丹參酮II-A通過調控TLR4/IκBα/NFκB信號通路抑制LPS誘導的細胞炎癥
2021-02-07 06:09:08侯道榮崔斯童
中國藥理學通報 2021年2期
關鍵詞:檢測
侯道榮,劉 振,崔斯童,馬 駿
(1.南京醫科大學醫藥實驗動物中心,江蘇 南京 211166;2.南京醫科大學附屬南京醫院神經外科,南京市第一醫院,江蘇 南京 210006)
炎癥通常被認為是對感染或損傷的防御反應,但是,如果過度或持續,它將導致多種炎癥疾病,如敗血癥、關節炎、癌癥、動脈粥樣硬化和自身免疫性疾病的發生[1]。炎癥的發病機制是一個復雜的過程,受細胞因子和多種促炎性因子的誘導,如前列腺素E2(PGE2)、一氧化氮(NO)、白細胞介素1β(IL-1β)、白細胞介素6(IL-6)和腫瘤壞死因子-α(TNF-α)的調節[2]。在許多急慢性炎癥性疾病中,過度產生的促炎癥介質會進一步加重了炎癥浸潤和免疫反應[3]。因此,這些促炎性介質是開發抗炎藥的重要靶點。
巨噬細胞廣泛分布于體內,在炎癥反應中起重要作用[4]。巨噬細胞通過模式識別受體(PPR)識別致病物質并利用促炎性因子(如IL-1β、IL-6和TNF-α)調節炎癥反應[5]。脂多糖(LPS)是常用的巨噬細胞激活劑[6]。LPS是革蘭陰性菌外膜的主要成分,其誘導的巨噬細胞的激活是TLR4介導的細胞內包含NFκB信號級聯的結果[7]。研究表明[8],NFκB的激活可導致促炎性介質,如誘導型一氧化氮合酶(iNOS)、環氧合酶-2(COX-2)、IL-1β、IL-6和TNF-α的釋放。
丹參酮Ⅱ-A(Tan IIA)是從丹參中提取的脂溶性化合物。已證明Tan IIA在抗炎活性中起重要作用[9]。Tan IIA用于治療各種炎癥反應,如脂多糖誘導的THP巨噬細胞炎癥骨腫瘤、動脈粥樣硬化和心血管疾病的脊髓炎癥[7];Tan IIA能有效抑制大鼠腦缺血/再灌注損傷模型中促炎細胞因子,包括腫瘤壞死因子(TNF)和白介素-6的釋放[10]。本研究將探究Tan IIA對LPS誘導的RAW 264.7巨噬細胞炎癥的具體的作用機制。
1 材料
1.1 細胞株小鼠單核巨噬細胞RAW 264.7細胞購自上海中科院細胞庫。
1.2 藥品與試劑Tan IIA(T4952,美國Sigma公司),使用DMSO 溶液配制;LPS(L2630,美國Sigma公司);DMEM 高糖培養基(11960044,美國Gibco公司)、胎牛血清(10099,美國Gibco公司)、青鏈霉素(15070063,美國Gibco公司)、L-谷氨酰胺(25030081,美國Gibco公司);TNF-α ELISA 檢測試劑盒(EK0527,武漢博士德生物工程有限公司)、IL-6 ELISA 檢測試劑盒(EK0411,武漢博士德生物工程有限公司);IL-1β ELISA檢測試劑盒(AE58504HU,中國Abebio公司)、CCK-8試劑盒(OCPA(C)20132001,中國和元生物技術有限公司);蛋白檢測試劑盒(23225,美國Thermo Fisher Scientific公司);ECL