典型化學(xué)加工條件對(duì)不溶性蠶蛹蛋白凝膠特性影響
2021-01-29 09:35:44黎重陽謝盛莉馬良侯勇付余張宇昊
食品與發(fā)酵工業(yè) 2021年2期
關(guān)鍵詞:體系
黎重陽,謝盛莉,馬良,,侯勇,付余,張宇昊,*
1(西南大學(xué) 食品科學(xué)學(xué)院,重慶, 400715)2(西南大學(xué) 前沿交叉學(xué)科研究院,生物學(xué)研究中心,重慶, 400715)
蠶蛹(silkworm pupa,SP)是蠶蛾科家蠶蛾的蛹,屬于蠶繭抽絲后的副產(chǎn)物,其于1998年被衛(wèi)生部批準(zhǔn)為“可作為普通食品管理的新資源食品”,是第一個(gè)被認(rèn)定的昆蟲類食品。我國作為養(yǎng)蠶大國,近年來全國年產(chǎn)蠶蛹量在30萬t以上,約占全世界總產(chǎn)量的80%[1]。蠶蛹蛋白在蠶蛹中占比極高,幾乎是蠶蛹干基中的主要營養(yǎng)物質(zhì),因此大多研究以蠶蛹蛋白為主體研究對(duì)象。呂汶駿等[2]研究發(fā)現(xiàn)蠶蛹蛋白中8種必需氨基酸含量占比為49.09%,必需氨基酸與非必需氨基酸之比為96.43%,高于WHO/FAO提出的參考蛋白模式(8種必需氨基酸含量占比為40%,必需氨基酸與非必需氨基酸之比為60%);由此可見,蠶蛹蛋白是一種高營養(yǎng)價(jià)值的蛋白資源,但由于其含有潛在過敏原,如30K家族蛋白、幾丁質(zhì)酶、卵黃原蛋白、丙糖磷酸異構(gòu)酶等[3],部分人食用蠶蛹相關(guān)產(chǎn)品后會(huì)出現(xiàn)頭暈、惡心、嘔吐等過敏反應(yīng),從而限制了蠶蛹在實(shí)際生產(chǎn)過程中的應(yīng)用。因此,在食品安全的前提下充分開發(fā)利用蠶蛹資源,必須解決蠶蛹蛋白過敏問題。謝盛莉等[4]按照溶解性對(duì)蠶蛹蛋白進(jìn)行分級(jí)提取,包括水溶性、醇溶性、鹽溶性、堿溶性蛋白和不溶性蛋白,并檢測(cè)其中存在的潛在過敏原,結(jié)果發(fā)現(xiàn)蠶蛹蛋白中不溶性蛋白含量高達(dá)53%,在不同種類蠶蛹蛋白中所占比例最高,且過敏原分布顯示,其中僅檢測(cè)出3種過敏原,僅占不溶性蛋白9.87%,含量較低,且經(jīng)過多次堿洗處理后理論上可以降低其致敏性,使不溶性蠶蛹蛋白有直接開發(fā)為蛋白產(chǎn)品或者作為改善食品功能性質(zhì)的添加物的可能。
蛋白質(zhì)的功能特性在食品原料的加工利用或儲(chǔ)存過程中起著極其重要的作用,穆利霞等[5]研究表明繅絲后蠶蛹蛋白的乳化活性(25.52 m2/g)和溶解性(氮溶解指數(shù)60.17%)最好;練釗等[6]使用Osborn法分級(jí)提取繅絲后的蠶蛹水溶性、鹽溶性、醇溶性以及堿溶性蛋白,并對(duì)其功能特性進(jìn)行測(cè)定,結(jié)果表明水溶性蛋白的溶解性和保水性最強(qiáng),鹽溶性蛋白具有良好的持油性,醇溶性蛋白具有良好的乳化活性。由此可見,蠶蛹蛋白具備一定功能特性,且按照溶解性對(duì)其進(jìn)行分級(jí)提取后可根據(jù)不同功能特性而應(yīng)用于更加廣泛的食品加工領(lǐng)域。
一般動(dòng)物性不溶性蛋白研究較多集中在凝膠特性方面,在食品加工過程中常見的條件如pH、離子強(qiáng)度、添加谷氨酰胺轉(zhuǎn)氨酶(transglutaminase,TG酶)等會(huì)對(duì)蛋白質(zhì)的功能特性產(chǎn)生一定影響,但目前關(guān)于蠶蛹中不溶性蛋白的研究報(bào)道較少,且其凝膠特性尚無報(bào)道。因此,為明確典型食品加工條件對(duì)蠶蛹蛋白凝膠特性的影響規(guī)律,實(shí)驗(yàn)以鮮蠶蛹為原料提取不溶性蛋白,對(duì)其氨基酸組成進(jìn)行測(cè)定,采用不同的pH、離子強(qiáng)度處理蛋白,并添加TG酶,進(jìn)行流變學(xué)以及拉曼光譜學(xué)的分析,以期為進(jìn)一步的蠶蛹蛋白產(chǎn)品開發(fā)提供理論參考。
