大口黑鱸的卵巢發育周年變化及反季節繁殖研究
2021-02-04 06:39:26崔慶奎沈志剛齊飄飄黃紅紅樊啟學
水生生物學報 2021年1期
崔慶奎 沈志剛 田 宇 齊飄飄 黃紅紅 劉 洋 陳 敏 于 躍 樊啟學
(華中農業大學水產學院,農業農村部淡水生物繁育重點實驗室,教育部長江經濟帶大宗水生生物產業綠色發展工程研究中心,池塘健康養殖湖北省工程實驗室,武漢 430070)
大口黑鱸(Micropterus salmoides),俗稱加州鱸,又稱黑鱸,隸屬鱸形目(Perciformes)、鱸亞目(Percoidei)、太陽魚科(Centrarchidae)、黑鱸屬(Micropterus),原產于北美洲,是當地重要的游釣魚類[1—3]。自1980年引入廣東后,因其肉質鮮美,無肌尖刺,生長迅速,養殖周期短等優勢,迅速成為國內重要的養殖品種[4,5]。近些年對大口黑鱸營養需求的研究不斷深入,一系列配合飼料應運而生,打破了傳統投喂餌料魚的養殖模式,養殖產量急速上升,從2012年的9.6×107kg增長至2018年的43.2×107kg,7年時間增長了3.5倍。有學者便推斷大口黑鱸憑借其獨特優勢,未來20年內,產量有望突破100×107kg,養殖前景十分廣闊[5]。然而華中地區大口黑鱸養殖產業的發展并非一帆風順,現階段主要存在三個問題:一是周年價格波動較大,2019年本地區大口黑鱸塘口價(收購價)最高出現在8月底,高達24.0元/斤,最低出現在1月底,僅有10.5元/斤(圖1),此價格變化規律與此前2015—2018年價格規律非常一致[6];二是地區發展極不平衡,2018年廣東省大口黑鱸產量占全國總產量的59.8%,蘇浙地區占21.0%,而華中地區作為我國水產養殖最重要的區域,產量僅占2.8%[6—8](2019中國漁業統計年鑒)。三是本地區氣候條件不利于正常季節大口黑鱸的養殖,對于1齡苗種而言,下塘后不久便經歷高溫期(6—9月),停止攝食,不利于其快速生長,使得越冬前大部分魚達不到上市規格,且積溫過高,加速其性腺發育成熟[9—11],而越冬期水溫又較低,持續時間長,回暖較慢,回暖后性腺快速發育又消耗大量能量,生長緩慢[12]。

圖1 華中地區大口黑鱸價格周年變化Fig.1 Annual price fluctuation of largemouth bass prices in central China
解決以上問題,目前最有效的方法是培育反季節大口黑鱸苗種,即讓親本一年繁殖兩次,除了4和5月份自然繁殖外,通過一系列技術措施,使其在9和10月份二次產卵,從而培育大規格越冬苗種,將上市時間大大提前,填補市場上7—9月的缺魚期,同時,反季節苗種也將有助于解決因苗種短缺導致的地區發展不平衡的問題,最重要的是反季節苗種能充分利用華中地區氣候條件,出苗后便迎來第一個最適生長期(秋季),越冬回暖后迎來第二個快速生長期(春季),這種養殖模式能提高其生長速度和飼料利用效率[13,14]。
大口黑鱸的反季節繁殖,關鍵在于如何讓性腺退化后的親本再次啟動發育。在自然條件下,大口黑鱸一年只有一個繁殖期[15],陳建酬等[16]曾報道,大口黑鱸產卵季節在3月中旬到4月中旬,Beamish等[17]認為,大口黑鱸產卵季節為7—9月,而Gross等[15]認為其產卵季節為1月中旬至5月底,可見,大口黑鱸的適應能力極強,不同地區差異很大。我們為開展大口黑鱸反季節繁殖,有必要研究清楚本地區其性腺發育規律和環境因子對其性腺的影響規律等,進而確定科學、高效的技術措施。團隊前期利用水庫底層低溫水進行了規模化的大口黑鱸反季節繁殖實驗,通過程序性降溫-低溫維持-升溫措施,成功在9和10月繁育出大口黑鱸苗種[6]。為了深入了解反季節繁殖過程中性腺發育規律、控制成本和尋求最優程序性溫度控制等,特開展此研究。