Western blot底物(35055,美國Thermo Fisher公司);MMP-2 兔單克隆抗體(ab92536,1 ∶1 000,英國Abcam公司)、MMP-9兔多克隆抗體(ab38898,1 ∶1 000,英國Abcam公司)、NF-κB 兔單克隆抗體(ab32360,1 ∶1 000,英國Abcam公司)、p-NFκB 兔多克隆抗體(ab28849,1 ∶1 000,英國Abcam公司)、TLR4兔單克隆抗體(ab22048,1 ∶1 000,英國Abcam公司)、IκB-α 兔單克隆抗體(ab32518,1 ∶1 000,英國Abcam公司)、p-IκB-α兔單克隆抗體(ab92700,1 ∶1 000,英國Abcam公司);GAPDH兔單克隆抗體(A19056,1 ∶1 000,中國Abclonal公司);羊抗兔IgG辣根過氧化物酶二抗(ab7090,1 ∶1 000,英國Abcam公司);羊抗鼠IgG辣根過氧化物酶二抗(ab97040,1 ∶1 000,英國Abcam公司)。
1.3 儀器全波長多功能酶標儀(美國PerkinElmer公司),遷移小室(美國Corning公司),二氧化碳培養箱(美國Thermo Fisher公司);光學倒置顯微鏡(德國Leica公司)。
2 方法
2.1 細胞培養將小鼠巨噬細胞RAW 264.7細胞以1×106/孔的密度接種于6孔板中。細胞培養基為DMEM完全培養基添加10% (V/V) FBS,1×105IU·L-1青霉素G,100 kg·L-1鏈霉素和2 mL谷氨酰胺。細胞于5% CO2,37 ℃培養箱中培養。取對數生長期細胞用于實驗。
2.2 分組與干預RAW 264.7細胞以1 × 106/孔的密度鋪于6孔板,鋪板后d 2使用隨機數字表法將細胞板分隨機分為模型組(LPS)、干預組(LPS+Tan IIA)和對照組(DMSO)。LPS組,細胞加入1 mg·L-1LPS培養24 h;LPS+Tan IIA組,細胞加入Tan IIA(終濃度為10 μmol·L-1,Tan IIA溶于DMSO)培養2 h,隨后加入1 mg·L-1LPS繼續培養24 h;DMSO組,細胞加入等體積的DMSO培養24 h。所有實驗重復3次。
2.3 細胞增殖測定RAW264.7細胞活力測定使用CCK-8試劑盒進行。各組細胞更換培養基后,按照CCK-8試劑盒說明書操作。RAW264.7細胞以每孔1 × 104的密度鋪于96孔板中(設置6個復孔),并且使用DMSO、LPS、LPS +Tan IIA處理細胞。72 h的孵育期間,每24 h每孔加入10 μL CCK-8。使用酶標儀測定每孔450 nm的吸光度值(OD)。