1 材料與方法
1.1 材料與儀器
鮮蠶蛹(家蠶實(shí)驗(yàn)品種“夏芳”,25 ℃,桑葉飼養(yǎng),化蛹后第3天放置于-20 ℃凍存?zhèn)溆?、無水乙醇(99.9%)、HCl、NaCl、十二烷基硫酸鈉、二甲基硅油。所有試劑均為分析純,所有水均為二次蒸餾水。
JA3003B電子天平,上海精天電子儀器有限公司;DW-3型數(shù)顯電動(dòng)攪拌器,昆山市超聲儀器有限公司;HH-4數(shù)顯恒溫?cái)嚢杷″仯虾P轮Z儀器設(shè)備有限公司;Multifuge X3R高速冷凍離心機(jī),賽默飛世爾科技有限公司;L8900全自動(dòng)氨基酸分析儀,日本日立公司;DHR-1流變儀,美國TA公司;DXR2拉曼光譜儀,賽默飛世爾科技有限公司。
1.2 實(shí)驗(yàn)方法
1.2.1 蠶蛹不溶性蛋白提取
蠶蛹預(yù)處理:蠶蛹解凍,沸水漂洗5 min,料理機(jī)粉碎。70%(體積分?jǐn)?shù))乙醇脫脂,除醇溶蛋白(2 h,2次),10 g/L NaOH除去堿溶性蛋白(2 h,3次),200目濾布過濾除蛹?xì)ぃ瑸V液離心(10 000 r/min,10 min),用純水多次洗滌蛋白至離心液中性,得到不溶性蛋白。
1.2.2 氨基酸分析
參照國標(biāo)GB 5009.124—2016測(cè)定氨基酸。
1.2.3 不同條件處理對(duì)蠶蛹不溶性蛋白流變性能以及結(jié)構(gòu)的影響
1.2.3.1 流變性能測(cè)定
不同的前處理方式:(1)不同pH處理對(duì)蠶蛹不溶性蛋白流變性能的影響(頻率及溫度掃描):取10 g不溶性蛋白分別置于pH 5.0~8.0的溶液中,4 ℃過夜,離心,室溫平衡30 min后進(jìn)行流變測(cè)試。(2)不同鹽濃度對(duì)蠶蛹不溶性蛋白流變性能影響(頻率及溫度掃描):取10 g不溶性蛋白分別加入NaCl,使得鹽質(zhì)量濃度分別為0、10、20、25、35 g/L,攪拌均勻,4 ℃過夜,室溫平衡30 min后進(jìn)行流變測(cè)試。(3)TG酶添加對(duì)蠶蛹不溶性蛋白流變性能的影響(頻率及溫度掃描):取10 g不溶性蛋白,加入5% TG酶40 ℃反應(yīng)2 h,4 ℃過夜,次日取出室溫平衡30 min后進(jìn)行流變測(cè)試。
參照文獻(xiàn)[7]方法測(cè)定蛋白流變性能,并略做修改。測(cè)定指標(biāo)為儲(chǔ)能模量(G′)、損失模量(G″)和tanδ值。頻率掃描參數(shù):溫度25 ℃,應(yīng)變?yōu)?%,頻率范圍為0.01~10 Hz。溫度掃描參數(shù):將樣品從25 ℃開始以2 ℃/min的升溫速率升溫至80 ℃,5 ℃/min的降溫速率降溫至25 ℃。頻率為0.1 Hz,應(yīng)變1%。
1.2.3.2 拉曼光譜分析
不同前處理方式:(1)不同pH前處理:取10 g不溶性蛋白分別置于pH 5.0~8.0的溶液中,4 ℃過夜,離心,室溫平衡30 min后進(jìn)行拉曼光譜測(cè)試。(2)不同鹽濃度前處理:取10 g不溶性蛋白分別加入NaCl,使得鹽質(zhì)量濃度分別為0、10、20、25、35 g/L,攪拌均勻,4 ℃過夜,室溫平衡30 min后進(jìn)行拉曼光譜測(cè)試。使用拉曼光譜儀記錄凝膠樣品400~3 050 cm-1的拉曼光譜。測(cè)量參數(shù):激發(fā)波長785 nm;物鏡20×焦距;光闌50 μm狹縫;曝光時(shí)間10 s;樣品曝光15次。
