宏基因組測序分析野生林麝瘤胃、小腸和大腸微生物組成和抗生素抗性基因
2021-02-07 01:02:38曹新芳鄭雪莉王洪永蔣本模卜書海
動物營養學報 2021年1期
何 森 曹新芳 鄭雪莉 王 豆 王洪永 蔣本模 卜書海*
(1.西北農林科技大學生命科學學院,楊凌712100;2.西北農林科技大學林學院,楊凌712100;3.鳳縣逢春濟民可信科技養殖有限公司,鳳縣721000)
林麝(Moschusberezovskii)屬于麝科(Moschidae)麝屬(Moschus),目前已被列為我國一級保護動物,其所產麝香是一種藥用價值極高的動物產品。林麝在野生狀態下屬于泛食、嫩食動物,即使在食物匱乏期仍會回避高纖維植物[1]。對野生林麝消化道微生物進行研究有助于我們了解自然狀態下林麝消化道微生物組成,對野生林麝保護及圈養林麝飼養具有借鑒和參考價值。
近年來對圈養反芻動物瘤胃微生物的研究,摸清了牛、羊等動物瘤胃中微生物群落組成,發現其對食物發酵與初步消化具有重要意義,消化道微生物活動會影響動物產品質量[2-3]。林麝屬于反芻動物,這使它們容易受到消化道微生物群落活動的影響[4]。在圈養條件下,由環境問題和飼喂不當引起的菌群失衡已經成為影響麝健康的重要因素[5]。目前因采樣受限,對林麝消化道微生物的研究進展緩慢[6],僅有的林麝糞便研究表明,環境、食物、健康狀態等因素都會影響林麝消化道微生物組成,其中與營養分解相關的核心菌群(擬桿菌門和厚壁菌門細菌)豐度變化不大,而非核心菌群的種類和豐度變化明顯[6-8],這些微生物在免疫反應和營養消化方面發揮著重要作用[9]。
目前林麝消化道微生物研究樣本以糞便為主,常用的方法是16S rRNA高通量測序,這種方法的缺點是測序深度有限,而宏基因組測序不僅能獲得全面的微生物類群信息,還能提供基因功能層面的預測,近年來被廣泛用于微生物群落研究[10]。糞便采樣具有集合了貫穿胃腸道分散微生物群落的優勢,但其與大腸微生物相似度較高,不能完全反映除大腸外其他消化道部位微生物群落結構的真實情況[11]。為全面了解野生林麝消化道微生物結構及其抗生素抗性基因(ARGs)種類,對其消化道分部位取樣并進行宏基因組測序是十分必要的。
鑒于此,本試驗采用宏基因組測序的方法,對1只珍貴的野生林麝消化道不同區段微生物群落結構和ARGs進行研究,旨在更全面地分析野生林麝消化道不同部位微生物組成及所攜帶ARGs的特點和差異,為人工養麝防治胃腸道疾病提供深入的數據支持與參考。
1 材料與方法
1.1 試驗材料
