土壤干旱及復水對側柏葉綠素熒光參數的影響
2021-02-05 09:06:16張玉玉王進鑫呂國利馮樹林
水土保持研究 2021年2期
關鍵詞:差異
張玉玉, 王進鑫, 馬 戌, 呂國利, 馮樹林
(1.西北農林科技大學 水土保持研究所, 陜西 楊凌 712100; 2.西北農林科技大學資源環境學院, 陜西 楊凌 712100; 3.農業部 西北植物營養與農業環境重點實驗室, 陜西 楊凌 712100)
由于自然界的降雨脈沖及人為灌溉等原因,不僅在干旱半干旱地區經常會出現土壤缺水和干濕交替的現象,而且在南方等非干旱地區也經常出現季節性干旱。干旱脅迫是影響植物生長、生理生化的主要限制因子。當植物處于干旱環境時,會通過調動自身的生理生化過程或啟動自我的保護機制來避免自身受到損傷[1-2]。植物當前的生長狀態不僅與過去的水分歷時有關,而且與當前所處的水分環境有關。Acevedo等[3]研究發現,植物經受一定強度的干旱脅迫后復水,因干旱造成的生長抑制及生物量的損失等可以彌補,復水后植物會出現短暫的加速生長。側柏[Platycladusorientalis(L.) Franco]屬常綠喬木,常為陽坡造林樹種,也是常用的園林綠化及困難立地植被恢復樹種。適應性強,對土壤要求不嚴,在酸性、中性、石灰性和輕鹽堿土壤中均可生長。既耐干旱瘠薄,又喜生于濕潤肥沃排水良好的鈣質土壤,除青海、新疆外全國均有分布。因此,本文研究耐旱喜濕的典型造林樹種側柏經干旱脅迫后復水對其葉綠素熒光參數影響。
在正常的環境條件下,植物葉片葉綠素所吸收的太陽光能通過3個途徑來消耗,分別是光合電子傳遞、葉綠素熒光發射和熱耗散,而這3個途徑之間存在著一定的彼此消長的相互關系,光合作用和熱耗散的變化都會引起葉綠素熒光參數相應的變化[4]。作為聯系環境和植物光合作用的內在指標葉綠素熒光參數,近些年來,廣泛作為研究植物在干旱、高溫、低溫、強光等逆境脅迫條件下抗逆性指標的有效探針[5-10]。在干旱脅迫的條件下,葉綠素熒光參數會發生一定的變化,Guo等[11]對黑枸杞(LyciumruthenicumMurr)經干旱脅迫后的研究發現NPQ增加,最大熒光、最大量子產量等降低,說明干旱脅迫影響了PSⅡ反應中心的開放性。Lang等[12]研究連翹(Forsythiasuspensa)幼苗經土壤干旱脅迫后,RSWC從51.84%下降到37.52%時,NPQ有增加的趨勢,當RSWC低于37.52%時,Fv/Fm,Yield,NPQ和qP顯著降低。劉筱等[13]研究干旱脅迫對何首烏(PolygonummultifloraThunb.)葉綠素熒光參數的影響表明,隨著干旱脅迫的加劇,Fv/Fm和ETR值呈現出先升后降的趨勢,而NPQ則呈現降低趨勢。楊文權等[14]研究干旱及復水對小冠花(CoronillavariaL.)葉綠素熒光參數的影響顯示,干旱脅迫天數過長及脅迫程度過大不僅會影響小冠花的光合作用,還會使小冠花的光反應中心受損;但由于小冠花有一定的抗旱性,脅迫解除后,光合作用能很快恢復至正常水平,葉綠素熒光參數(Fv/Fm,Yield,ETR,NPQ等)也能快速恢復到原始的水平。