1.3 數(shù)據(jù)處理
用SPSS 19.0進(jìn)行顯著性分析,使用單因素方差分析(ANOVA)方法分析數(shù)據(jù),當(dāng)P<0.05時(shí)判定組間存在顯著差異,并使用Origin 9.0軟件作圖。拉曼圖譜處理:拉曼圖譜基線校正、歸屬采用OMINIC軟件,歸一化處理以苯丙氨酸作為內(nèi)標(biāo),譜圖的擬合采用Peakfit軟件。
2 結(jié)果與分析
2.1 氨基酸分析
由表1可以得出,不溶性蛋白質(zhì)氨基酸種類齊全,必需氨基酸/總氨基酸(essential amino acids/total amino acids,E/T)達(dá)到40.27%,符合FAO/WHO推薦模式40%,也符合高質(zhì)量的蛋白質(zhì)E/T比值36%;谷物[8]中的限制性氨基酸賴氨酸含量約為2.5~3.7 g/100g,大豆等[9]油料作物蛋白的限制性氨基酸蛋氨酸含量約為5.5 g/100g。因此,不溶性蠶蛹蛋白是一種營養(yǎng)價(jià)值較高的蛋白資源,可作為一種重要的營養(yǎng)強(qiáng)化劑,添加到谷物制品、大豆蛋白制品中,彌補(bǔ)其限制性氨基酸造成的營養(yǎng)缺陷。
蛋白凝膠作用與本身的氨基酸分布有關(guān),蛋白凝膠的形成與疏水性氨基酸有關(guān),氨基酸中的非極性片段之間的疏水相互作用可以讓蛋白之間結(jié)合緊密。蛋白質(zhì)中疏水氨基酸的比率會(huì)影響凝膠的形成,當(dāng)?shù)鞍字械氖杷园被岢^31.5%時(shí),有可能形成不透明的凝結(jié)型凝膠,非極性氨基酸殘基低于31.5%時(shí)的蛋白質(zhì)則形成半透明或透明型凝膠。疏水相互作用對(duì)凝膠形成有重要作用。由表1可得蠶蛹不溶性蛋白中的疏水性氨基酸比例約為40%,理論上在一定條件下有可能形成不透明的凝結(jié)凝膠[10-11]。
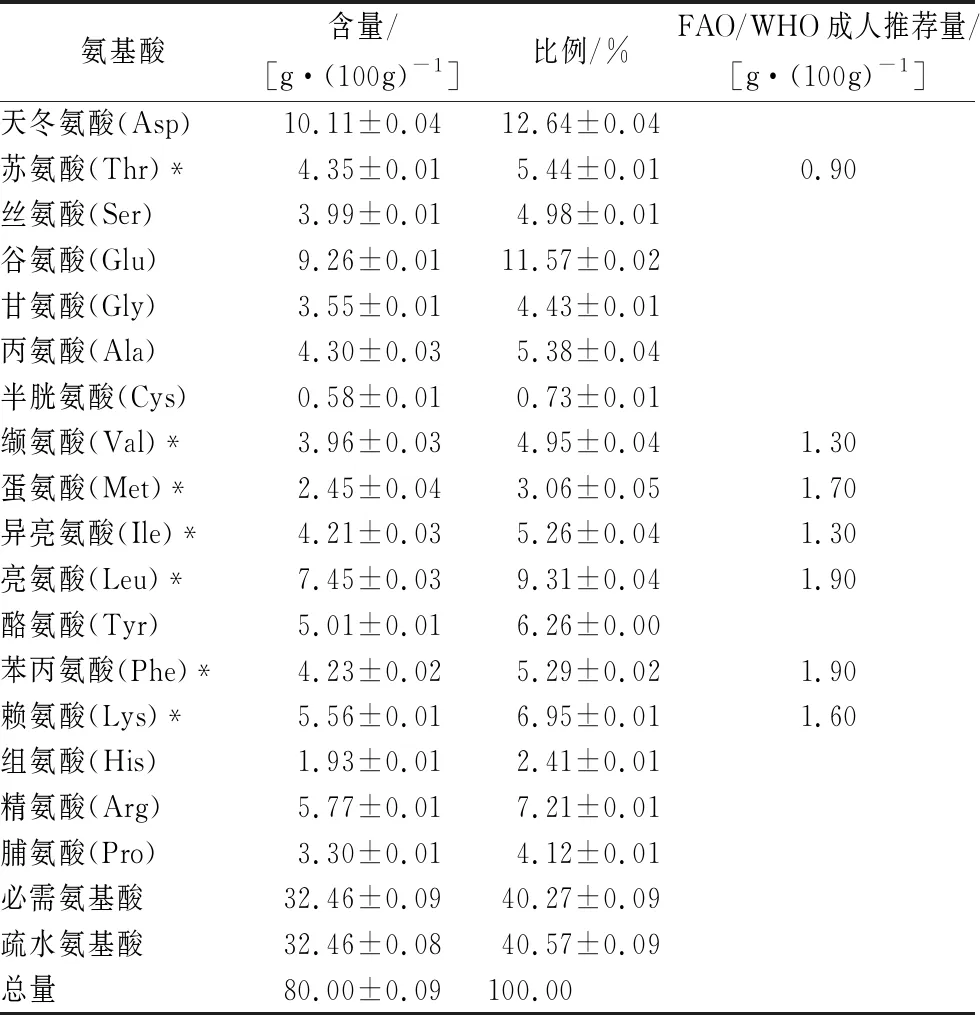
表1 不溶性蛋白氨基酸分析
2.2 不同pH處理對(duì)蠶蛹不溶性蛋白流變性能的影響
儲(chǔ)能模量G′也稱彈性模量,反映蛋白凝膠網(wǎng)絡(luò)結(jié)構(gòu)的形成情況,代表樣品的彈性特征;耗能模量G″也稱黏性模量,代表樣品的黏性特征[12]。損耗角正切tanδ是G″與G′模量的比值,黏性模量(G″)小于彈性模量(G′),tanδ<1,表明凝膠體系中彈性占主導(dǎo)地位[13],反之則表示黏性占主導(dǎo)地位。tanδ值越高則表明樣品組分越黏稠或彈性越低,tanδ值越小,表明樣品彈性越高[14];當(dāng)G′值明顯大于G″值時(shí),tanδ越小,表明該體系屬于弱凝膠體系[15]。
蛋白在加工過程中其凝膠特性受外界條件溫度、pH、離子強(qiáng)度等影響,pH的變化會(huì)引起蛋白分子內(nèi)部結(jié)構(gòu)及分子間作用力發(fā)生變化。食品中體系一般為弱酸性或弱堿性,故選擇pH 5.0~8.0為模擬加工體系,對(duì)蠶蛹蛋白處理后對(duì)其流變性能進(jìn)行測(cè)定。由圖1-a、圖1-b可以看出,在pH 5.0~8.0,隨著頻率的增加,G′和G″模量出現(xiàn)上升趨勢(shì),這表明頻率的增加導(dǎo)致了蛋白分子內(nèi)作用力增強(qiáng)。在相同的振蕩頻率下,pH 6.0時(shí)G′和G″值相對(duì)最高,表明pH 6.0時(shí)蛋白分子間的相互作用較強(qiáng),有利于蛋白凝膠網(wǎng)絡(luò)形成,凝膠性能較好;pH 8.0時(shí)G′和G″值相對(duì)最低,表明此條件下蛋白相互作用相對(duì)較弱,凝膠性能較差。