試驗樣本取自陜西省寶雞市扶風縣一柿園內遭家犬追咬死亡的野生雄性林麝,年齡1.5歲,經解剖后觀察,樣本發育良好,器官均無病變,表明樣本在死亡前健康狀況良好。采樣時間為初夏(5月中旬),樣本死亡后及時取瘤胃、小腸和大腸內容物加50%甘油于-40 ℃保存。
1.2 試驗方法
1.2.1 DNA提取與宏基因組建庫
采用十二烷基苯磺酸鈉(SDS)法對樣本基因組DNA提取[12],采用瓊脂糖凝膠電泳(AGE)和Qubit 2.0熒光計對DNA質量進行檢測,然后用Covaris超聲波破碎儀將樣品DNA隨機打斷成長度為350 bp的片段,經末端修復、加A尾、加測序接頭、純化、PCR擴增等步驟完成文庫制備。DNA文庫構建完成后,使用Qubit 2.0熒光計進行初步定量,稀釋文庫濃度至2 ng/μL,Agilent 2100分析儀對文庫的插入片段進行檢測。符合預期后使用定量PCR(q-PCR)法對DNA文庫有效濃度進行準確定量,不同文庫按照有效濃度及目標下機數據量需求合并,然后用Illumina PE150測序儀測序,最終得到原始數據。
1.2.2 生物信息學分析
對下機的原始數據進行質控,過濾掉低質量片段和宿主片段后得到有效數據,然后對有效數據進行宏基因組裝,為保證數據可靠,將各樣品未被利用的片段放在一起進行混合組裝。從單樣品和混合組裝樣品中不含N端的片段出發,采用MetaGeneMark軟件進行基因預測,將各單樣品和混合組裝樣品預測產生的基因放在一起進行去冗余,去冗余后的片段用于構建基因目錄,綜合各樣品有效數據獲得基因目錄在各樣品中的豐度信息。將基因目錄與MicroNR庫進行比對,獲得每個基因的物種注釋信息,然后與KEGG數據庫、Card數據庫進行比對得到相關基因注釋結果。
2 結果與分析
2.1 宏基因組測序
經質控過濾,小腸、瘤胃和大腸中分別獲得了11 354.99、13 246.22和14 315.52 Mbp有效數據。有效數據中測序錯誤率小于1%(Q20)和1‰(Q30)的堿基數目比例分別達到97%和92%以上,表明測序數據可靠性較高(表1)。基因預測共得到11 461 517 條開放閱讀框(ORFs)用于后續物種注釋和基因功能分析。
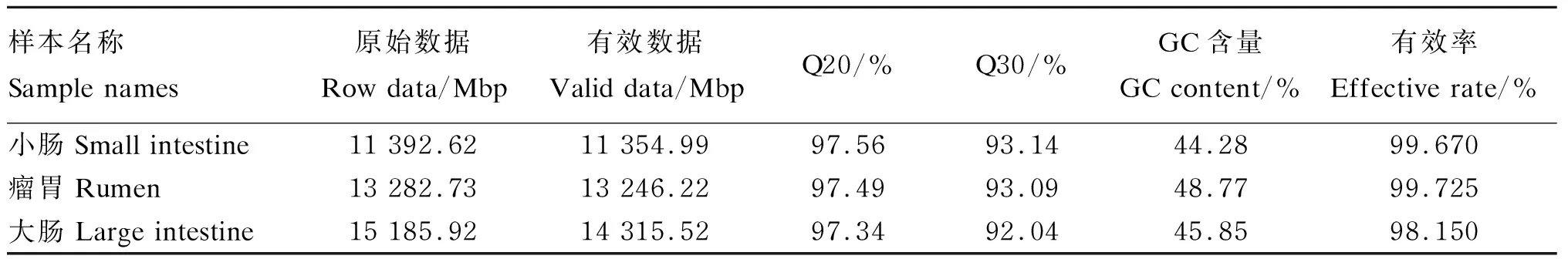
表1 數據預處理統計
2.2 野生林麝瘤胃、小腸和大腸微生物組成
2.2.1 野生林麝瘤胃、小腸和大腸細菌組成