植物在生長發育過程中,均呈現出一定的階段性變化規律,且在不同生長階段對環境的要求也存在著明顯的異質性[15-16]。王進鑫等[17]研究發現,對側柏幼樹蒸騰耗水影響較大的是生長盛期的土壤供水狀況。因此,側柏幼苗在生長盛期對土壤水分的響應更加明顯。張慕黎[18]研究側柏經干旱脅迫后光合速率隨著脅迫程度的加劇逐漸降低,復水后脅迫程度越小其恢復能力越強。龐云龍[19]研究側柏幼樹光合速率、蒸騰速率在復水24 h后均有不同程度的恢復。目前有關側柏生長與土壤水分之間關系的研究,多以光合作用的研究為主,而對光合作用內在探針即葉綠素熒光參數的研究較少。因此,本文研究生長盛期側柏幼苗經過不同程度土壤干旱脅迫及旱后復水對葉綠素熒光參數的影響,分析葉綠素熒光參數與土壤干旱及旱后復水之間的關系,掌握側柏幼苗對干旱脅迫的響應機制,進一步反映土壤干旱及旱后復水對側柏光合作用的影響,從而為側柏在逆境條件下更好的生長提供技術支撐。
1 材料與方法
1.1 試驗地點和材料
試驗在陜西楊凌西北農林科技大學南校區防雨旱棚(108°07′E,34°16′N)內進行,采用旱棚人工控水的方法進行布設。該地區屬于暖溫帶半濕潤大陸性季風氣候,降水季節分配不均勻,有78%的雨量集中在5—9月,且時有暴雨出現。年平均相對濕度70%左右,年平均氣溫12.9℃,1月平均氣溫-1.2℃,7月平均氣溫26.0℃,≥10℃年有效積溫4 169.2℃,年干燥度1.1。
試驗選取地徑、株高和根系基本一致的1 a生側柏幼苗,沖洗根系后用吸水紙吸干苗木表面水分,記錄地徑、株高和鮮重,于2018年3月15栽植在口徑29 cm、深27 cm統一規格的塑料桶中,每桶栽植2株,并對其進行編號。栽培基質為土,通透性良好,田間持水量為22.3%,充分風干后,進行過篩,除去石礫、結核及林草根系,充分混勻后每盆裝10 kg。采用邊埋邊壓邊提苗的方式栽植,并在土層表面鋪蓋2 kg石礫以減少土體表面蒸發。澆灌一定體積的水,以保證苗木成活。然后放置室外自然透風塑料旱棚內,使側柏幼苗盆栽土壤水分維持在田間持水量的80%~90%。
1.2 試驗設計
本試驗土壤干旱脅迫程度參照王進鑫[17],吉增寶[20]等的研究,設置4個土壤水分梯度[土壤相對含水量(RSWC)為40%,52.16%,70%,87.84%],每個處理4個重復,另設一個對照(RSWC為100%,充分供水)。干旱脅迫外的所有時間內按照100%相對含水量對各處理供水,試驗期間栽培基質保持自然肥力,不施肥。整個試驗過程中,以大棚內自然溫度和光照為主,采用人為適時控制補水的辦法,使土壤水分達到試驗要求。
2018年6月15日開始對側柏幼苗盆栽進行水分控制,將表面的石礫去掉,增大土壤蒸發或適當補水,使其在6月20日的土壤相對含水量達到40%,52.16%,70%,87.84%,對照組幼苗盆栽土壤相對含水量保持100%。采用稱重法對側柏盆栽進行控水處理,歷時60 d。于8月21日上午8∶30—11∶30對其葉綠素熒光相關指標進行復水前測定,8月22日早晨6∶30對各處理進行復水(按土壤相對含水量100%復水),分別于復水后2,24,48,72 h對其指標進行測定。