與WESTPHALEN等[16]研究結(jié)果類似,隨著pH值的增加豬肉肌球蛋白的G′值逐漸減小,其原因可能是隨著pH的變化,蛋白質(zhì)分子間的作用力發(fā)生了變化,導(dǎo)致凝膠性能發(fā)生變化。圖1-c顯示,隨著頻率的增加,tanδ值出現(xiàn)下降趨勢(shì),這表明蛋白凝膠體系彈性進(jìn)一步增強(qiáng),tanδ值的變化與蛋白鍵間的弛豫行為有關(guān)[17],在頻率0~1 Hz tanδ出現(xiàn)極大值可能是與初始凝膠網(wǎng)絡(luò)結(jié)構(gòu)的部分松散[10]以及蛋白鍵持續(xù)作用時(shí)間相對(duì)較短有關(guān)[18]。在1~10 Hz頻率范圍內(nèi),pH 8.0時(shí)tanδ最小,此時(shí)體系彈性最好,但在該掃描模式下蛋白樣品的tanδ<1,蛋白的彈性模量(G′)明顯大于黏性模量(G″),表明蠶蛹不溶性蛋白凝膠體系在pH 5.0~8.0是一種弱凝膠。
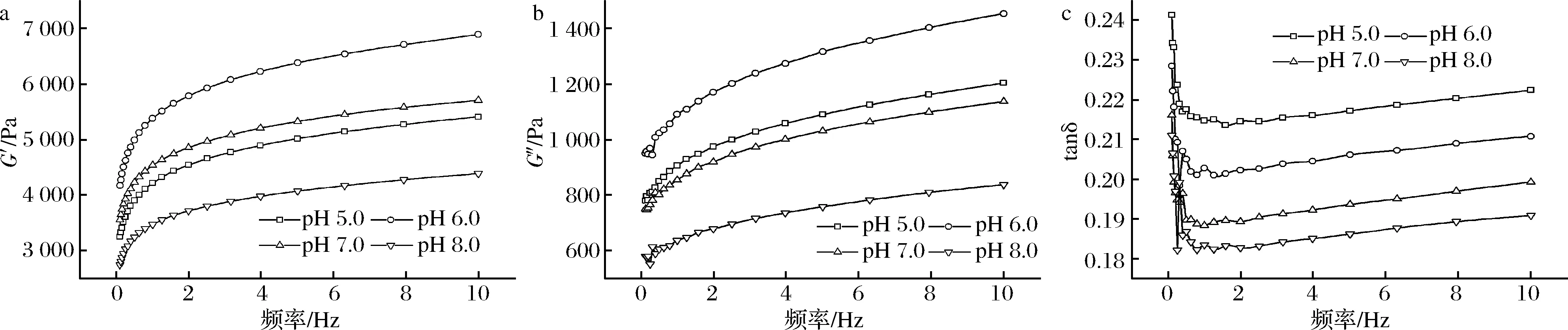
a-G′;b-G″;c-tanδ
由圖2-a、圖2-b可以看出,隨著溫度的升高,蛋白質(zhì)的G′和G″值總體呈現(xiàn)下降趨勢(shì)。溫度升高導(dǎo)致蛋白模量急劇下降,可能是由以下原因引起的[19]:(1)隨著溫度的升高,部分蛋白分子間作用受到破壞,凝膠網(wǎng)狀結(jié)構(gòu)缺少支撐而變得松散;(2)蛋白質(zhì)的構(gòu)象轉(zhuǎn)變可能使得凝膠網(wǎng)絡(luò)的完整性受到破壞,導(dǎo)致蛋白的黏彈性下降。而出現(xiàn)平臺(tái)期的原因可能是高溫下的蛋白凝膠之間的作用力達(dá)到了短暫的平衡。tanδ值下降,說明總體而言高溫下更有利于體現(xiàn)蠶蛹蛋白的彈性,原因是高溫下構(gòu)象轉(zhuǎn)變,蛋白分子黏性受到的破壞更強(qiáng)。tanδ值出現(xiàn)快速下降趨勢(shì),表明隨著溫度升高固體性質(zhì)增強(qiáng)以及流體性質(zhì)減弱。整個(gè)凝膠體系tanδ<1,表明該模式下蠶蛹不溶性蛋白凝膠體系仍為弱凝膠體系。
G′和G″值在不同pH下發(fā)生變化的原因是pH變化會(huì)引起蛋白質(zhì)結(jié)構(gòu)變化,導(dǎo)致蛋白分子內(nèi)部網(wǎng)絡(luò)結(jié)構(gòu)的作用力受到了影響,從而引起了蛋白凝膠性能發(fā)生變化。可以根據(jù)在不同的pH下蛋白結(jié)構(gòu)是否發(fā)生變化來推測(cè)流變學(xué)特性發(fā)生改變的原因。
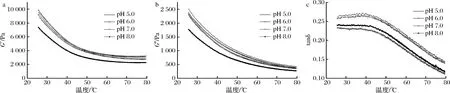
a-G′;b-G″;c-tanδ
2.3 不同pH處理對(duì)蠶蛹不溶性蛋白結(jié)構(gòu)的影響

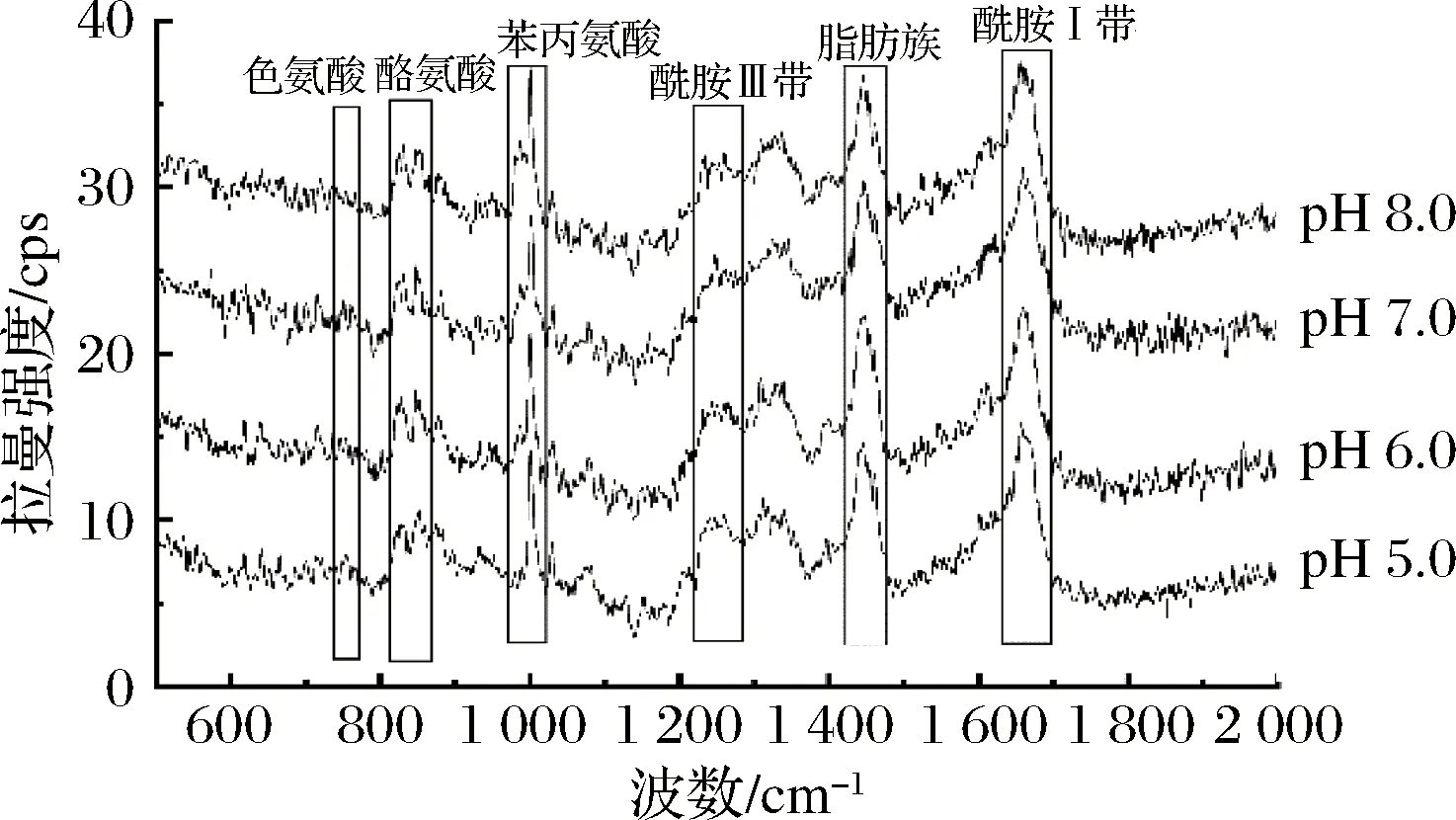
圖3 不同pH條件下的拉曼譜圖