2.4 細胞遷移測定將所有細胞培養試劑和遷移板放在37 ℃培養箱溫育;待RAW264.7細胞培養至對數生長期,消化細胞,用PBS和無血清培養基先后洗滌1次,用100 μL無血清培養基懸浮細胞,計數,調整濃度為2×108·L-1;在遷移板下室(即24孔板底部)加入700 μL 含10%血清的培養基,上室加入100 μL細胞懸液,繼續在孵箱培養24 h;用鑷子小心取出遷移檢測板,吸干上室液體,移到預先加入約800 μL甲醇的孔中,室溫固定30 min;取出遷移檢測板,吸干上室固定液,移到預先加入約800 μL Giemsa染液的孔中,室溫染色20 min;輕輕用清水沖洗浸泡數次,取出小室,吸去上室液體,用濕棉棒小心擦去上室底部膜表面上的細胞;用小鑷子小心揭下膜,底面朝上晾干,移至載玻片上用中性樹膠封片;顯微鏡下隨機取9個視野計數,統計結果。實驗組中遷移的細胞數除以對照組中遷移的細胞數,再乘以100%即為相對遷移率。
2.5 細胞炎性因子檢測取對數生長期的RAW264.7 細胞,按5×105/孔接種于6孔板中培養24 h。待其貼壁后棄去上清液,加入試劑繼續培養24 h,然后用1.5 mL離心管收集上清,按照ELISA試劑盒說明書步驟檢測細胞上清液中IL-6、TNF-α、MCP-1和IL-1β的含量。
2.6 Western blot 檢測使用Western blot檢測各組細胞中信號通路相關蛋白MMP-2、MMP-9、NF-κB、p-NFκB、TLR4、IκB-α和p-IκB-α的表達。細胞勻漿后分離蛋白,用含1mmol·L-1PMSF的RIPA冰裂解緩沖液4 ℃孵育10 min,12 000g離心10 min。用Pierce BCA蛋白檢測試劑盒檢測蛋白濃度。然后用SDS-PAGE將同樣定量的蛋白質分離并轉移到0.2 μm PVDF膜上。在室溫下用含3% (W/V) BSA的TBST緩沖液中封閉后,一抗4 ℃孵育過夜。用HRP標記的二抗室溫孵育1 h。使用Pierce ECL Western blot底物檢測蛋白。使用ImageJ軟件定量,使用GraphPad Prism軟件作圖。
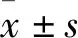
3 結果
3.1 Tan IIA對LPS處理的RAW 264.7細胞增殖的作用LPS能誘導RAW264.7的分化并且抑制RAW264.7的增殖[6]。為了評估丹參酮II-A對RAW 264.7細胞增殖活力的影響,我們使用不同濃度Tan IIA(0、2、5、10、15 μmol·L-1)對RAW 264.7細胞進行處理,培養24 h后,每孔加人10 μL的CCK-8試劑孵育2 h。酶標儀檢測OD450值(Fig 1)。實驗結果表明LPS明顯抑制RAW 264.7細胞的增殖;和LPS處理組比較,Tan IIA(終濃度10 μmol·L-1)處理24 h對可明顯逆轉LPS對RAW 264.7增殖的抑制作用(P<0.05)。因此我們選擇10 μmol·L-1Tan IIA用于后續實驗。
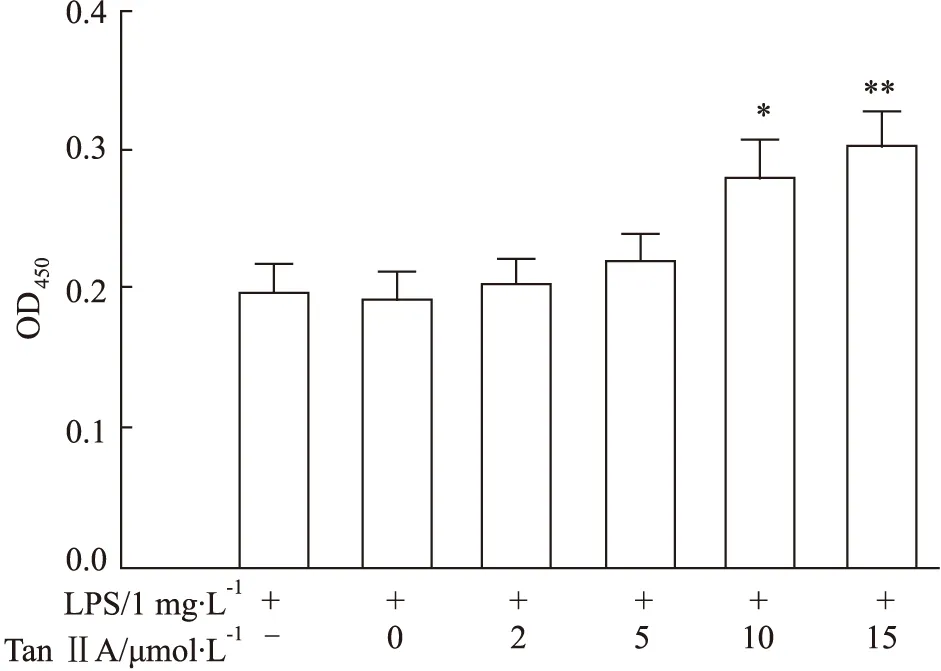
Fig 1 Effects of different concentrations of Tan IIA on RAW264.7 cell viability (,n=6)*P<0.05,**P<0.01 vs LPS group.