1.3 葉綠素熒光參數測定
使用德國WALZ公司生產的便攜式調制葉綠素熒光儀mini-PAM2500對側柏葉片進行活體測定。試驗于8月21日上午8∶30開始,測定前先對各處理進行暗適應30 min,然后分別測定其相應指標。用弱測量光測定初始熒光Fo[強度低于1 μmol/(m2·s)],接著照射飽和脈沖光[強度大于7 000 μmol/(m2·s),0.8 s]測得最大熒光Fm,根據Fo和Fm計算PSⅡ的最大光化學量子產量Fv/Fm=(Fm-Fo)/Fm。等熒光恢復到Fo附近打開光化光[606 μmol/(m2·s)]進行熒光誘導,約4~5 min熒光達到穩態測量,再次打開飽和脈沖測量即時熒光(F)、即時最大熒光(Fm′),根據F和Fm′計算PSⅡ實際光化學量子產量Yield=(Fm′-F)/Fm′。相對光合電子傳遞速率(ETR)和非光化學淬滅系數(NPQ)可直接讀取。
1.4 數據處理
數據統計分析采用Microsoft Excel和SPSS軟件結合進行,使用單因素方差分析(one-way ANOVA),并運用Duncan檢驗方法來確定每個指標在各處理間差異的顯著性。
2 結果與分析
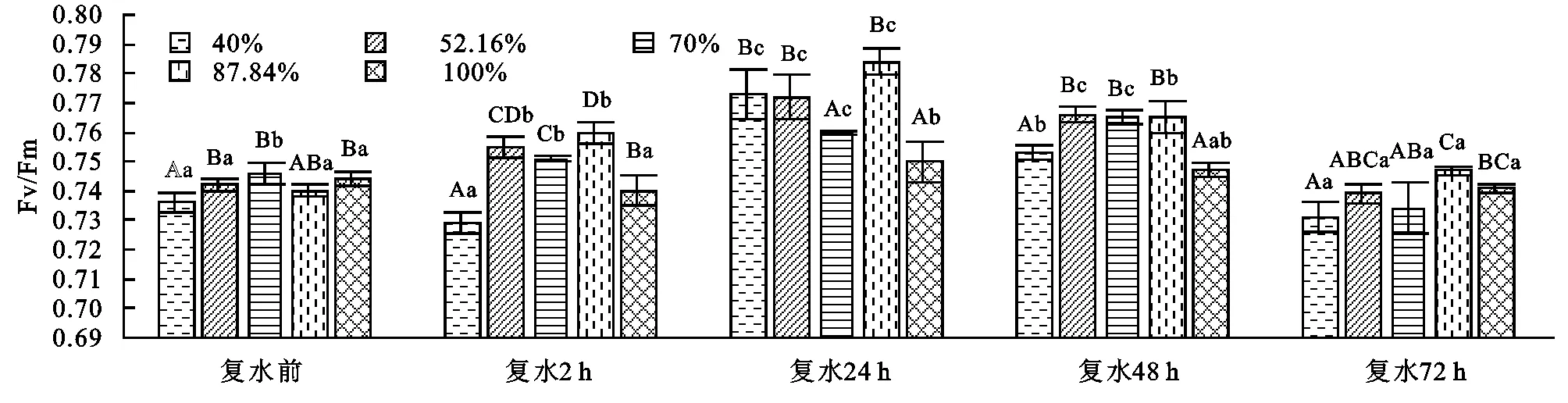
2.1 側柏幼苗最大光化學量子產量Fv/Fm對土壤干旱及旱后復水的響應
葉綠素熒光參數Fv與Fm的比值(Fv/Fm)是PSⅡ最大光化學量子產量,能反映PSⅡ反應中心最大原初光化學量子效率,在正常的環境條件下生長的植物,Fv/Fm值的變化范圍極小,Fv/Fm是反應光抑制程度的可靠指標和良好的探針[21]。由圖1可以看出,側柏幼苗Fv/Fm隨著土壤干旱脅迫程度的加劇呈現先降后升再降的趨勢,各處理差異顯著(p<0.05)。復水后,不同干旱脅迫條件下側柏幼苗Fv/Fm隨著復水時間的延長呈現先增大后減小的趨勢,在復水24 h除70%的處理其他處理都達到最大值,之后緩慢降低。70%的處理在復水48 h達到最大值,與對照差異顯著,比對照增加了2.4%。而復水72 h重度脅迫(40%)比對照下降了1.3%,與對照差異顯著,這說明側柏幼苗Fv/Fm對復水做出了短暫的恢復,而輕度脅迫87.84%的處理仍比對照高了0.8%,與對照差異不顯著。復水2~48 h各處理與對照相比差異極顯著(p<0.01),復水72 h各處理與對照差異顯著(p<0.05),同一脅迫處理在不同的復水時段差異極顯著(p<0.01)。說明側柏幼苗對復水能做出及時的響應,隨著復水的進行各處理及時恢復并超過對照,出現補償效應。
2.2 側柏幼苗實際光化學量子產量Yield對土壤干旱及旱后復水的響應