在本研究中,我們就大口黑鱸性腺周年發育規律做了詳盡的研究,并結合環境因子進行分析,初步得出溫度和光照對其性腺啟動發育和產卵繁殖的影響規律,并基于這些規律,探究了大口黑鱸反季節繁殖的方法。
1 材料與方法
1.1 實驗材料
實驗魚來自湖北黃優源漁業發展有限公司,來源是2017年從廣東大口黑鱸繁育基地購買的新品種“優鱸1號”,實驗期間養殖在面積4000 m2、水深2 m的池塘中,養殖數量為3000尾左右。反季節繁殖實驗開始前從池塘隨機挑選體質量在400—800 g的親魚進行實驗。
1.2 參數測量
實驗魚捕回后置于100 mg/kg的MS-222(間氨基苯甲酸乙酯甲磺酸鹽)水桶中麻醉,每次采3尾魚,主要測定項目包括體長(BL)、體質量(BW)、性腺重(GW)、肝臟重(LW)和腸系膜脂肪重(MFW),并分別以性腺重量、肝重量和腸系膜脂肪重量相對于體質量的百分數來表示性腺指數(GSI)、肝體比(HSI)和腸系膜脂肪系數(MFI)。
絕對繁殖力(F)是在繁殖產卵前隨機選取10尾雌魚進行測定,每尾取0.1—0.5 g卵巢,固定后置于解剖鏡下計數,推算出卵巢總懷卵量,并以絕對繁殖力比上體質量和體長表示相對體重繁殖力(FW)和相對體長繁殖力(FL)。
池塘水溫采用連續溫度記錄儀(分辨率0.1℃)記錄,探頭深度為水面下60 cm,每天記錄6次,取平均值表示當天水溫。光照時長以世界時間網(https://time.guowaitianqi.com/)公布的武漢地區日出、日落時間為準,計算每天光照時長。
1.3 組織學觀察
在親魚解剖后,將性腺樣品置于Bouin’s固定液中固定48h后轉移至70%的乙醇中保存,經過酒精梯度脫水后,用二甲苯透明,石蠟包埋,連續切片厚度為6—8 μm,切片采用蘇木精-伊紅染色(HE染色),用Axio Imager A2顯微鏡拍照觀察。參照Gross等[15]對大口黑鱸卵巢發育特征的描述進行時期劃分,統計卵巢中不同時相卵母細胞數量比時,各采樣時間隨機選取10張玻片,每張玻片隨機挑選一個視野統計該視野下各時相卵母細胞數量比,最后以平均值表示各時相卵母細胞數量比。
1.4 卵巢發育周年樣品的采集
卵巢發育周年樣品的采集從2019年1月23日開始到2020年1月22日結束,根據不同時間卵巢的發育快慢,分為繁殖前期、繁殖期、退化階段、啟動發育階段和越冬期共五個階段進行采樣,共計采樣15次。
繁殖前期采樣時,除采集常規樣品外,還需統計親魚的絕對繁殖力。
繁殖期提前在池塘鋪設產卵網片(網片由棕櫚樹皮編織而成,大小20 cm×30 cm),每天早晚各收集一次,記錄網片數目,每天隨機挑取3片統計產卵量,之后轉移至孵化池孵化。
1.5 大口黑鱸反季節繁殖
2019年9—11月進行反季節親本培育實驗,實驗在3個相同水泥池(面積10 m×10 m×1.2 m)中進行,水深1 m,池頂架設可調節的遮陽網,分別為控溫組、促熟組和對照組,每組隨機從池塘挑選80尾親魚進行實驗。實驗魚轉入水泥池后暫養1周實驗,開始實驗前采樣1次。實驗期間每天飽食投喂,各組間保持一致,并加強水質管理,定期吸污換水,溶氧控制在(6.5±1.0)mg/L。
控溫組:該組采用地下水(常年水溫19—21℃)進行控溫處理。第一步是降溫處理,水溫由29—30℃逐漸降至21℃,歷時30d,期間為保證降溫效果,正午時進行部分遮蔭處理,降溫結束后采樣1次,記錄性腺發育狀況。之后維持水溫20—21℃繼續培育,直至繁殖產卵,期間每10天采樣1次,觀察性腺發育狀況。
促熟組:該組不進行控溫處理,僅對親本進行連續、低劑量的外源性激素注射,探究激素處理對親本性腺啟動發育的影響。主要激素有促黃體素釋放激素類似物(LHRH-A2)+馬來酸地歐酮(DOM)+魚用絨毛膜促性腺激素(HCG),劑量為10 μg/kg的LHRH-A2、5 mg/kg的DOM、700 IU/kg的HCG,每隔10天在胸鰭基部注射一次,采樣時間與控溫組相同。