消化道3個區段中優勢菌門組成存在一定差異,瘤胃中優勢菌門為厚壁菌門(51%)、擬桿菌門(23%)和互養菌門(12%);小腸中優勢菌門為變形菌門(43%)、放線菌門(23%)和厚壁菌門(17%);大腸中優勢菌門為厚壁菌門(73%)和擬桿菌門(22%),兩者比值明顯高于瘤胃(圖1-A)。屬水平上瘤胃和小腸細菌組成差異十分明顯,3個區段之間沒有共享的優勢菌屬(圖1-B)。瘤胃中主要菌屬是與碳水化合物、蛋白質分解相關的厭氧菌屬:普雷沃氏菌屬(16%)、月形單胞菌屬(8%)、依賴桿菌屬(6%)、擬桿菌屬(3%);而小腸中多為致病菌屬和條件致病菌屬:分枝桿菌屬(23%)、鏈球菌屬(15%)、埃希氏菌屬(14%)、無形體屬(12%);大腸中多為與營養物質分解相關的菌屬:梭菌屬(10%)、擬桿菌屬(8%)、另枝菌屬(8%)。
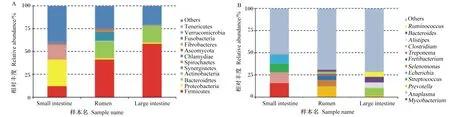
Small intestine:小腸;Rumen:瘤胃;Large intestine:大腸;Firmicutes:厚壁菌門;Proteobacteria:變形菌門;Bacteroidrtes:擬桿菌門;Actinobacteria:放線菌門;Synergistetes:互養菌門;Spirochaetes:螺旋體門;Chlamydiae:衣原體門;Ascomycota:子囊菌門;Tenericutes:柔膜菌門;Fibrobacteres:纖維桿菌門;Fusobacteria:梭桿菌門;Verrucomicrobia:疣微菌門;Mycobacterium:分枝桿菌屬;Anaplasma:無形體屬;Prevotella:普雷沃氏菌屬;Streptococcus:鏈球菌屬;Echerichia:埃希氏菌屬;Selenomonas:月形單胞菌屬;Fretibacterium:依賴桿菌屬;Treponema:密螺旋體屬;Alistipes:另枝菌屬;Bacteroides:擬桿菌屬;Clostridium:梭菌屬;Ruminococcus:瘤胃球菌屬;Others:其他。
2.2.2 野生林麝瘤胃、小腸和大腸古菌、真菌和病毒組成
消化道3個區段主要古菌為廣古菌門和奇古菌門,大腸中有少部分屬于泉古菌門(1%)和初古菌門(1%)(圖2-A)。在屬水平上,瘤胃和大腸優勢古菌屬相似,相對豐度最大的2個古菌屬均為甲烷八疊球菌屬、甲烷短桿菌屬,小腸中的古菌都屬于亞硝化侏儒菌屬(圖2-B)。
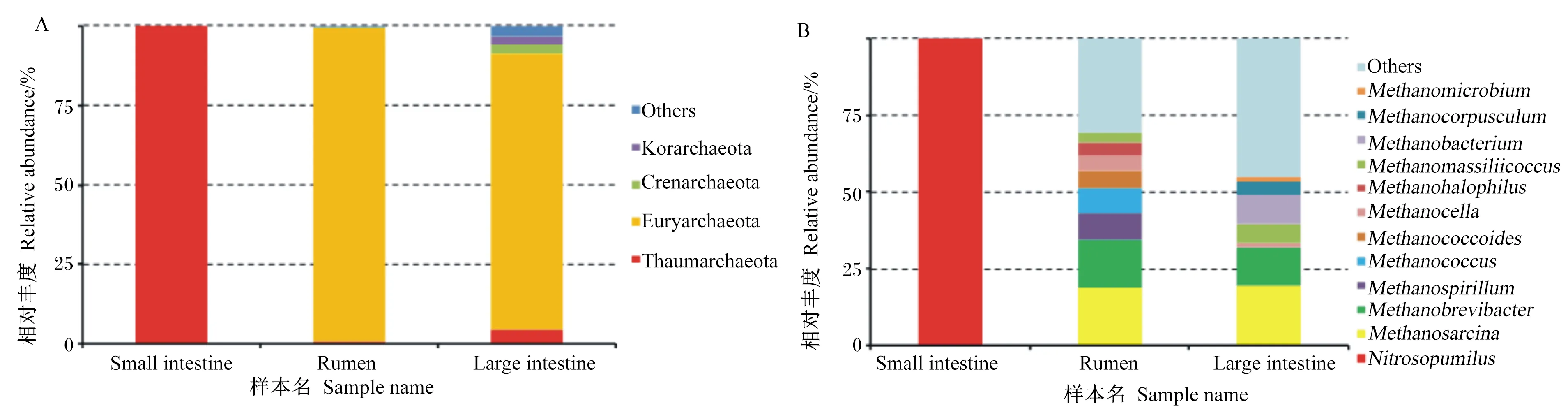