酰胺I帶拉曼特征峰的結(jié)構(gòu)普遍歸屬[23]分為:α-螺旋結(jié)構(gòu)對(duì)應(yīng)范圍為1 645~1 658 cm-1;β-折疊結(jié)構(gòu)對(duì)應(yīng)范圍為1 665~1 680 cm-1;β-轉(zhuǎn)角結(jié)構(gòu)對(duì)應(yīng)范圍為1 640~1 644 cm-1與1 681~1 690 cm-1;無規(guī)卷曲結(jié)構(gòu)對(duì)應(yīng)范圍為1 659~1 664 cm-1。通常利用酰胺Ⅰ帶或者酰胺Ⅲ帶檢測(cè)蛋白質(zhì)的主鏈結(jié)構(gòu),根據(jù)拉曼原始數(shù)據(jù)擬合蛋白質(zhì)二級(jí)結(jié)構(gòu)的相對(duì)含量,本研究利用酰胺Ⅰ帶并根據(jù)ALIX等[24]的方法分析不同pH處理后蛋白質(zhì)樣品中的α-螺旋,β-折疊,β-轉(zhuǎn)角和無規(guī)卷曲的含量。
如圖4所示,α-螺旋相對(duì)含量在pH 5.0~8.0隨著pH的增大而增大,而β-轉(zhuǎn)角和無規(guī)則狀態(tài)的相對(duì)含量在此范圍內(nèi)減少,表明蛋白質(zhì)的二級(jí)結(jié)構(gòu)隨著pH的增大有序化度增加[25]。根據(jù)以上流變學(xué)結(jié)果顯示,無論是頻率掃描還是溫度掃描,pH 8.0時(shí)樣品的G′和G″模量相對(duì)較低,可能說明不溶性蠶蛹蛋白結(jié)構(gòu)有序化不利于凝膠網(wǎng)絡(luò)的形成。凝膠網(wǎng)絡(luò)的形成和蛋白質(zhì)間的相互作用有關(guān),進(jìn)一步通過微環(huán)境分析蛋白質(zhì)間相互作用變化趨勢(shì)。

圖4 不同pH條件下的二級(jí)結(jié)構(gòu)相對(duì)含量
760 cm-1附近的拉曼光譜帶歸屬為色氨酸側(cè)鏈,可用于研究色氨酸殘基的疏水性微環(huán)境的變化,當(dāng)色氨酸殘基從被包埋態(tài)疏水微環(huán)境變成被暴露態(tài)極性環(huán)境時(shí),760 cm-1附近的峰強(qiáng)度可能下降[26]。如圖5所示,在pH 6.0時(shí)的I760歸一化強(qiáng)度達(dá)到一個(gè)極大值,說明更多的色氨酸殘基沒有暴露在表面,可能是蛋白質(zhì)內(nèi)部更多的疏水作用形成而導(dǎo)致的疏水基團(tuán)包埋,結(jié)合流變學(xué)結(jié)果,pH 6.0時(shí)G′和G″模量較高,可能說明分子內(nèi)部疏水作用的形成,更有利于蠶蛹蛋白凝膠網(wǎng)絡(luò)的形成。當(dāng)色氨酸殘基趨于暴露時(shí),可能導(dǎo)致蛋白內(nèi)部疏水相互作用降低,不利于蠶蛹蛋白凝膠網(wǎng)絡(luò)的形成,G′和G″值出現(xiàn)降低趨勢(shì)。
酪氨酸的特征雙峰(850/830)是酪氨酸殘基上對(duì)位取代苯環(huán)的振動(dòng)引起的[27],其雙峰比值(I850/I830)可作為微環(huán)境及酚羥基上氫鍵的探針以及酪氨酸組分埋藏和暴露的程度[28],當(dāng)I850/I830的比值為1.25~1.40時(shí),表明此時(shí)酪氨酸殘基完全暴露于分子表面;當(dāng)比值為0.3~0.5時(shí),酪氨酸殘基完全包埋而藏于分子內(nèi)部;當(dāng)比值為0.7時(shí),酪氨酸殘基此時(shí)處于電離狀態(tài)[29]。由圖5可得,在pH 5.0~6.0,I850/I830比值均大于1,pH 6.0時(shí)I850/I830比值相對(duì)最低,此時(shí)酪氨酸暴露最少。酪氨酸作為氫鍵的供體[30],酪氨酸暴露減少說明分子間氫鍵作用減少,結(jié)合流變學(xué)結(jié)果,pH 6.0時(shí)G′和G″模量較高,此時(shí)氫鍵作用較弱,因此推測(cè)過多的氫鍵形成可能不利于不溶性蠶蛹蛋白凝膠網(wǎng)絡(luò)體系的形成。