對照組:該組不作任何處理,使親魚性腺自然發育,期間的投喂、水質管理、采樣時間等均與實驗組相同。
1.6 數據分析
采用SPSS(IBM SPSS Statistics,Version 24)統計軟件進行實驗數據的統計學分析,采用單因素方差分析(One-way ANOVA)進行顯著性檢驗,然后進行用Duncan’s多重比較來確定組間差異的顯著性,P<0.05表示差異具有統計學意義。采用皮爾遜相關系數(Pearson correlation coefficient)分析各組數據的相關性。采用Origin 2017繪圖軟件進行圖表制作。
2 結果
2.1 溫度與光照時長變化對大口黑鱸卵巢周年發育規律的影響
大口黑鱸雌魚GSI與水溫、光照的周年變化規律如圖2所示,可見,華中地區周年水溫在5.7—34.1℃,高溫(水溫高于30℃)和低溫(水溫低于15℃)時間較長,接近6個月,而這段時間大口黑鱸攝食量顯著降低或停止,可見華中地區接近6個月的高溫與低溫期并不利于大口黑鱸的生長。周年光照時長在10.2—14.1 h/d間,雌魚GSI周年變化在0.63%—7.95%。
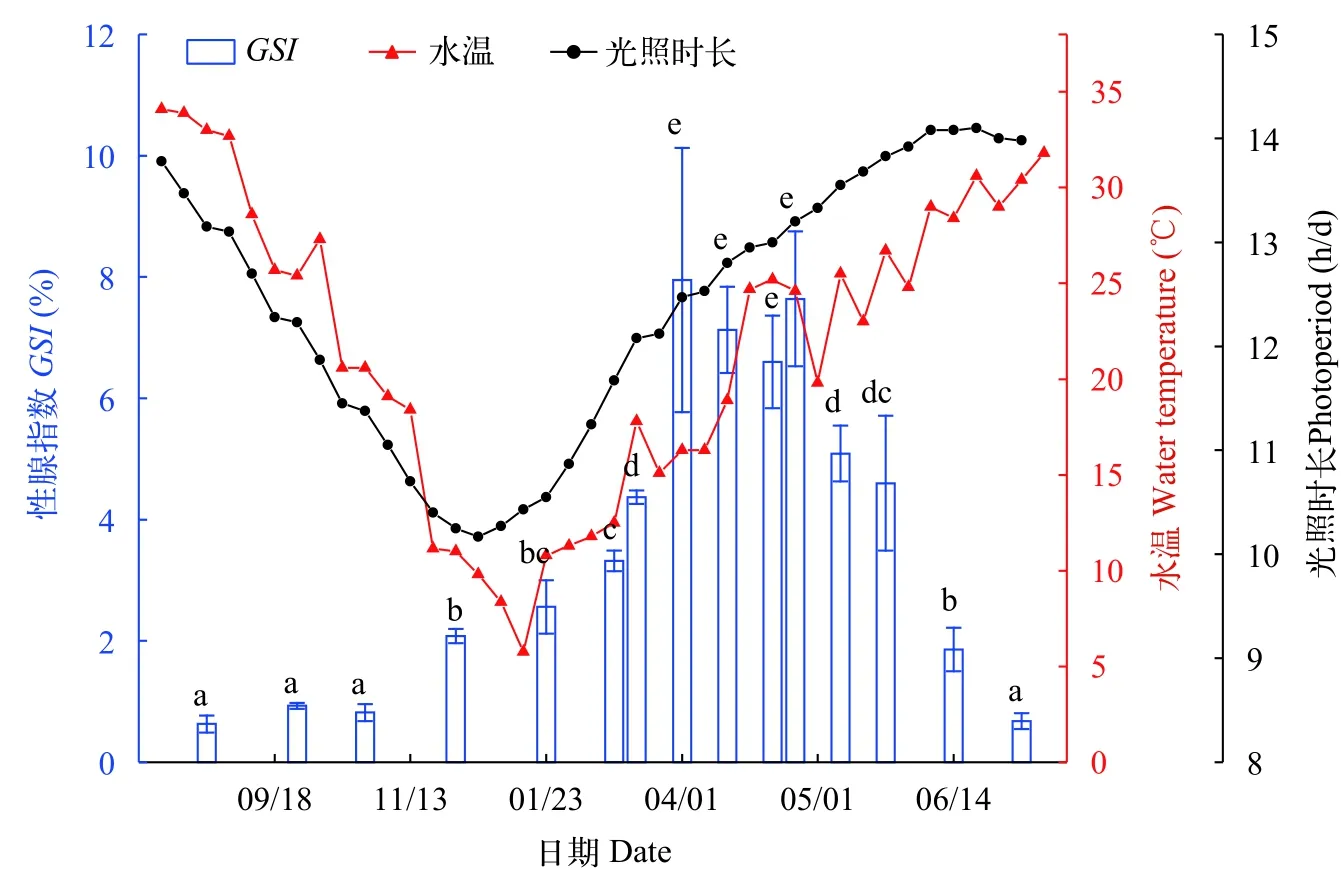
圖2 大口黑鱸卵巢性腺指數(GSI)的周年變化Fig.2 Changes annual gonadosomatic index in largemouth bass不同字母表示差異顯著(P<0.05)
繁殖前期:當水溫回升至15.1℃后,卵巢進入快速發育階段,卵母細胞大量積累卵黃,從3月19日至4月19日,GSI由4.37%增長至最大值7.95%,差異顯著(P<0.05)。此階段以Ⅳ期卵巢為主,呈米黃色,幾乎充滿整個腹腔(圖3B),組織學觀察可見卵母細胞內充滿卵黃顆粒,其間布滿油滴,有的進一步融合成體積較大的油球(圖4B)。Ⅳ時相卵母細胞數量比由15%增至31%,Ⅴ時相卵母細胞數量比由8%增至37%(圖5)。
繁殖期:實驗觀察到大口黑鱸繁殖期從4月19日開始,5月23日產卵活動結束,此時卵巢顏色變深,卵粒透明(圖3C),卵黃顆粒水合作用顯著,大部卵黃顆粒融合,油滴聚合,油球變大,組織切片觀察還可看見部分空濾泡膜存在(圖4C)。GSI由7.64%降至4.60%,前后差異顯著(P<0.05),值得注意的是,繁殖結束時GSI仍然較大,卵巢中仍有大量成熟卵子未能產出,Ⅴ時相卵母細胞數量比占15%,Ⅳ時相卵母細胞數量比占13%(圖5)。繁殖期水溫在19.8—26.7℃波動。

圖3 大口黑鱸卵巢解剖圖Fig.3 Ovary anatomy of largemouth bass
退化階段:當水溫高于26.7℃后,產卵活動結束,卵泡閉鎖,大約歷時50d左右卵巢退化回Ⅱ期(圖3E),GSI由4.60%降至0.68%,差異顯著(P<0.05),此階段卵巢中以Ⅱ時相、Ⅵ時相卵母細胞為主,其中Ⅵ時相卵母細胞無規則形狀,為退化崩解的Ⅴ時相、Ⅳ時相卵母細胞(圖4D),卵巢外觀松軟,呈肉紅色(圖3D)。此后直至10月中旬,卵巢一直處于Ⅱ期(圖3E、圖4E),且GSI差異不顯著(P>0.05)。
啟動發育階段:華中地區卵巢啟動發育的時間是在10月中旬至12月初,GSI由0.82%增長到2.08%,開始出現顯著差異(P<0.05)。卵巢由Ⅱ期發育至Ⅲ期(圖3F和圖4F),開始沉積卵黃,出現放射帶,濾泡膜細胞逐漸變為兩層,卵巢中Ⅲ時相卵母細胞數量比升至34%。此間水溫逐漸降低,由20.6℃降至11.0℃,光照時長由11.5 h/d降至10.3 h/d,接近一年最低值(圖2)。