Small intestine:小腸;Rumen:瘤胃;Large intestine:大腸;Euryarchaeota:廣古菌門;Thaumarchaeota:奇古菌門;Crenarchaeota:泉古菌門;Korarchaeota:初古菌門;Methanosarcina:甲烷八疊球菌屬;Methanobrevibacter:甲烷短桿菌屬;Methanospirillum:甲烷螺菌屬;Methanococcus:產甲烷球菌屬;Nitrosopumilus:亞硝化侏儒菌屬;Methanococcoides :擬甲烷球菌屬;Methanocella:甲烷胞菌屬;Methanohalophilus:甲烷嗜鹽菌屬;Methanobacterium :甲烷桿菌屬;Methanocorpusculum :甲烷粒菌屬;Methanomicrobium:甲烷微菌屬;Others:其他。
瘤胃、小腸和大腸中共有的優勢真菌屬于擔子菌門、子囊菌門(圖3-A)。在屬水平上三者共有的優勢真菌為曲霉屬和根內球囊霉屬,瘤胃中優勢真菌還有Phaeomoniella(12%),小腸還有Saitoella(6%),大腸中還有假絲酵母菌屬(5%)、柄孢殼菌屬(3%)(圖3-B)。

Small intestine:小腸;Rumen:瘤胃;Large intestine:大腸;Basidiomycota:擔子菌門;Ascomycota:子囊菌門;Mucoromycota:毛霉亞門;Microsporidia:微孢子門;Chytridiomycota:壺菌門;Neocallimastigomycota:新麗鞭毛菌門;Cryptomycota:隱真菌門;Blastocladiomycota:芽枝霉門;Zoopagomycota:捕蟲霉門;Glomeromycota:球囊菌門;Aspergillus:曲霉屬;Rhizophagus:根內球囊霉屬;Saitoella:齊藤酵母屬;Phanerochaete:平革菌屬;Clavispora:棒孢酵母菌屬;Tilletia:腥黑粉菌屬;Puccinia:柄銹菌屬;Pseudogymnoascus:假裸囊菌屬;Candida:假絲酵母菌屬;Podospora:柄孢殼菌屬;Others:其他。
瘤胃中主要病毒種類是弧菌噬菌體(22%)、地中海噬菌體(22%)和芽孢桿菌噬菌體(9%);小腸中相對豐度最高的是副痘病毒屬(80%),其次是丙型反錄病毒屬(5%)、乙型反錄病毒屬(3%);大腸中優勢病毒種類為無害的地中海噬菌體(10%)、肌尾噬菌體(7%)和長尾噬菌體(6%)和(圖4)。
2.3 野生林麝瘤胃、小腸和大腸微生物基因功能注釋與差異分析
2.3.1 KEGG數據庫注釋差異分析
