳藤竹的上角質層厚度、泡狀細胞數量、泡狀細胞截面積、氣孔長度在雨季高于響子竹,而下表皮厚度在雨季低于響子竹。2種攀緣竹的葉厚、乳突厚度、氣孔寬度、單個氣孔器面積、氣孔密度和氣孔器面積占比差異顯著(P<0.05),葉厚、乳突和氣孔是反映2個竹種葉片解剖結構差異的關鍵指標。
植物葉性狀與植物的生長對策和植物的資源利用能力密切相關(Reichetal., 2010)。本研究表明,無耳藤竹的葉片凈光合速率、葉厚、氣孔寬度、單個氣孔器面積和氣孔器面積占比在各月均顯著高于響子竹,但二級維管束間距低于響子竹。無耳藤竹有極強的攀爬能力,其枝條可攀抵熱帶雨林的林冠層,葉片更適應強光環境,這與馬光天等(2018)研究認為強光環境下,葉片通過增大葉厚減小單葉面積以減少光輻射量,避免強光對葉片的傷害; 弱光環境下,植物傾向于減小葉厚增大單葉面積,以增強葉片細胞對弱光的捕獲能力的研究結果一致。林冠層葉片偏向旱生結構,在結構上偏向于減少水分散失,在功能上提高水分利用效率,從而適應水分虧缺(何春霞等, 2013)。葉厚是評價植物干旱適應性的重要指標,葉片越厚則適應干旱能力越強(馬建靜等, 2012),較厚葉片可幫助無耳藤竹緩解林冠層水分虧缺和旱季雨量減少造成的干旱脅迫。氣孔寬度、單個氣孔器面積和氣孔器面積占比是控制葉片光合作用氣體交換的關鍵結構指標,它們的較高數值利于提高葉片同化二氧化碳的效率。無耳藤竹葉片的二級維管束間距較小,說明其葉片單位面積內維管束密度較高,輸送養分水分的功能更強; 以上葉片特征有助于無耳藤竹提高其光合和攀爬能力、適應冠層水分虧缺和旱季脅迫。與無耳藤竹相反,響子竹的凈光合速率、氣孔寬度、單個氣孔器面積、氣孔器面積占比較低,很難抵達林冠層,決定了其較為耐蔭的習性; 乳突可減少氣孔水分蒸發,利于響子竹適應逆境脅迫(蔣琴等, 2018),響子竹憑借較厚的乳突和上表皮、數量多而面積小的氣孔來減少水分蒸發,可更高效地利用水分(王媛媛等, 2018),以適應旱季脅迫。
3.3 2種攀緣竹資源的保護和利用 無耳藤竹和響子竹均自然分布于熱帶雨林中,其攀援特性決定了必須依靠其他植物才能更好生長。尤其是無耳藤竹,需依靠其他樹木的支撐才能攀至林冠層,滿足其喜陽特性的生境要求; 響子竹較耐蔭,不需抵達林冠層,但其竿莖纖細柔軟,必須依附于其他樹木才能更好生存。因此,為保護攀緣竹,在加強攀緣竹種群和個體保護的同時,還要重視對其群落和支撐樹木的保護。在攀緣竹利用上,無耳藤竹和響子竹植株均呈攀援或懸垂狀,明顯區別于直立竹,具有較高觀賞價值。響子竹竹竿纖細,葉片纖長柔美,竹態曼妙,可作為廊道、花架和墻體等垂直綠化特色竹種; 無耳藤竹的竹莖可達3 cm,柔韌性較好,可作為編織材料,其竹筍可食用,頗受當地農民喜愛,亦可作特色筍用竹。在攀緣竹資源培育上,2種攀緣竹屬特殊的叢生竹,可采用分蔸移植。無耳藤竹較喜陽,苗期管理時要保證一定光照強度,而響子竹較耐蔭,需營造遮蔭環境。在野外狀態下,2種攀緣竹的光飽和點存在一定的月份差異,光照條件營造需根據月份適當調整。無耳藤竹和響子竹的各月平均光飽和點分別為(1 032±217)和(877±183) μmol·m-2s-1(徐瑞晶等, 2020),可作為其育苗光照條件的參考,但苗期的最適光照強度還需進一步研究。無耳藤竹和響子竹的光合能力均在雨季月份(6月和9月)最高,培育竹苗時這段時間內要加強水肥供給,以保證竹苗更好生長; 在旱季月份(12月和3月),要保證足夠水分供給,減少干旱對生長的脅迫。
4 結論
2種攀緣竹葉片的結構、光合特性和形態性狀特征決定了它們不同的生態習性和生存策略。無耳藤竹和響子竹在雨季通過增大與物質運輸和光合氣體交換相關的結構,來實現快速生長,同時為即將到來的旱季脅迫做好物質儲備。無耳藤竹具有較高的葉片指標可塑性,表明其有更強的旱季適應性,能憑借較大的光合能力、葉厚、氣孔和較低的單葉面積、比葉面積、二級維管束間距等指標,來適應林冠層的強光環境、冠層水分虧缺和旱季脅迫。響子竹則通過其葉子的較高的單葉面積、比葉面積、上表皮厚度、乳突厚度和較薄的葉片及小而密的氣孔,適應旱季脅迫和林下弱光環境。