越冬期:大口黑鱸以Ⅲ期卵巢越冬,越冬階段卵巢發育緩慢,在12月10日時GSI為2.08%,3月6日時僅為3.32%,前后存在顯著差異(P<0.05),但歷時較長,相對發育較慢(圖3F和圖4A)。此期間Ⅲ時相卵母細胞在29%—34%。越冬期溫度在5.76—13.5℃之間波動。
2.2 大口黑鱸MFI、HSI變化規律
池塘養殖大口黑鱸雌魚HSI的周年變化范圍為1.47%—3.05%,12月份達到最大,HSI全年最低值出現在4月中旬。HSI與GSI相關性分析發現,兩者在10月至次年4月產卵前關系緊密,呈顯著負相關(P<0.05,r=-0.531),在5—9月,兩者不存在顯著相關性(P>0.05,圖6A)。
池塘養殖大口黑鱸雌魚的MFI周年變化范圍在0.64%—2.09%,MFI最大值出現在10月份,表明此時腸系膜脂肪積累量達到最大,這為越冬儲存了足夠能量,而5月底MFI為一年最低值,也正是繁殖活動結束的時間。將MFI與GSI進行相關性分析發現,在10月至次年4月份兩者顯著相關(P<0.05),呈負相關關系(r=-0.681);在5—9月,MFI和GSI也同樣存在顯著性(P<0.05)負相關(r=-0.683,圖6B)。
2.3 自然繁殖季節大口黑鱸繁殖力的統計
由表1可知,2齡大口黑鱸性腺發育到V期時,絕對繁殖力為44637—69853粒,平均值為54127.80粒;相對體重繁殖力在54.35—66.25粒/g,平均值為59.12粒/g;相對體長繁殖力變化幅度較大,在137.20—192.00粒/mm,平均值為160.68粒/mm。絕對繁殖力在45000—65000粒的個體占80%(圖7C)。經相關性分析發現,大口黑鱸絕對繁殖力與體質量(r=0.879)、性腺重(r=0.819)、體長(r=0.704)顯著(P<0.05)正相關,而與GSI相關性不顯著(P>0.05)。其中,絕對繁殖力與體質量的線性回歸方程為Y=62.12X-2532.15(R2=0.71,圖7A),絕對繁殖力與體長的線性回歸方程為Y=238.39X-25801.62(R2=0.45,圖7B)。

圖4 大口黑鱸卵巢各發育時期的組織學觀察Fig.4 Histological observation of ovarian development in largemouth bass
在整個繁殖季節共兩個產卵期,中間間隔7天,第一次共連續收集產卵網片356片,第二次182片,隨著水溫升高,產卵網片數逐漸減少,當水溫高于26.7℃后,產卵停止(圖7D)。
2.4 大口黑鱸反季節繁殖
實驗開始前大口黑鱸卵巢GSI為0.53%(圖8A),處于Ⅱ期(圖9E和圖10E),精巢GSI為0.21%(圖8B),同樣處于Ⅱ期(圖9A),精巢切片可以發現僅有少量小葉腔內可見少量精子(圖10A)。
在降溫處理結束后(9月29日),控溫組雌魚GSI增至0.83%,對照組增至0.79%,促熟組增至0.67%,三組間差異不顯著(P>0.05,圖8A)。雄魚GSI三組間差異也不顯著(P>0.05,圖8B)。
與雌魚相比,雄魚精巢發育較早,成熟也較快(圖8C和圖8D),在10月8日便發現,控溫組通過擠壓雄魚腹部可見乳白色精液從生殖孔流出,將精液在顯微鏡下用淡水激活,精子運動劇烈。此時雄魚GSI為0.58%,對照組GSI為0.36%,促熟組GSI為0.34%,控溫組和對照組存在顯著差異(P<0.05,圖8B),而促熟組和對照組差異不顯著(P>0.05,圖8B)。

圖5 不同采樣時間點大口黑鱸卵巢各時相卵母細胞的數量比Fig.5 The number ratio of oocytes in the ovary of largemouth bass at different sampling time points