去冗余后的ORFs經過KEGG數據庫注釋,結果如圖5所示。消化道中微生物在數據庫中注釋數量最多的通路是新陳代謝。對注釋數據進行富集差異分析發現,瘤胃、小腸、大腸中微生物在不飽和脂肪酸生物合成、靈桿菌素生物合成以及新霉素、慶大霉素、卡那霉素生物合成這些方面差異顯著(P<0.05),還有一些通路上富集的差異基因數量很多,如淀粉和蔗糖代謝、其他抗生素生物合成、氨基糖和核苷酸糖代謝等。
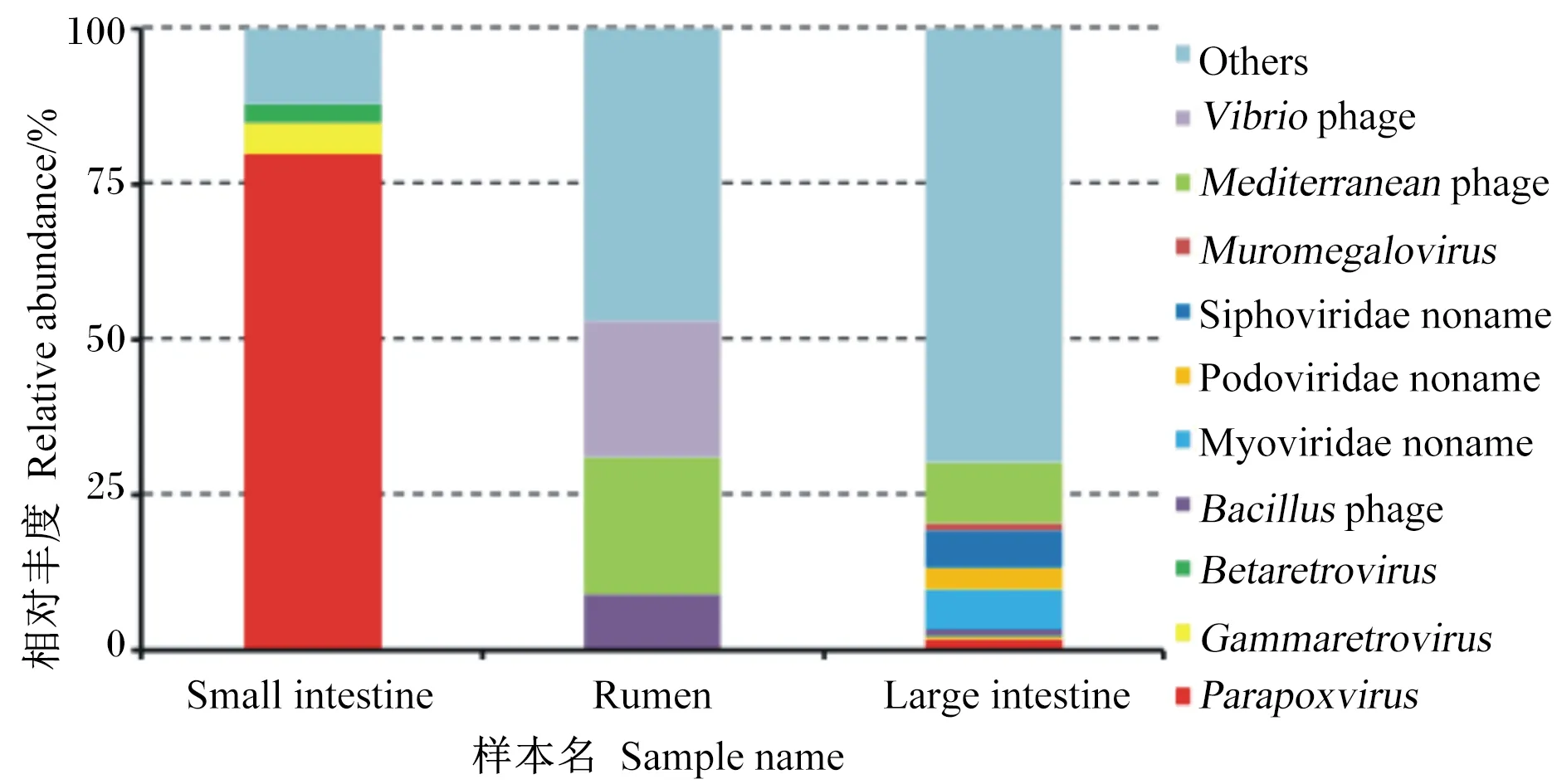
Small intestine:小腸;Rumen:瘤胃;Large intestine:大腸;Vibrio phage:弧菌噬菌體;Mediterranean phage:地中海噬菌體;Muromegalovirus:鼠巨細胞病毒屬;Siphoviridae noname:長尾噬菌體科未命名;Podoviridae noname:短尾噬菌體科未命名;Myoviridae noname:肌尾噬菌體科未命名;Bacillus phage:芽孢桿菌噬菌體;Betaretrovirus:乙型反轉錄病毒屬;Gammaretrovirus:丙型反轉錄病毒屬;Parapoxvirus:副痘病毒屬;Others:其他。
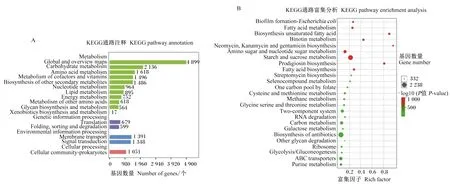
圖A展示了注釋在不同通路中的基因的數量;圖B展示了消化道不同區段差異基因的富集分析,點的大小代表差異基因個數,富集因子是當前通路差異基因數與注釋在這個通路中所有基因數的比值,顏色代表-log10(P值)的大小,值越大代表差異越顯著。The figure A shows the number of genes annotated in different pathways; the figure B shows the enrichment analysis of genes at different digestive tract segments. The size of the dots represents the number of differentially annotated genes. The rich factor is the ratio of the number of differentially annotated genes in the current pathway to the number of all genes in the pathway, and the color represents P-value, the larger value means the more significant the difference.