在動(dòng)物性蛋白中,肌原纖維蛋白的熱誘導(dǎo)凝膠過程中疏水作用和二硫鍵的形成起到重要作用[15],而膠原蛋白類凝膠形成過程中氫鍵起到更重要的作用[31],維系不溶性蠶蛹蛋白凝膠特性的可能主要是疏水作用,由此可見,其凝膠維系方式可能與肌原纖維蛋白更加接近。

圖5 不同pH條件下的歸一化強(qiáng)度
2.4 不同鹽濃度處理對(duì)蠶蛹不溶性蛋白流變性能的影響
除了pH的影響外,離子強(qiáng)度會(huì)引起蛋白分子內(nèi)部結(jié)構(gòu)及分子間作用力發(fā)生變化,從而引起蛋白凝膠性質(zhì)發(fā)生變化。依據(jù)肉糜蛋白類制品加工過程中常用鹽濃度,探究不同鹽濃度處理后蛋白質(zhì)的G′、G″值和tanδ值的變化情況,進(jìn)行頻率掃描后的變化如上圖所示。由圖6-a、圖6-b可以看出,隨著頻率的增加,不同鹽濃度條件下的G′和G″值呈現(xiàn)增加趨勢(shì),在相同的振蕩頻率下,低鹽質(zhì)量濃度(10~20 g/L)的G′值小于對(duì)照組,與BERGROS等[32]類似,鹽的加入影響了羅非魚蛋白凝膠的G′值,鹽處理后的蛋白G′值明顯低于未加鹽處理的蛋白;中高鹽濃度(25~35 g/L)的G′值高于對(duì)照組,可能是鹽影響了蛋白分子間的作用,導(dǎo)致蛋白質(zhì)凝膠網(wǎng)絡(luò)的穩(wěn)定性發(fā)生變化。在鹽質(zhì)量濃度為35 g/L時(shí),體系的G′和G″值相對(duì)最高,表明此條件下蛋白質(zhì)分子間的相互作用較強(qiáng),此時(shí)有利于蛋白凝膠網(wǎng)絡(luò)的形成,流變性能較好。由圖6-c可以看出,tanδ值隨著頻率的增加出現(xiàn)下降趨勢(shì),tanδ值的下降表明樣品彈性增強(qiáng)。在1~10 Hz,對(duì)照組的tanδ最小,此時(shí)體系彈性最好,經(jīng)過鹽處理后蛋白體系彈性下降。在鹽質(zhì)量濃度為0~35 g/L時(shí),所有體系的G′值明顯大于G″值,tanδ值分布在0.12~0.24,表明蠶蛹不溶性蛋白凝膠體系屬于弱凝膠體系。
由圖7-a、圖7-b可以看出,隨著溫度的升高,不同鹽質(zhì)量濃度(0~35 g/L)處理后蛋白的G′和G″值總體呈現(xiàn)下降趨勢(shì),G′值在25~70 ℃出現(xiàn)快速下降的趨勢(shì),70 ℃后出現(xiàn)一個(gè)相對(duì)平穩(wěn)的平臺(tái)期;G″值在25~70 ℃出現(xiàn)快速下降的趨勢(shì),70 ℃后出現(xiàn)一個(gè)較小的下降趨勢(shì);在相同掃描溫度下,在鹽質(zhì)量濃度為35 g/L時(shí)的G′和G″值最高,此條件下蛋白質(zhì)分子間的相互作用較強(qiáng),有利于蛋白質(zhì)凝膠網(wǎng)絡(luò)的形成,流變性能較好;由圖7-c可以看出,tanδ值總體呈現(xiàn)下降趨勢(shì),隨著溫度的升高,tanδ值在25~40 ℃時(shí)呈現(xiàn)先緩慢下降趨勢(shì),40 ℃之后出現(xiàn)快速下降的趨勢(shì)。該掃描模式下tanδ值曲線分布密集,tanδ值總體分布范圍在0.12~0.24,G′明顯大于G″值,表明蠶蛹不溶性蛋白凝膠體系屬于弱凝膠體系。
G′和G″值在不同鹽濃度下發(fā)生變化的原因是離子濃度的變化可能會(huì)引起蛋白結(jié)構(gòu)變化,導(dǎo)致蛋白內(nèi)部分子間維持凝膠網(wǎng)絡(luò)結(jié)構(gòu)的作用力受到影響,從而引起了蛋白凝膠性能發(fā)生變化。可以根據(jù)在不同的鹽濃度下蛋白結(jié)構(gòu)是否發(fā)生變化來推測(cè)流變學(xué)特性發(fā)生改變的原因。

a-G′;b-G″;c-tanδ