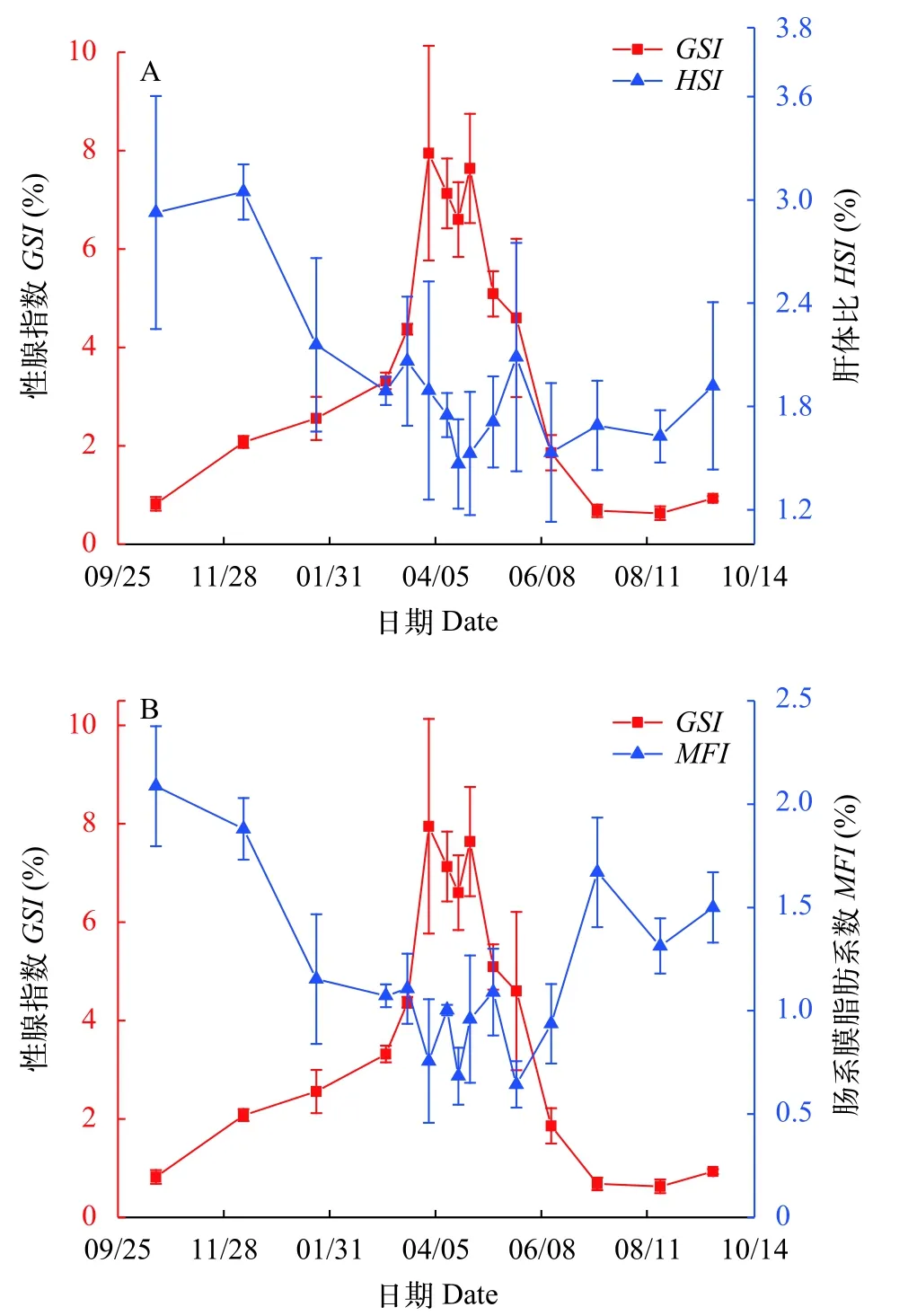
圖6 大口黑鱸HSI、MFI和GSI間的關系Fig.6 Relationship between HSI,MFI and GSI in largemouth bass
在10月29日時,此時控溫組和對照組雌魚GSI開始存在顯著差異(P<0.05,圖8A),控溫組GSI增至1.97%,對照組GSI為1.36%,而促熟組和對照組差異仍不顯著(P>0.05,圖8A)。
在11月26日結束時,控溫組和對照組雌魚GSI存在顯著差異(P<0.05,圖8A),促熟組和對照組差異不顯著(P>0.05,圖8A),此時控溫組卵巢發育至Ⅳ期后期(圖9F和圖10F),對照組卵巢發育至Ⅲ期(圖9G和圖10G),促熟組卵巢發育至Ⅲ期后期(圖9H和圖10H)。控溫組和對照組雄魚GSI存在顯著差異(P<0.05,圖8B),但兩組精巢均發育至Ⅳ期末,小葉腔內充滿大量成熟精子(圖9B、圖9C、圖10B和圖10C),促熟組和對照組在11月15日時差異不顯著(P>0.05),但通過組織切片觀察促熟組精巢才發育至Ⅲ期,以初級精母細胞為主(圖10D)。
遺憾的是此后氣溫和水溫驟降,地下水難以維持適宜大口黑鱸性腺發育產卵的水溫,實驗被迫結束,但以上結果仍表明本實驗方法有效促進了大口黑鱸性腺啟動發育。
3 討論
3.1 大口黑鱸卵巢隨水溫、光照時長變化的周年發育規律
魚類的性腺發育主要是指精卵的增殖、生長和成熟的過程,而溫度是調控這一過程的關鍵,通常認為,溫度是通過啟動下丘腦-垂體-性腺軸的生理功能來發揮作用[18,19]。有學者報道,在自然水域中,受熱源污染地區的大口黑鱸幼魚性腺發育成熟要早于非熱源污染區[20],同樣在文昌魚(Branchiostoma belcher)[10]、牙鲆(Paralichthys olivaceus)[9]中,也表明升溫能夠促進性腺發育成熟。當然,也可通過控制低溫,推遲性腺發育,在對繁殖前期大口黑鱸進行低溫處理,水溫平均16℃,能使親魚產卵時間推遲到6—9月份,而對照組(平均水溫23℃)自然情況下5月初便開始產卵[21]。因此,研究溫度和性腺發育的關系將有助于我們實現人工調控性腺發育周期的目的,這也是我們通過控溫手段來實現大口黑鱸反季節繁殖的基礎。
溫度影響性腺的發育成熟。Martyniuk等[23]對德克薩斯州野生大口黑鱸性腺發育研究中指出,卵子的增殖過程和小生長期是在6—9月,這期間主要進行卵母細胞數量的增加和濾泡膜的初步形成,但GSI并無顯著差異,在10—12月,初級卵母細胞進入大生長期,積累卵黃,體積增大,GSI與前幾個月表現出顯著差異(P<0.05),這與我們研究的池塘大口黑鱸卵子增殖及生長的時間大致相似,在7—10月,Ⅱ時相卵母細胞為主,濾泡膜逐漸形成,到12月時卵母細胞已進入Ⅲ時相,出現兩層濾泡膜、出現放射膜,細胞質中開始沉積卵黃。
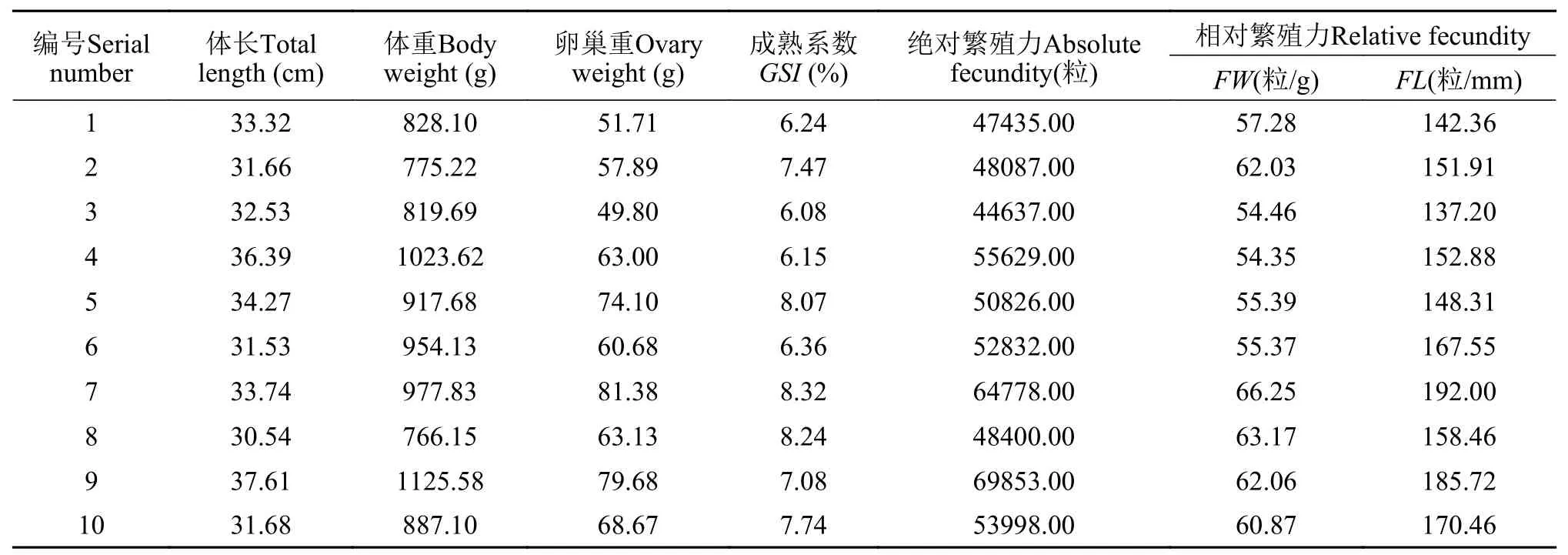
表1 池塘養殖大口黑鱸雌魚的繁殖力Tab.1 Fecundity of pond-raised largemouth bass(n=10)

圖7 池塘養殖大口黑鱸相對繁殖力的統計Fig.7 Relative fecundity statistics of pond-raised largemouth bass
對比兩地水溫可以發現,均是在降溫的過程中性腺開始啟動發育,在Martyniuk等[23]研究的野生大口黑鱸中,水溫從24.4℃降至16.1℃時,性腺啟動發育,GSI前后出現顯著差異;我們研究的池塘養殖大口黑鱸的水溫由34.1℃降至20.6℃時,卵巢中仍以第Ⅰ、Ⅱ時相卵母細胞為主,但水溫由20.6℃降至11.0℃時,卵巢中逐漸以Ⅲ時相卵子為主,開始積累卵黃,同時GSI發生顯著變化;而在Brown等[24]的研究中同樣發現,在美國中西部地區,當水溫由19.4℃降至13.9℃時,大口黑鱸卵巢中開始出現Ⅲ時相卵子,表明性腺開始發育,可見在不同環境條件下,大口黑鱸性腺開始發育的水溫不盡相同,但均需要經歷降溫刺激的過程。