2.3.2 ARGs注釋差異分析
消化道細菌的ARGs對野生林麝的耐藥性貢獻較大,細菌ARGs注釋結果顯示,野生林麝瘤胃、小腸和大腸ARGs種類存在明顯差異(圖6)。絕對豐度最大的3種ARGs都來自大腸細菌,分別是macB、sav1866和bcrA。大腸中絕對豐度最高的ARGs是macB,瘤胃中絕對豐度最高的ARGs是Aminocoumarin_resistant_alaS,小腸中絕對豐度最高的ARGs是adeG。小腸微生物對adeG、TaeA絕對豐度的貢獻較高,分別達到50%和30%。
瘤胃中細菌ARGs主要來源于厚壁菌門(44%)和擬桿菌門(26%);小腸中細菌ARGs主要來源于變形菌門(93%),厚壁菌門和擬桿菌門中ARO所占比例較少(共3%);大腸中細菌ARGs主要來源于厚壁菌門(58%)和擬桿菌門(18%)。(圖7)
3 討 論
本研究對1只野生林麝的瘤胃、小腸和大腸內容物進行了宏基因組測序,研究野生林麝腸道微生物組成。雖然樣本量少,但是關鍵在于該野生林麝在自然界中自由采食,成長過程中未接觸人工抗生素,研究該林麝腸道微生物組成和ARGs種類,能為野生林麝保護和圈養林麝養殖提供重要參考依據。
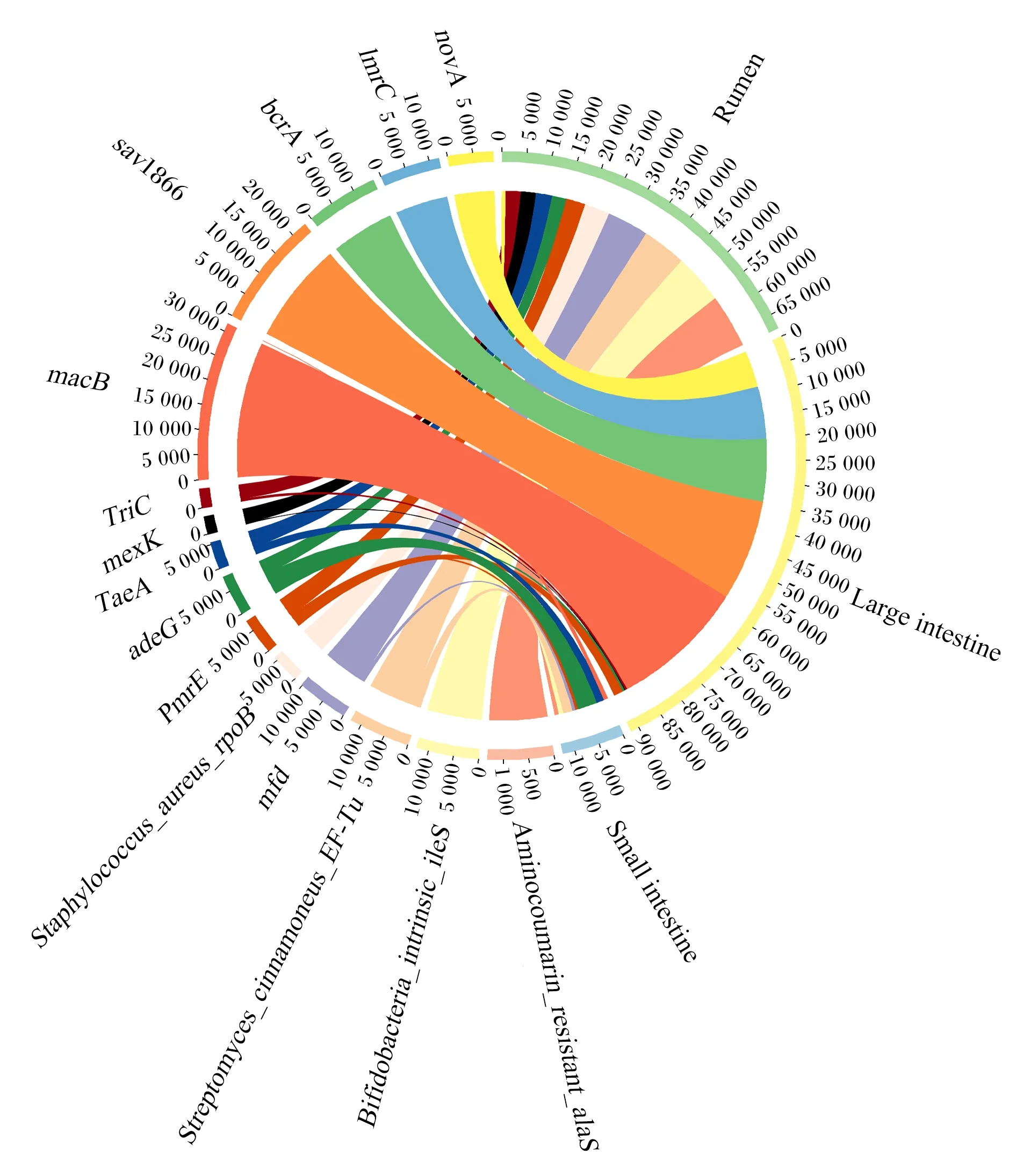
Small intestine:小腸;Rumen:瘤胃;Large intestine 大腸。
3.1 野生林麝瘤胃、小腸和大腸中細菌組成差異
通過對消化道3個區段微生物組成分析發現,厚壁菌門和擬桿菌門為瘤胃的優勢菌門,厚壁菌門能夠分解纖維素并產生機體可以直接吸收利用的短鏈脂肪酸(SCFA),對維持腸道穩態和減少病原菌定植有重要作用[13],擬桿菌門主要作用是降解蛋白質、碳水化合物和脂肪[14];大腸中擬桿菌門和厚壁菌門相對豐度達75%以上,與Li等[6-7]、Hu等[8]對林麝糞便中微生物測序結果一致。相較于瘤胃,大腸中厚壁菌門相對豐度較高,小腸中厚壁菌門相對豐度明顯下降,擬桿菌門幾乎消失,大腸和小腸中厚壁菌門、擬桿菌門的比值大幅上升。Islam等[15]指出,用含膽酸的食物飼喂大鼠會導致回腸厚壁菌門、擬桿菌門的比值大幅增加。林麝等反芻動物膽汁中膽酸含量較高[16],這可能是導致小腸中擬桿菌門相對豐度下降的原因。
變形菌門在小腸中的相對豐度明顯偏高,這與趙文靜等[11]對小鼠小腸細菌觀察到的結果相似。小腸中的優勢菌屬多為致病菌,其中大腸桿菌屬于γ變形菌門中的一種條件致病菌,在宿主免疫系統健全的情況下能刺激小腸免疫系統發育和免疫應答,促進腸上皮細胞分裂并分泌殺菌物質[17-18]。麝在管理不當或受到強烈刺激時會導致免疫力下降,易受到大腸桿菌影響導致腹瀉[19]。林麝小腸細菌中多為病原菌的原因可能是小腸中充滿營養豐富的食糜,是病菌與機體競爭吸收營養物質的主要體外場所。至今為止,對麝消化道中病菌的研究較少,這些病菌在麝免疫系統和消化系統中發揮的作用還需要進一步探究。