實際光化學量子產量Yield是PSⅡ實際光化學量子效率,反映了PSⅡ反應中心有部分關閉的情況下實際原初光能捕獲效率,即用于光化學反應所占PSⅡ吸收光能的比例。復水前側柏幼苗Yield隨著土壤干旱脅迫程度的加劇逐漸降低,其中重度脅迫40%的處理比對照降低了4.9%,各處理與對照差異極顯著(p<0.01)。隨著復水時間的延長,各脅迫處理Yield先增大后減小,其中土壤相對含水量為87.84%的處理在復水24 h達最大值比對照上升了2.3%,與對照差異顯著,在復水48 h其他脅迫處理下Yield達到最大值,其中70%的處理比對照升高了1.4%,與對照差異顯著(圖2)。Yield值越大,說明光能轉化效率越高,形成的光合產物就越多。對側柏幼苗Yield進行方差分析發現,不同復水時段各處理與對照差異極顯著(p<0.01),同一脅迫處理在不同的復水時段差異顯著(p<0.05)。說明土壤干旱脅迫在側柏幼苗的耐旱閾值內,幼苗Yield經土壤干旱脅迫后復水都得到了相應的恢復并在復水48 h各處理出現了補償效應,之后出現降低的趨勢。
注:圖中大寫字母表示同一時間各處理差異的顯著性(p<0.05),小寫字母表示同一處理不同恢復時間差異的顯著性(p<0.05)。
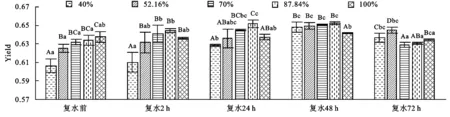
圖2 不同程度土壤干旱及旱后復水對側柏幼苗Yield的影響
2.3 側柏幼苗相對光合電子傳遞速率ETR對土壤干旱及旱后復水的響應
相對光合電子傳遞速率ETR是光合機構吸收光能之后發生電荷分離,產生的電子,沿著電子傳遞鏈向下傳遞的速率。由圖3可以看出,側柏幼苗ETR值隨著土壤干旱脅迫程度的增加呈現出先降后升再降的趨勢,各處理與對照差異極顯著(p<0.01),其重度脅迫40%的處理比對照下降了4.5%。復水后ETR值呈現先增大后減小的趨勢,在復水48 h各處理ETR達最大值,其中重度脅迫40%的處理比對照增加了1.2%,與對照差異顯著,在復水72 h后又呈現下降的趨勢。在復水2 h和24 h隨著復水的進行各處理與對照差異極顯著(p<0.01),復水48 h和72 h各處理與對照差異顯著(p<0.05),同一脅迫處理在不同的復水時段差異極顯著(p<0.01)。說明側柏幼苗隨著土壤干旱脅迫程度的加劇,光合作用逐漸下降,而這種脅迫對側柏幼苗沒有造成不可逆的損傷,隨著復水的進行,幼苗能及時調節自身的生理機制,使其快速恢復并在復水48 h各處理超過對照出現補償效應。
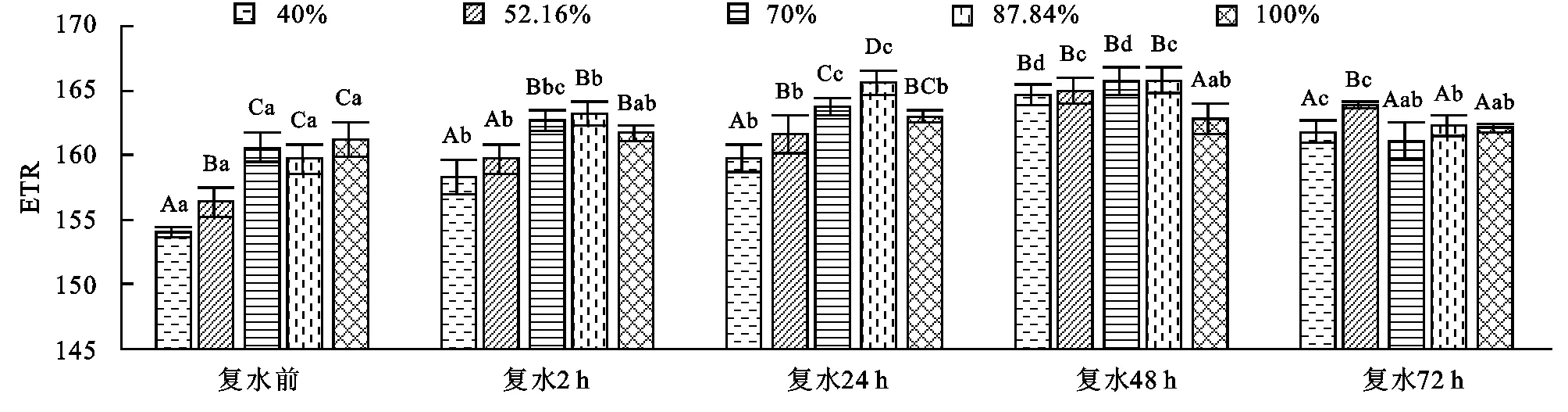
圖3 不同程度土壤干旱及旱后復水對側柏幼苗ETR的影響
2.4 側柏幼苗非光化學淬滅系數NPQ對土壤干旱及旱后復水的響應
非光化學淬滅系數NPQ是PSⅡ反應中心天線色素吸收的光能不能用于光合電子傳遞,而以熱的形式耗散掉多余的那部分光能的比例,熱耗散對植物光合機構免受破壞起到積極的作用。由圖4可以看出,復水前隨著土壤干旱脅迫程度的加劇,側柏幼苗NPQ值逐漸升高,其中重度脅迫40%的處理比對照增加了22.6%,與對照差異極顯著(p<0.01)。NPQ增大說明側柏以增加熱耗散的方式來降低光抑制對其產生的傷害,NPQ的基礎是天線復合體的結構重組,作為一個重要的光保護機制,廣泛的存在于植物體和各種藻類中[22]。隨著復水時間的推移,NPQ值整體上呈現降低的趨勢,其中40%的處理在復水2 h和24 h這兩個時間段呈現上升的趨勢,這可能是由于重度脅迫使側柏幼苗光保護機制破壞嚴重,復水后幼苗沒有及時恢復或NPQ對復水響應比較緩慢的原因,在復水48 h后開始降低,72 h低于對照。隨著復水的進行,各處理與對照差異極顯著(p<0.01),同一脅迫處理在不同的復水時段差異極顯著(p<0.01)。說明側柏幼苗經過不同程度的土壤干旱脅迫后NPQ受到影響,但是隨著復水的進行,這種影響都是可以消除的。