3.2 Tan IIA對LPS處理的RAW264.7細胞遷移的作用我們使用細胞遷移實驗檢測Tan IIA對LPS誘導的RAW264.7細胞遷移的作用(Fig 2)。實驗結果表明,LPS處理明顯增加了RAW 264.7細胞的遷移,和DMSO組比較,P<0.01(Fig 2A,B);Tan IIA處理可明顯抑制LPS對RAW 264.7遷移的促進作用,和LPS組比較,P<0.01。
3.3 Tan IIA抑制LPS誘導的RAW264.7細胞炎癥反應RAW264.7細胞培養基上清的4種主要的趨化因子和炎性因子(TNF-α、MCP-1、IL-6和IL-1β)使用ELISA方法檢測(Fig 3)。結果表明,LPS誘導RAW264.7細胞炎性因子TNF-α、IL-6、IL-1β和趨化因子MCP-1明顯增加,和DMSO組比較,P<0.01;Tan IIA處理可明顯降低RAW 264.7細胞的炎性因子水平,和LPS組比較,P<0.01。這些結果表明,Tan IIA可以通過降低RAW264.7細胞炎性因子的產生來抑制LPS引發的炎癥反應。
3.4 Tan IIA通過RAW264.7細胞炎性通路相關蛋白表達的影響我們使用Western blot方法檢測Tan IIA對RAW264.7細胞炎性相關通路蛋白的表達(Fig 4A)。結果表明,與DMSO組比較,LPS誘導MMP-2和MMP-9的明顯升高(P<0.05和P<0.01。而和LPS組比較,Tan IIA明顯抑制LPS誘導的MMP-2和MMP-9的升高(P<0.05 和P<0.01)。各組細胞間NFκB和IκB的表達差異無顯著性。LPS誘導p-NFκB、TLR4和p-IκB的明顯提高,和DMSO組比較,P<0.01。Tan IIA明顯抑制LPS誘導的p-NFκB、TLR4和p-IκB的升高,和LPS組比較,P<0.01。
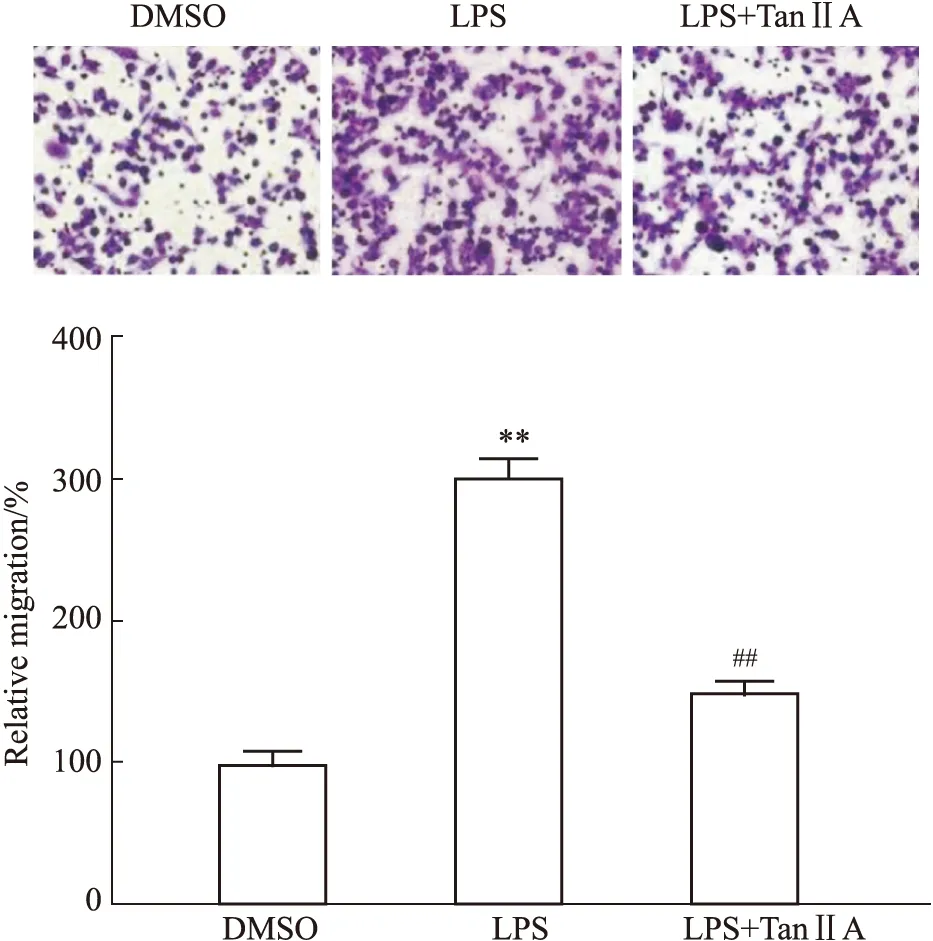
Fig 2 Tan IIA restored migration capacity of LPS-stimulated RAW264.7 cells (,n=6)**P<0.01 vs DMSO group;##P<0.01 vs LPS group.

Fig 3 Tan IIA treatment ameliorated inflammatory cytokine up-regulation in LPS-stimulated RAW264.7 cells(,n=6)The concentration of four inflammatory cytokines IL-1β (A),TNF-α (B),IL-6 (C) and MCP-1 (D) in RAW264.7 cells was examined by ELISA.**P<0.01 vs DMSO group;##P<0.01 vs LPS group.