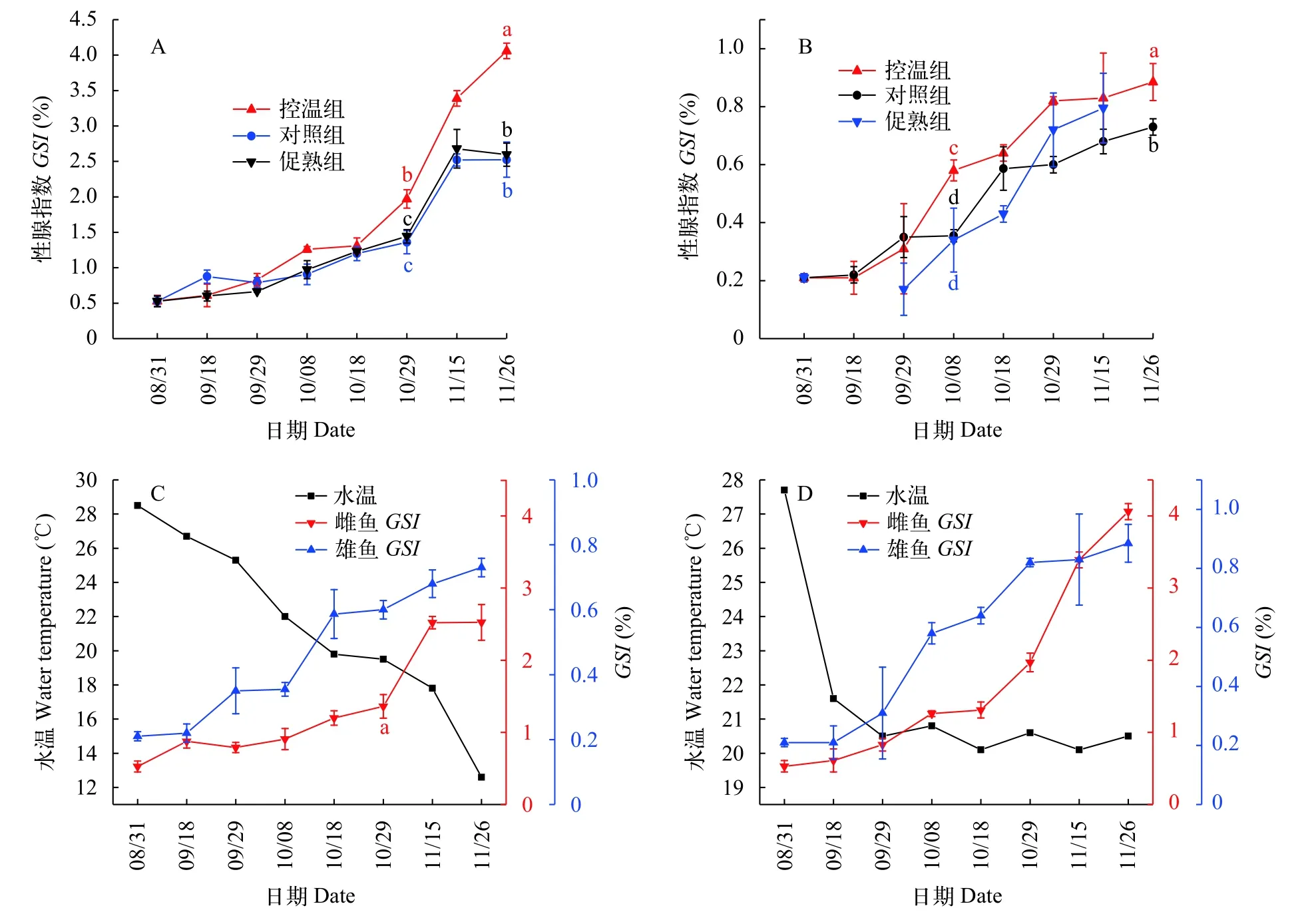
圖8 反季節大口黑鱸培育中性腺指數(GSI)變化規律Fig.8 Gonadosomatic index(GSI)changes of largemouth bass in off-season spawning
溫度同樣影響魚類的產卵退化。當溫度高于或低于繁殖水溫時,魚類性腺便開始退化,如在成熟的雌性鱘魚養殖中,水溫高于15℃時,引起了卵巢卵泡閉鎖、胚泡遷移和排卵受阻等現象[22]。Gross等[15]曾報道過佛羅里達州池塘養殖大口黑鱸的繁殖期是在1月中旬至6月中旬,長達5個月的繁殖期,而在本研究中,華中地區大口黑鱸的繁殖期是4月中旬至5月底,僅僅1個多月。兩地區大口黑鱸的繁殖水溫相似,佛羅里達州繁殖期水溫在18.3—28.3℃,華中地區繁殖期水溫在19.2—26.7℃,但佛羅里達州的氣候條件使得池塘水溫相對穩定,保證了長時間適宜的繁殖水溫,而華中地區氣溫變化較大,冬季氣溫低,夏季氣溫高,這使得適宜繁殖的時間較短。我國南北氣候差異較大,華南地區冬無嚴寒,夏無酷暑,全年平均氣溫25℃左右,使得自然環境下大口黑鱸2月便進入繁殖期,5月初結束產卵活動,同樣與華中地區存在很大差異[4]。Brown等[24]也曾報道,美國中西部地區大口黑鱸的繁殖期在6月中旬至7月底,繁殖水溫在16.6—18.5℃,這與華中地區、華南地區及佛羅里達州地區均存在差異,這一切表明大口黑鱸適應能力極強,在不同環境下已經演變出了適應當地環境的性腺發育規律與繁殖規律,因此開展相關研究,應基于本地區氣候條件進行。
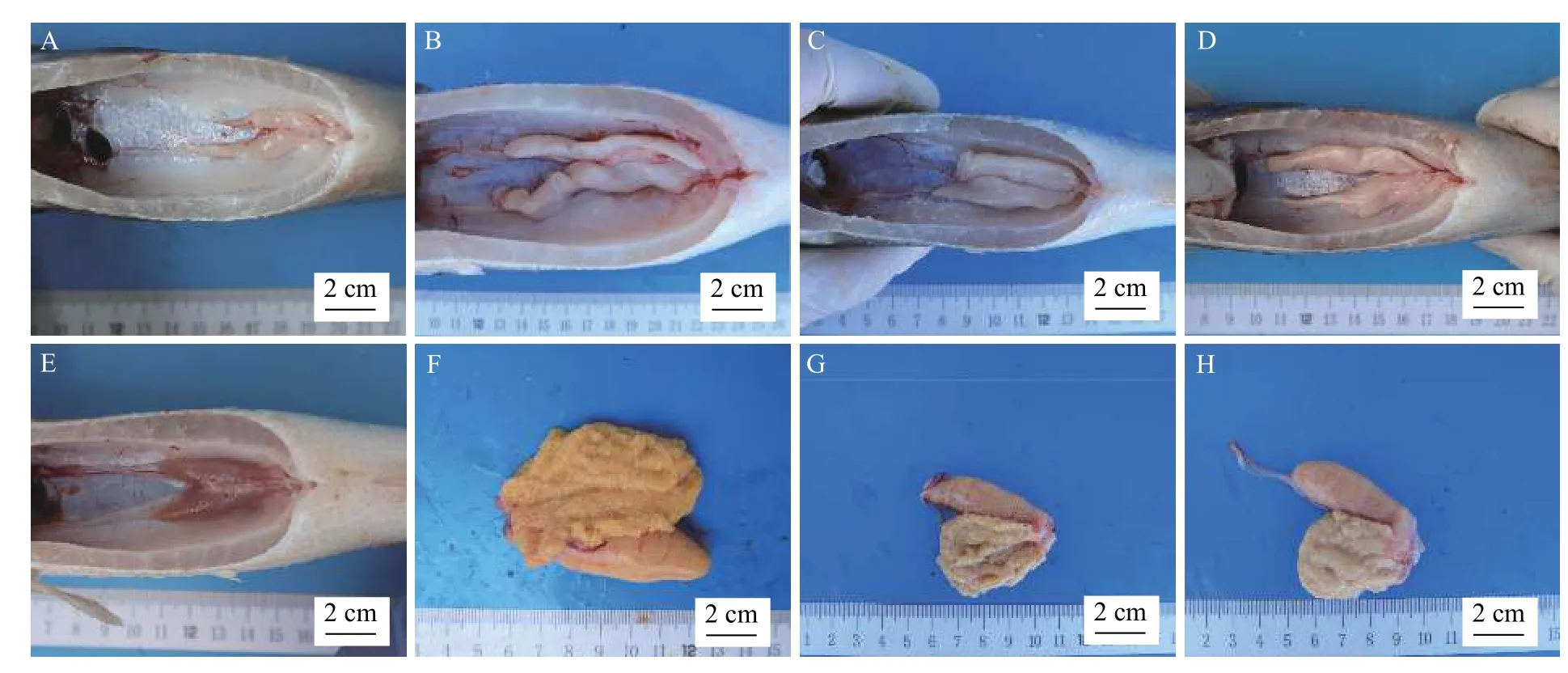
圖9 大口黑鱸反季節繁殖中各組性腺解剖圖的比較Fig.9 Comparison of gonad development in different groups during the off-season spawning in largemouth bass
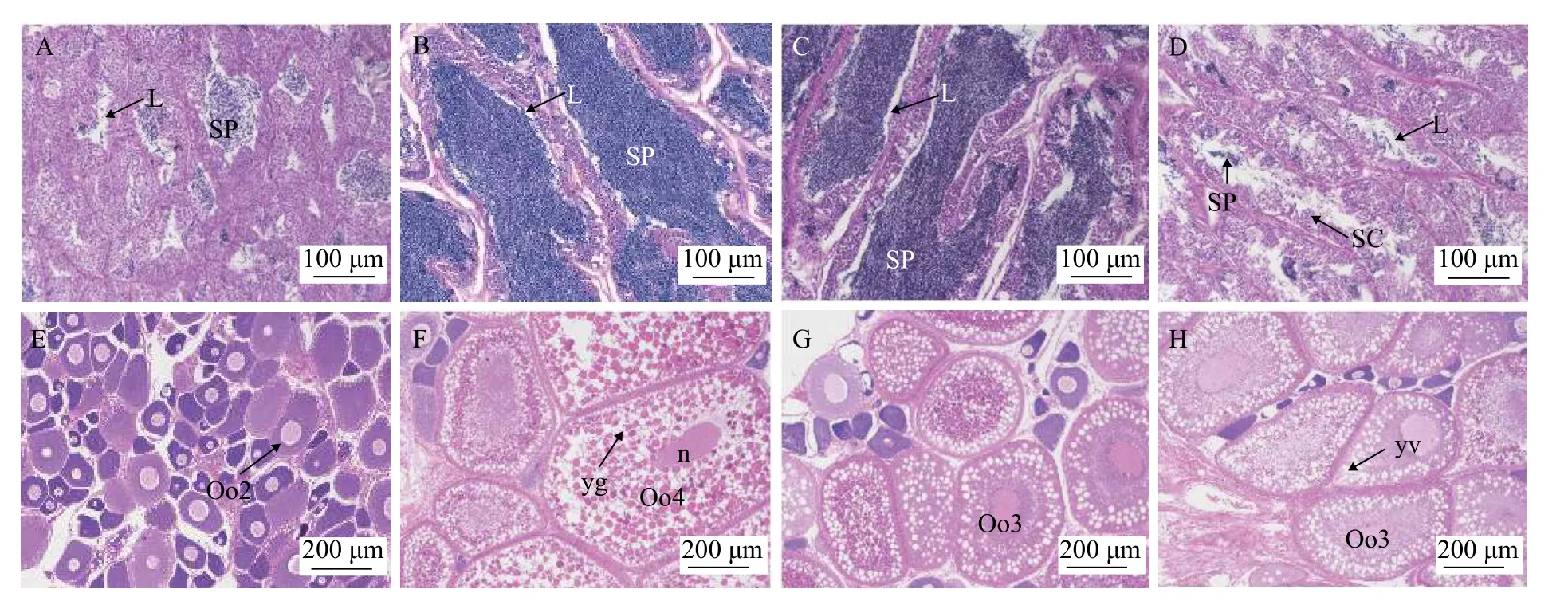
圖10 大口黑鱸反季節繁殖中性腺組織切片觀察Fig.10 Gonadal histological observation of largemouth bass in off-season spawning
華中地區高溫較早來臨使大口黑鱸的繁殖效率極低,在繁殖期結束時,雌魚GSI仍有4.6%,而佛羅里達州地區產卵結束時雌魚GSI僅有2.3%[15]。華中地區在5月24日繁殖結束后雌魚卵巢中仍有大量成熟待產的卵(13%的Ⅳ時相卵母細胞、15%的Ⅴ時相卵母細胞),但水溫過高,卵泡閉鎖,性腺開始退化,我們推測這是造成華中地區苗種生產不足問題的一個重要原因。因此,如果有條件保持適宜的繁殖水溫,我們相信對于分批產卵的大口黑鱸而言,在本地區同樣可以延長繁殖期。
光照時長同樣影響著魚類性腺發育,Jourdran等[25]研究指出,光照時長增加(12、18和24h/d)顯著抑制了河鱸(Perca fluviatilis)幼魚的性腺發育,尤其是對雄性抑制效果更佳。光周期也會影響著魚類性腺啟動發育,在Migaud等[26]研究中發現,連續的光照(24h/d)會抑制河鱸性腺啟動發育,卵巢啟動發育的占比低于2%,精巢啟動的占比低于1%,而恒定的光周期(16h/d)條件下河鱸性腺會啟動發育,但發育不同步,而對照組(自然光照時長)20%的雌性,40%的雄性性腺均啟動發育。同樣,我們研究發現,大口黑鱸卵巢啟動發育時光照時長從11.4 h/d降至10.3 h/d(圖2),屬于一年中的短光照時長,這在一定程度上表明,大口黑鱸性腺啟動發育同樣需要短光照時長的刺激。先前團隊在大口黑鱸規模化反季節繁育中就推測,光照可能對于卵子的早期發育更重要,而溫度對于卵黃生成和最終成熟更加重要[6]。在今后大口黑鱸繁育的科研與生產實踐中,為了提升繁育效果,也應當考慮光照時長的控制。