圖4 不同程度土壤干旱及旱后復水對側柏幼苗NPQ的影響
3 討論與結論
3.1 討 論
植物葉綠素熒光參數作為光合作用內在的有效探針,是研究植物抗旱性的良好指標。葉綠素熒光技術檢測到的植物發出的葉綠素熒光信號包含了大量的光合作用信息,而這些信息與植物本身受脅迫程度和自身的營養關系密切[23]。葉綠素熒光技術所檢測到的葉綠素熒光信息主要是來自于植物葉綠體PSⅡ反應中心天線色素蛋白復合體中的葉綠素a分子[24]。
Fv/Fm是PSⅡ反應中內稟光能的轉換效率,能反映PSⅡ利用光能的能力。結果表明,隨著土壤干旱脅迫程度的增大,側柏幼苗Fv/Fm值呈現先降后升再降的趨勢,不同土壤干旱脅迫對側柏幼苗Fv/Fm都有影響,重度脅迫40%的處理與對照差異顯著,其他處理差異不顯著,這可能是因為側柏耐旱性較強,不同程度土壤干旱脅迫60 d對側柏幼苗Fv/Fm的影響也不是很明顯,只有重度脅迫對幼苗Fv/Fm影響顯著。這一結果與董果等[25]對側柏葉綠素熒光特性的研究結果一致。盧福順等[26]研究表明只有在重度干旱脅迫的條件下,Fv/Fm才會出現顯著的變化。復水后,側柏幼苗Fv/Fm呈現先增加后減小的趨勢,在復水24 h各處理出現補償效應,說明即使重度脅迫對側柏幼苗Fv/Fm有顯著的影響,但這種影響是可以通過復水來消除的,所以土壤干旱脅迫對側柏幼苗Fv/Fm的影響是可逆的,這與李曉梅等[27]對兩種辣椒(CapsicumannuumL.)葉綠素熒光參數的研究一致。
Yield能反映植物對吸收的太陽光能的轉換能力。經過不同程度土壤干旱脅迫后,側柏幼苗Yield隨著脅迫程度的加劇而下降,由此說明不同程度土壤干旱脅迫使得側柏幼苗PSⅡ反應中心受到一定的損傷,抑制了光合作用的原初反應,這與孫志勇等[28]對雜交鵝掌楸(Liriodendronchinense×L.tulipifera)和張國盛等[29]對6種樹葉綠素熒光參數的研究結果一致。復水后側柏幼苗Yield值不斷升高,并出現了補償現象,在復水72 h開始緩慢下降,這可能是因為復水后測定時間比較短,側柏幼苗耐旱性較強,對復水做出了短暫的恢復響應后,隨著長時間的復水,幼苗在生理上又做出了不同的響應機制,這一結論還需后期進一步驗證。
葉綠素熒光參數ETR能反映光合機構運轉調節的規律。結果發現在重度脅迫的條件下,側柏幼苗相對光合電子傳遞速率ETR值急劇下降,這一結果與馬仁義等[30]的研究一致,表明側柏幼苗的生長對土壤水分有一定的需求。復水后逐漸恢復并超過對照,復水72 h出現了下降的趨勢,說明側柏幼苗PSⅡ反應中心因土壤干旱脅迫而遭到破壞,光合速率降低,復水后幼苗能及時調整自身光保護機制,使其逐漸恢復。這一結果與蒙祖慶等[31]對油菜(BrassicanapusL.)葉綠素熒光參數的響應和王振夏等[32]對楓楊(PterocaryastenopteraC. DC.)幼苗經土壤濕干交替變化葉綠素熒光的響應結果一致。