Small intestine:小腸;Rumen:瘤胃;Large intestine:大腸;Firmicutes:厚壁菌門;Proteobacteria:變形菌門;Bacteroidrtes:擬桿菌門;Synergistetes:互養菌門;Verrucomicrobia:疣微菌門;Spirochaetes:螺旋體門;Others:其他。
本研究結果顯示,互養菌門是瘤胃的優勢菌門(相對豐度達到10%),在小腸和大腸樣本中沒有檢測到,在先前以糞便為樣本的研究[6-8]中也未發現。有報道稱互養菌門在陜北白絨山羊瘤胃中主要負責降解氨基酸和丙酮酸,在幼年山羊瘤胃中與母乳消化有關[20]。林麝屬于嫩食動物,嫩葉中蛋白質含量豐富,互養菌門細菌可能是林麝為適應高蛋白質食物所保有的共生菌,在氨基酸消化中起重要作用。
在屬水平上,普雷沃氏菌屬、月形單胞菌屬、依賴桿菌屬等厭氧有益菌在瘤胃中可以分解纖維素和糖,產生營養物質,改善胃腸道環境的代謝物[21],對維持消化道穩態起到非常重要的作用。有益菌還能通過刺激免疫系統、消耗空間和氧氣等方法抑制多數需氧致病菌的繁殖[22]。基于以上有助于消化、免疫的野生林麝消化道微生物,后續可以開展麝用益生菌制劑的研究和開發,對于改善圈養林麝的健康狀態有重要意義。
一些細菌在消化道中豐度較低,但對消化道穩態影響較大,如艾克曼嗜黏蛋白菌,通過吞噬胃腸道黏膜中的黏蛋白調整黏液層厚度,其在消化道分布是由黏液層厚度決定的[23]。本試驗僅在瘤胃和大腸中檢測到少量艾克曼嗜黏蛋白菌,這與Li等[6]認為該菌只存在于圈養林麝的推斷不同。Li等[6]推測由于圈養林麝與人接觸多,艾克曼嗜黏蛋白菌可能通過人傳給了圈養林麝,通過本研究結果推測,艾克曼嗜黏蛋白菌可以正常定植在野生林麝的消化道中,但該菌過量繁殖可能會對宿主產生不利影響,例如艾克曼嗜黏蛋白菌繁殖會干擾宿主黏膜平衡,從而加劇傷寒沙門氏菌誘發的腸道炎癥[24]。艾克曼嗜黏蛋白菌在麝消化道中的作用還不清楚,其定植與人為活動、麝胃腸炎之間的關系還有待進一步驗證。
3.2 野生林麝瘤胃、小腸和大腸中古菌、真菌和病毒組成差異
瘤胃中真菌的主要作用是提高細菌對纖維素的利用率并參與細胞壁多糖降解后的發酵[25]。林麝屬于嫩食性動物,其食物中纖維素含量低,所以其瘤胃中真菌數量也低于牛、馬等粗飼動物[26]。麝瘤胃中優勢真菌是根內球囊霉,它是一種植物共生菌,能提高植物抗旱性能和對微量元素的獲取[27],其可能是伴隨植物一起進入瘤胃的。小腸中優勢真菌是黃曲霉,其在林麝小腸中占比較大,這種真菌在無氧條件下不產生黃曲霉素,對牦牛消化道微生物的研究表明其主要抑制擬桿菌門中普雷沃氏菌屬的增殖[28],這從另一個角度解釋了小腸中擬桿菌門相對豐度下降的現象。