3.2 HSI和MFI隨性腺發育的變化規律
魚類的性腺發育過程十分復雜,目前并未清楚地闡明性腺發育和個體能量轉換間的關系,魚類的能量主要以中性脂肪的形式儲存于肝臟、腸系膜和肌肉中[27,28],為了滿足性腺發育需要,脂肪中的能量物質需要不斷轉向性腺,尤其在卵黃積累階段,肝細胞更是直接合成卵黃蛋白原供給性腺發育[29]。有研究表明,大口黑鱸肌肉中總脂肪含量的季節變化范圍很小,僅為濕重的0.7%—0.9%,表明在性腺發育過程中,肌肉中脂肪供能相對較少,主要以肝臟和腸系膜脂肪供能為主[30]。
在我們研究中,自卵巢啟動發育至繁殖產卵前,GSI分別與HSI、MFI呈顯著(P<0.05)負相關,其中越冬期HSI、MFI下降較快,除了維持正常生理活動消耗外,主要是轉移至卵子內積累卵黃顆粒和油滴,組織切片可以發現越冬前多數卵母細胞內并未沉積卵黃顆粒,越冬后大量卵黃顆粒由卵膜側向核膜側沉積(圖4F和圖4A),后續發育主要是卵子積累營養物質的過程,將消耗大量營養物質,作為分批產卵的魚類,HSI和MFI和外界營養供給將直接影響著性腺的發育質量,在自然水域環境下,由于食物相對匱乏,大口黑鱸GSI一般小于4%[23],而在人工養殖條件下GSI最大可達14%[15];同樣在Brown等[31]研究結果中也表明,MFI在繁殖前降到最低值。因此我們認為,隨著性腺發育,肝臟和腸系膜脂肪中的儲存能量處于不斷消耗的狀態,且主要流向性腺。在性腺退化后,直至性腺啟動發育前,HSI和MFI均不斷增加,這段時間主要是肝臟、腸系膜脂肪重新積累能量的過程,這種高效的能量分配策略保障了性腺的充分發育。因此,性腺發育啟動之前,肝臟和內臟中能量的積累,對于后期繁育效果至關重要。同時,肝臟和內臟中的能量,何時、如何轉移至性腺中,不論對于正常季節還是反季節繁育都非常關鍵。
對比反季節親本與正常季節親本的GSI后發現,在性腺同樣發育至Ⅳ末時,反季節親本成熟系數僅有4.06%,而正常季節已達到7.13%,團隊在大口黑鱸反季節親本培育實驗中,產卵前1個月時GSI僅為2.50%[6],可見性腺接近成熟時三者GSI差別較大。對比性腺啟動發育時三者脂肪沉積情況發現,正常季節親魚的MFI為2.09%,達到了一年中的最大值,在本研究中反季節親本MFI僅有1.34%,團隊先前研究的親本MFI僅1.1%[6],三組MFI存在差異主要是受攝食情況影響,華中地區正常季節10—12月間水溫在20℃左右,非常適宜大口黑鱸攝食生長,脂肪積累量達到一年最大值,而反季節培育前本地區屬于高溫(>30℃)期,大口黑鱸幾乎處于停止攝食的狀態,缺少了營養強化的過程,不利于脂肪沉積。因此我們推斷導致繁殖前大口黑鱸卵巢GSI差異較大可能與親魚攝食情況、脂肪積累量有關,同時比較反季節實驗中控溫組和對照組攝食情況可以發現,對照組共投喂15.5 kg配合飼料,控溫組由于水溫適宜,共投喂25.3 kg配合飼料,控溫組平均每尾魚的飼料消耗量大于對照組,但試驗結束時對照組MFI為2.12%,控溫組雌魚MFI僅為1.53%,控溫組MFI卻顯著低于對照組,再次表明性腺發育消耗了大量的腸系膜脂肪。由于大口黑鱸卵巢屬于不同步發育類型,充足的能量供給將能夠保證更多的卵子得到發育,從而保證了后期性腺的發育質量,不過這仍需后續實驗驗證。但我們必須注意到脂肪積累對性腺發育質量的重要性,尤其是在反季節親本培育中,為保證性腺的發育質量,前期必須進行營養強化,在一定脂肪積累量的基礎上再進行培育。