2.5 不同鹽濃度處理對(duì)蠶蛹不溶性蛋白結(jié)構(gòu)的影響
圖8為不同鹽質(zhì)量濃度(0~35 g/L)處理后蛋白樣品的拉曼光譜曲線。不同鹽濃度處理后的拉曼光譜圖基本一致,利用酰胺Ⅰ帶并根據(jù)ALIX等[24]的方法分析不同鹽濃度處理后蛋白樣品中的α-螺旋,β-折疊,β-轉(zhuǎn)角和無規(guī)卷曲的含量。
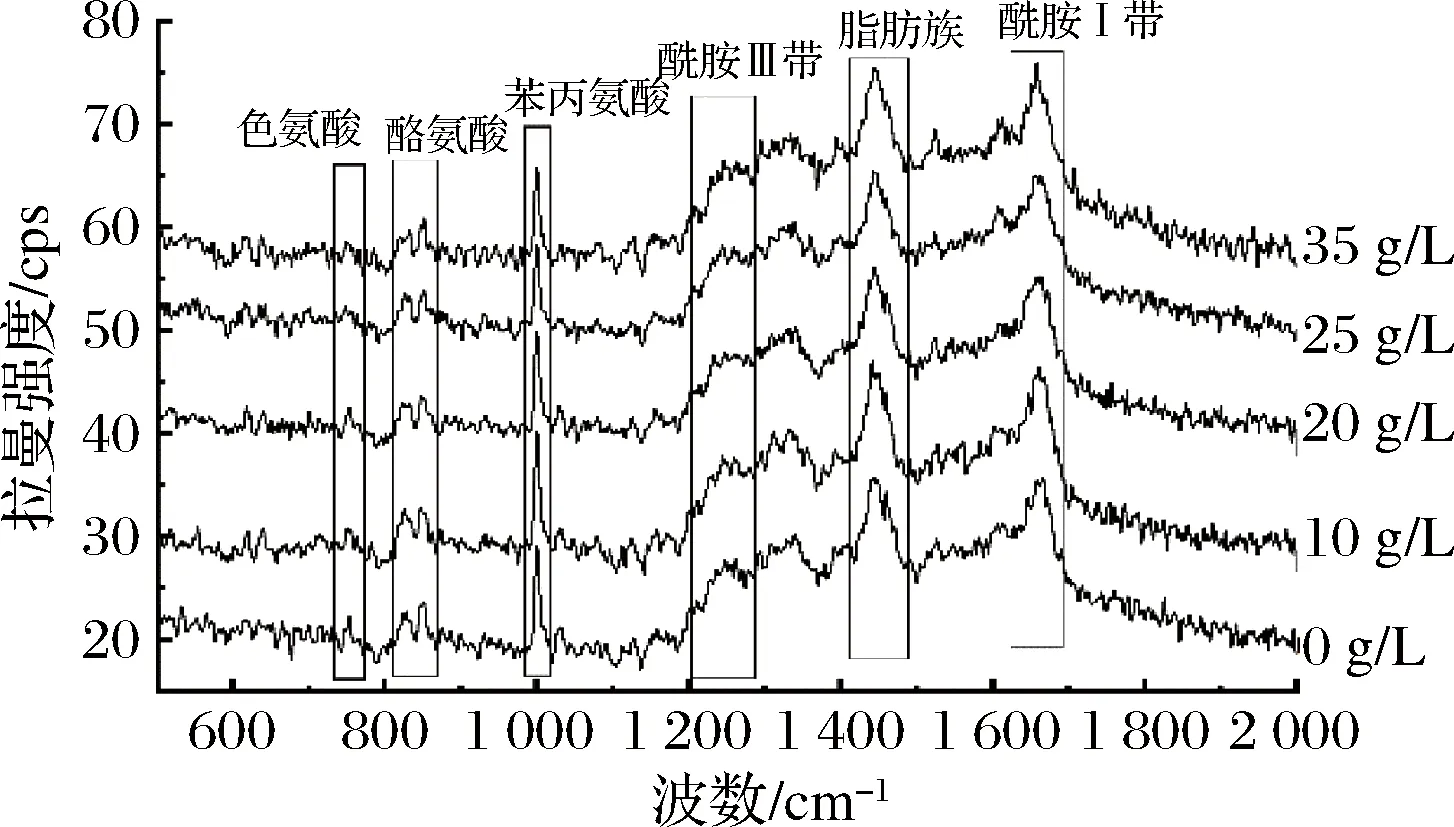
圖8 不同鹽濃度條件下的拉曼譜圖
如圖9所示,在鹽質(zhì)量濃度為0~35 g/L時(shí),α-螺旋和β-折疊的相對(duì)含量變化不明顯,α-螺旋和β-折疊的相對(duì)含量均分布在65%~75%,表明此條件下的蛋白質(zhì)內(nèi)部結(jié)構(gòu)比較穩(wěn)定有序。
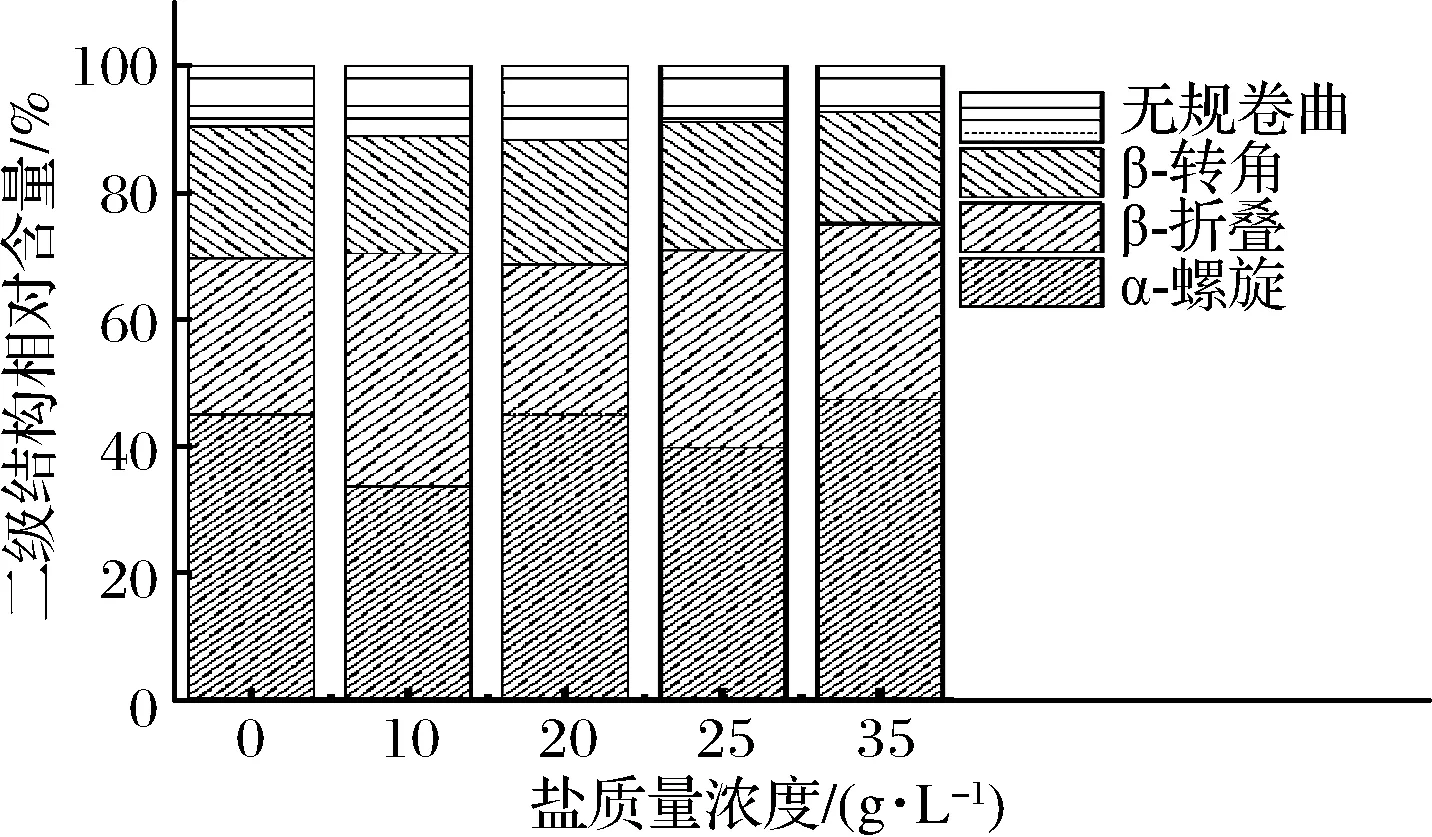
圖9 不同鹽濃度條件下的二級(jí)結(jié)構(gòu)相對(duì)含量
如圖10所示,在鹽質(zhì)量濃度范圍為0~35 g/L,鹽質(zhì)量濃度為35 g/L時(shí),I760強(qiáng)度達(dá)到一個(gè)極大值,色氨酸趨于包埋,說明此時(shí)更多的色氨酸殘基沒有暴露在表面,可能是蛋白內(nèi)部更多的疏水作用形成而導(dǎo)致的疏水基團(tuán)包埋,結(jié)合流變學(xué)結(jié)果,鹽質(zhì)量濃度35 g/L時(shí)G′和G″模量較高,可能說明分子內(nèi)部疏水作用的形成,更有利于蠶蛹蛋白質(zhì)凝膠網(wǎng)絡(luò)的形成。當(dāng)色氨酸殘基趨于暴露時(shí),可能導(dǎo)致蛋白質(zhì)內(nèi)部疏水相互作用降低,不利于蠶蛹蛋白凝膠網(wǎng)絡(luò)的形成,G′和G″值出現(xiàn)降低趨勢(shì)。
由圖10可得,在鹽質(zhì)量濃度范圍0~35 g/L內(nèi),鹽質(zhì)量濃度為35 g/L時(shí)I850/I830比值最小,此時(shí)酪氨酸暴露最少,酪氨酸作為氫鍵的供體,酪氨酸暴露減少說明分子間氫鍵作用減少,結(jié)合流變學(xué)結(jié)果,此時(shí)G′和G″模量較高,氫鍵作用較弱,因此推測(cè)過多的氫鍵形成可能不利于不溶性蠶蛹蛋白質(zhì)凝膠網(wǎng)絡(luò)體系的形成。當(dāng)酪氨酸暴露增多時(shí),不利于蛋白質(zhì)凝膠網(wǎng)絡(luò)體系的形成,可能導(dǎo)致流變學(xué)特性變差,G′和G″值出現(xiàn)降低趨勢(shì)。
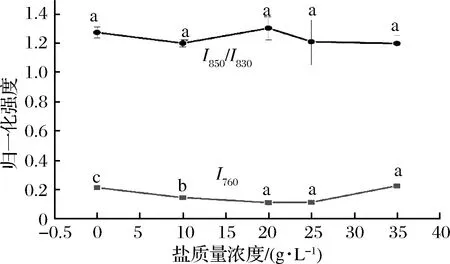
圖10 不同鹽濃度條件下的歸一化強(qiáng)度
2.6 TG酶處理對(duì)蠶蛹不溶性蛋白流變性能的影響
TG酶是一種催化蛋白質(zhì)分子側(cè)鏈中谷氨胺殘基與賴氨酸之間形成ε-(γ-谷氨酰基)賴氨酸共價(jià)鍵的蛋白酶,通常用于蛋白質(zhì)凝膠特性的改善[33]。由圖11-a~圖11-c可以看出,隨著頻率增大,蛋白體系的G′和G″值總體呈現(xiàn)上升趨勢(shì);由圖11-d~圖11-f可以看出,隨著溫度的升高,蛋白體系的G′和G″值總體呈現(xiàn)下降趨勢(shì)。無論是頻率掃描還是溫度掃描,蛋白凝膠體系的黏性模量(G″)始終小于彈性模量(G′),tanδ值<1,表明蠶蛹不溶性蛋白質(zhì)凝膠仍是彈性占主導(dǎo)地位的弱凝膠體系。加入TG酶處理后的蛋白體系的G′和G″模量明顯增大,黏彈性增強(qiáng),表明TG酶的添加可有效改善蠶蛹不溶性蛋白的凝膠特性。