瘤胃和大腸中優勢古菌屬是甲烷八疊球菌屬、甲烷短桿菌屬,小腸中優勢古菌屬是亞硝化侏儒菌屬。已有研究顯示圈養林麝糞便中的古菌主要有甲烷短桿菌屬和熱裸單胞菌屬[29],這與本試驗所得大腸結果有差距。甲烷八疊球菌屬是一種可以利用多種途徑產甲烷的古菌,通過消耗氫氣保證消化道正常發酵的進行[30]。造成這種差異的原因可能是由于野生林麝和圈養林麝生存環境不同導致,后續還需要多樣本驗證。
瘤胃和大腸中的病毒多為弧狀噬菌體、地中海噬菌體、肌尾噬菌體這類對宿主無害的噬菌體,它們主要侵染細菌,其中一些會導致細菌裂解。而小腸中多為對宿主有害的致病性病毒,如副痘病毒屬,其能引起動物組織病變、增生[31],也能被腸上皮樹突狀細胞吞噬,將其呈遞給T細胞,進而引發機體的免疫反應[32],對維持正常免疫反應有一定作用。
3.3 野生林麝瘤胃、小腸和大腸中ARGs差異
本研究發現小腸微生物ARGs主要是adeG和TaeA,多編碼膜內轉運體和外排泵,TaeA賦予小腸細菌對泰妙菌素的耐藥性;瘤胃中微生物ARGs主要為Aminocoumarin_resistant_alaS,賦予大腸桿菌對新生霉素的抗性;大腸中微生物ARGs主要編碼外排泵,其賦予大腸對大環內酯類抗生素、維拉帕米、四苯基氯化磷等抗生素的耐藥性。先前對圈養林麝消化道微生物ARGs的研究表明,其對阿莫西林、青霉素、四環素等常見抗生素具有耐藥性[33],這與本研究所得野生林麝消化道ARGs種類差距較大,導致差異的原因可能是生長環境不同。外排泵相關基因的表達是導致細菌多重耐藥性的主要原因[34],小腸和大腸中微生物外排泵相關基因表達會導致宿主多重耐藥性,這提示消化道后半部分中微生物與宿主耐藥性關系更密切,在治療消化道疾病用藥時,應注意避開已經有抗性的藥物以達到最佳治療效果。野生林麝耐藥性主要來自于消化道微生物所攜帶的ARGs,這些微生物的ARGs可能來自于土壤微生物基因的水平交換。本試驗中野生林麝消化道ARGs數據可為圈養林麝消化道微生物ARGs研究做參考。
4 結 論
① 本試驗所采樣的這只野生林麝瘤胃、小腸和大腸中細菌組成差異較大,共有優勢菌門為厚壁菌門和變形菌門。
② 古菌、真菌等在野生林麝瘤胃、小腸和大腸中組成差距較大,占比均相對較少;此外,一些病毒和真菌會對細菌的相對豐度產生影響。
③ 野生林麝大腸和小腸中微生物與宿主的多重耐藥性關系密切,表明這些部位的一些特殊菌群可能在麝免疫反應和耐藥性中起重要作用。