3.3 大口黑鱸的反季節繁殖
目前,華中地區大口黑鱸所面臨的問題已經嚴重限制了該產業的發展,解決這些問題的有效方法之一便是培育反季節苗種,但目前并沒有比較高效的親本培育方法。團隊之前利用水庫深層水進行控溫處理,模擬正常繁殖季節環境的變化規律(秋季的降溫過程、越冬時的低溫階段及春季的升溫過程),并按照一定比例縮減在140d內完成,最終在9月和10月成功獲得苗種[6],該模式對基礎設施要求較高,需要在夏季高溫時仍有充足的低溫水(10℃左右)供應,同時還要有充足的高溫水用于后續升溫培育,為了進一步控制成本,提高效率,程序性溫度控制至關重要,需要進一步摸索低溫限度、低溫時長等關鍵因素。Matthews等[32]采用的是大型加熱-制冷機組控溫,模擬自然環境下光照、溫度變化規律,依次先降溫,再維持低溫(10℃左右),最后升溫至產卵水溫,并在此過程中調控光照時長,由10 h/d降至8 h/d,后升至14 h/d,最終歷時130d成功孵化出苗種,但是該方法成本投入太高,且規模較小,不適合生產應用。可以發現,這兩種方法有一個共同的限制因素,即均需要在夏秋高溫季節時對親本進行低溫刺激(10℃左右),以達到性腺啟動發育的目的,因此,我們考慮能否充分利用華中地區的地下水資源,采用更高的水溫來刺激親本,以實現親本性腺啟動發育。基于我們已有的井水資源,探究了(20±1)℃的水溫刺激親本,結果同樣達到性腺啟動發育的目的。
Spengler等[33]也表示持續低溫(≤12℃)刺激是反季節大口黑鱸性腺啟動發育的關鍵,但在我們實驗中發現,短暫降溫后持續采用(20±1)℃水溫刺激,同樣實現了性腺啟動發育。還有一點值得注意,在團隊前期研究中,大口黑鱸卵巢由Ⅱ期增至Ⅲ期是在水溫回升至20℃之后,表明在前期的降溫、維持低溫及升溫至20℃前,卵巢始終處于Ⅱ期,并沒有發生顯著性差異(P>0.05)[6]。因此,可以初步推斷,大口黑鱸性腺啟動發育并不一定需要模擬越冬環境或低于某個溫度閾值的水溫刺激,在經過某個溫度差的降溫[本實驗由(29±1)℃降至(20±1)℃]后,保證降溫之后有適宜性腺生長發育的水溫[本實驗(20±1)℃]同樣能達到性腺啟動發育的目的,不過其他的降溫差及降溫后培育水溫能否啟動性腺發育仍需要后續實驗來驗證。
華中地區大口黑鱸反季節繁殖的最佳時間段是在9—10月份,一是孵化后池塘水溫仍然適合苗種生長,能保證以大規格苗種越冬;二是此時池塘中仍然可以培育出豐富的浮游生物供仔魚早期生長[6];三是此階段培育出來的苗種,在來年繁育季節并未達到性成熟,可以極大減少因性成熟所帶來的能量消耗(掉膘)和一定比例的死亡。比較團隊先前研究[6]和Matthews等[32]的實驗可以發現,整個反季節培育時長在4—5個月,而正常季節大口黑鱸性腺退化回II期是在6—7月,因此,反季節大口黑鱸培育的最佳起始時間是在7月初。由于我們培育實驗是在8月31日開始,后期氣溫驟降,導致最終未能順利完成產卵。在我們實驗中,在省去低溫(10℃左右)刺激后有效縮短培育周期,僅通過3個月處理,卵巢即發育至Ⅳ期末,出現核偏移現象,達到了生理成熟,表明該方法能適當縮短培育時間,提高效率。
本實驗通過對華中地區大口黑鱸卵巢隨水溫和光照時長變化的周年發育規律進行研究,并探索華中地區反季節繁殖的方法,我們發現水溫升高對于大口黑鱸卵黃沉積和卵子最終成熟非常重要,適宜繁殖水溫的維持對于延長華中地區產卵時間至關重要;對于反季節繁殖來說,溫度的降低或升高(溫度變化),而不是低溫的維持,可能更加重要。本研究對后續研究大口黑鱸在全世界范圍內迅速適應當地氣候環境、反季節繁育和種群控制等方面有指導意義。