在典型加工條件下,不溶性蠶蛹蛋白呈現(xiàn)弱凝膠特性,而TG酶可以顯著增強(qiáng)其凝膠特性,這表明在TG酶交聯(lián)條件下,不溶性蠶蛹蛋白具有開發(fā)凝膠類蛋白質(zhì)制品或作為在其中添加使用的可行性。
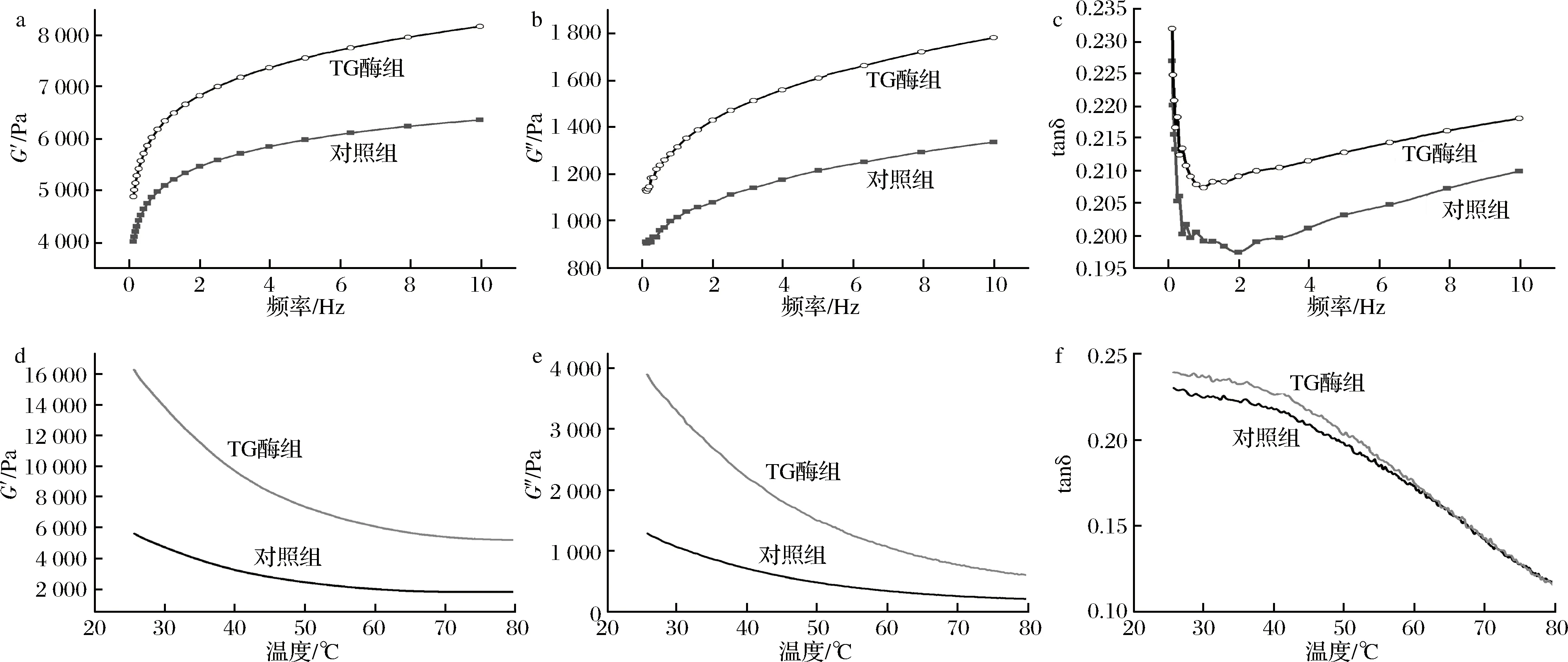
a-G′;b-G″;c-tanδ
3 結(jié)論
(1)不溶性蠶蛹蛋白中必需氨基酸含量40%,是一種營養(yǎng)價(jià)值較高的蛋白質(zhì)資源,且疏水性氨基酸比例達(dá)到40%,有蛋白質(zhì)分子間通過疏水作用力形成不透明凝膠的可能。
(2)不溶性蠶蛹蛋白經(jīng)過不同的pH和不同鹽濃度處理后,經(jīng)過流變學(xué)頻率掃描和溫度掃描顯示,在pH 6.0、鹽質(zhì)量濃度為35 g/L時(shí),不溶性蛋白體系的G′值和G″值最高,黏彈性最好;在pH 5.0~8.0、鹽質(zhì)量濃度為0~35 g/L時(shí),不溶性蛋白質(zhì)凝膠體系的G′值明顯大于G″值,tanδ<1,表明不溶性蛋白質(zhì)凝膠是一種弱凝膠。拉曼光譜學(xué)顯示,pH增大會(huì)引起蛋白質(zhì)結(jié)構(gòu)有序化增多,但不利于蛋白質(zhì)網(wǎng)絡(luò)的形成,不溶性蛋白體系的G′值和G″值在pH 8.0時(shí)最低。色氨酸的暴露與疏水相互作用有關(guān),色氨酸在pH 6.0及鹽質(zhì)量濃度35 g/L時(shí)趨于包埋態(tài),分子內(nèi)部疏水作用的形成,更有利于蠶蛹蛋白質(zhì)凝膠網(wǎng)絡(luò)的形成,此時(shí)G′值和G″值最高。當(dāng)色氨酸殘基趨于暴露時(shí),可能導(dǎo)致蛋白質(zhì)內(nèi)部疏水相互作用降低,不利于蠶蛹蛋白質(zhì)凝膠網(wǎng)絡(luò)的形成。酪氨酸的暴露與氫鍵作用力有關(guān),酪氨酸在pH 6.0時(shí)及鹽質(zhì)量濃度35 g/L暴露最少,說明分子間氫鍵作用減少,結(jié)合流變學(xué)結(jié)果,推測(cè)過多的氫鍵形成可能不利于不溶性蠶蛹蛋白凝膠網(wǎng)絡(luò)體系的形成。疏水相互作用和氫鍵可能是影響蛋白質(zhì)凝膠網(wǎng)絡(luò)的的關(guān)鍵作用力,蛋白微環(huán)境發(fā)生變化時(shí),維持蛋白質(zhì)凝膠網(wǎng)絡(luò)結(jié)構(gòu)的作用力也會(huì)發(fā)生變化,流變學(xué)特征的G′值和G″值發(fā)生變化。
(3)加入TG酶處理后的蛋白體系的G′和G″模量明顯增大,黏彈性增強(qiáng),表明TG酶的添加可改善不溶性蛋白的凝膠特性。實(shí)驗(yàn)結(jié)果可為不溶性蠶蛹蛋白質(zhì)的加工提供一定的理論參考。
猜你喜歡
商品與質(zhì)量(2021年43期)2022-01-18 05:31:22
杭州(2020年23期)2021-01-11 00:54:42
新世紀(jì)智能(數(shù)學(xué)備考)(2020年11期)2021-01-04 00:38:16
中國外匯(2019年17期)2019-11-16 09:31:14
中國衛(wèi)生(2015年12期)2015-11-10 05:13:40
現(xiàn)代企業(yè)(2015年1期)2015-02-28 18:43:18
汽車零部件(2014年5期)2014-11-11 12:24:28
新高考·高一物理(2014年1期)2014-09-18 01:26:07
浙江人大(2014年1期)2014-03-20 16:19:53
終身教育研究(2012年4期)2012-03-25 10:41:11