植物非光化學淬滅NPQ對植物光合機構免受破壞起到了積極的作用。試驗結果表明,側柏幼苗NPQ隨著土壤干旱脅迫程度的加劇呈現逐漸上升的趨勢,其中87.84%與對照差異不顯著,其他處理與對照差異顯著。在復水后有緩慢的下降趨勢。而在復水2 h和復水24 h經過重度脅迫40%處理的側柏幼苗NPQ依然在上升,這可能是由于重度脅迫使側柏幼苗光保護機制破壞嚴重,復水后沒有及時恢復,或者幼苗NPQ對復水響應比較緩慢的原因,這一結果還有待驗證。復水48 h之后幼苗NPQ開始逐漸降低,說明不同程度的土壤干旱脅迫對側柏幼苗NPQ有明顯的影響,隨著復水的進行,各處理及時恢復。
植物葉片的葉綠素熒光參數是研究植物抗逆性的可靠指標,能快速有效地反映植物對土壤水分變化的響應,本試驗側柏幼苗經60 d不同程度土壤干旱脅迫后,通過增加熱耗散(NPQ)與降低原初光能轉換效率(Fv/Fm)和相對光合電子傳遞速率(ETR)來實現對PSⅡ反應中心的保護。有學者[18]研究發現生長盛期側柏光合速率、蒸騰速率隨著土壤干旱脅迫程度的加劇逐漸降低,復水72 h后光合速率的恢復度呈現出輕度脅迫處理大于重度脅迫,蒸騰速率在土壤相對含水量為52.16%的處理恢復度最大為0.69。而側柏幼苗葉綠素熒光參數在復水后得到恢復,所有處理在復水48 h出現補償效應,說明側柏幼苗經過不同程度土壤干旱脅迫后,對光合機構造成了一定的破壞,但由于側柏耐旱性較強,前期干旱脅迫對其沒有造成不可逆的破壞,通過后期復水的進行得到恢復,并出現補償現象。
3.2 結 論
經過不同程度土壤干旱脅迫60 d的側柏幼苗,Fv/Fm隨著土壤干旱脅迫程度的增大呈現先降后升再降的趨勢,但不同程度土壤干旱脅迫對側柏幼苗Fv/Fm影響較小,復水后得到了補償,在復水24 h和48 h各脅迫程度出現了補償現象,其中87.84%的處理在復水24 h比對照升高了4.5%,與對照差異顯著,出現超補償現象。Yield隨著脅迫程度的加劇而下降,復水后Yield不斷升高,在復水48 h出現了超補償現象,復水72 h開始緩慢下降。在重度脅迫的條件下,側柏幼苗ETR急劇下降,40%的處理比對照降低了4.5%,與對照差異顯著。復水后逐漸恢復并超過對照,復水72 h出現了下降的趨勢。NPQ隨著干旱脅迫程度的增加呈現上升的趨勢,復水后有了緩慢的下降,而在復水2 h和24 h經過重度脅迫(40%)的側柏幼苗NPQ依然在上升,復水48 h開始緩慢下降。說明不同程度的土壤干旱脅迫對側柏幼苗PSⅡ反應中心有一定的損傷,降低了PSⅡ原初光能轉化效率,抑制PSⅡ反應中心的光化學活性,進而破壞了側柏幼苗光合作用的原初反應過程。但這種破壞是可以通過復水來彌補的。因此側柏幼苗PSⅡ能夠耐受較長時間的干旱脅迫,從而為側柏能在逆境條件下更好的生長提供技術支撐。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