Fig 4 Tan IIA inhibited protein expression of TLR4/IκBα/NFκB pathway and MMP-2,MMP-9 expression in LPS-stimulated RAW264.7 cells (,n=6)A:Western blot analysis of protein levels in DMSO,LPS or Tan IIA +LPS treated RAW264.7 cells;B:Data are shown as .*P<0.05,**P<0.01 vs DMSO group;#P<0.05,##P<0.01 vs LPS group.
4 討論
小鼠巨噬細胞細胞系RAW264.7被認為是檢測感染相關的促炎性介質最敏感和有效的模型[6]。它可以被LPS觸發,作為研究炎癥反應的經典模型。循環血單核細胞來源的巨噬細胞可參與炎癥反應[11]。RAW264.7細胞是Abelson小鼠白血病病毒轉化的巨噬細胞,是最常用的巨噬細胞實驗模型,在巨噬細胞相關炎癥信號通路的研究中發揮重要作用[3]。近年來,LPS誘導的RAW264.7細胞被廣泛用于篩選有效的抗炎藥物。因此,我們選擇這一典型模型,探討Tan IIA對LPS誘導的RAW264.7炎癥反應的作用和機制。
Tan IIA是丹參的主要脂溶性成分之一,是一種眾所周知的黃酮類化合物,對炎癥反應有較強的抑制作用[12]。Tan IIA對動脈粥樣硬化和相關炎癥反應具有明顯療效[13]。此外,研究顯示,Tan IIA能有效抑制大鼠腦缺血/再灌注損傷模型中促炎性細胞因子,包括TNF-α和IL-6等的釋放[10]。最近的研究表明,Tan IIA可以減少博萊霉素和LPS誘導的大、小鼠肺的炎癥細胞浸潤、促炎性細胞因子釋放和膠原沉積[14]。因此,Tan IIA被認為是一種很有前景的抗炎藥物。在本研究中,Tan IIA能有效抑制LPS誘導的RAW264.7細胞活力的下降、細胞遷移和TNF-α、IL-6、MCP-1、MCP-9等炎性因子的釋放和表達。
為了探究Tan IIA抗炎性作用的具體機制,本研究檢測了RAW264.7細胞炎性相關蛋白的表達。Tan IIA 能抑制LPS誘導的MMP-2和MMP-9的高表達。單核細胞趨化蛋白酶(MMPs)可特異的降解細胞外基質的主要成分Ⅳ型膠原,對細胞的遷移、侵襲、活化及血管重塑等均起著重要作用[15]。MMP-2和MMP-9增多可使細胞基質膜成分降解,介導單核細胞等穿過血管,在降解的細胞外基質中游走,向靶器官浸潤,促進炎癥反應及組織損傷[16]。MMP對細胞的遷移、侵襲、活化及血管重塑等均起著重要作用。有研究顯示,MMP-2和MMP-9高表達與卵巢癌的生長和侵襲轉移密切相關。因此,本研究表明Tan IIA可通過抑制MMPs的表達來發揮抑制巨噬細胞RAW264.7細胞炎性的作用。
此外,LPS還誘導TRL4、p-NFκB和p-IκB的高表達,而這些可以通過Tan IIA治療逆轉。因此,我們可以推斷Tan IIA通過抑制TRL4/IκB/NFκB信號通路發揮抗炎性作用。這表明,抑制RAW 264.7巨噬細胞IκB活化從而抑制NFκB信號通路,可調節LPS對炎癥介質和細胞因子的誘導作用。研究表明,抑制NFκB活性,可改善類風濕性滑膜的炎癥反應和組織破壞[2]。在結腸炎模型中,對NFκB的抑制可通過下調NFκB介導的促炎性細胞因子改善結腸炎癥損傷[17]。TLR4是Toll樣受體,是一種能夠識別LPS與細胞表面的髓樣分化因子2(MD2)的模式識別受體。TRL4被LPS激活后可磷酸化IκB-α,然后使NFκB磷酸化,從而使NFκB進入細胞核[18]。IκB-α磷酸化導致其降解并允許NFκB進入細胞核。NFκB的啟動子活性主要是由核移位和NFκB磷酸化引起的。NFκB磷酸化激活導致細胞因子和炎癥介質,如TNF-α、IL-1β、IL-6,MCP-1、MMP-9等的釋放。
綜上所述,本研究闡明Tan IIA可以通過調控TLR4/IκB-α/NFκB 炎癥信號通路,抑制LPS誘導的炎癥因子IL-6、IL-1β、TNF-α 和MMPs的表達來發揮抗炎活性,為抗炎性藥物篩選提供了實驗依據。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48