枯否細胞在肝移植脂肪供肝的缺血再灌注損傷中的作用研究進展
2021-01-28 07:54:02蔡佳暉王經(jīng)琳任昊楨施曉雷
肝膽胰外科雜志 2021年1期
蔡佳暉,王經(jīng)琳,任昊楨,施曉雷,
(1.南京醫(yī)科大學附屬鼓樓臨床醫(yī)學院 普外科,2.南京大學醫(yī)學院附屬鼓樓醫(yī)院 普外科,江蘇 南京210008)
缺血再灌注損傷(ischemia-reperfusion injury,IRI)是指缺血、缺氧的組織或器官在重新獲得血液及氧氣供應后,損害不但沒有減輕,反而加重的病理生理過程。一般分為四個階段:器官離開供體到冷凍前的熱缺血期,冷藏器官以降低代謝率的冷缺血期,器官與肝移植患者進行血管重建的復溫期,血流再通的再灌注期。肝臟IRI是肝移植的常見風險,影響手術的成功與否,也決定了移植后器官功能的恢復情況。
目前,肝移植患者的數(shù)量遠超可用供肝的數(shù)量,諸如脂肪供肝這類邊緣供肝的應用就愈發(fā)普遍[1]。然而,相較于正常供肝,脂肪供肝線粒體ATP生產(chǎn)能力下降[2],這使其對IRI的耐受能力下降。同時,脂肪供肝內(nèi)的細胞色素P450酶代謝多余的脂肪酸并生成大量活性氧(reactive oxygen species,ROS),導致枯否細胞(Kupffer cells,KCs)對凋亡信號更為敏感,促進了脂肪供肝的細胞凋亡。細胞凋亡需要ATP,當ATP耗竭,凋亡轉變?yōu)閴乃溃灾靖蔚奶攸c是中度凋亡和大量壞死(73%),這種細胞死亡類型的改變或許增強了脂肪供肝對促炎因子的敏感性[3]。而在IRI過程中,經(jīng)典活化的KCs將釋放大量炎癥介質,如腫瘤壞死因子-α(tumor necrosis factor-alpha,TNF-α)、白細胞介素(IL)-1β、IL-18、血小板活化因子(platelet activating facter,PAF)、不同的前列腺素、干擾素(interferon gamma IFN)等,對組織產(chǎn)生有害作用。這表明在IRI過程中,KCs對脂肪供肝有著重要的影響。
移植前,脂肪肝中的KCs 體積增大,數(shù)量增多,且處于激活狀態(tài)[4]。當發(fā)生IRI時,KCs進一步激活,這與再灌注時大量內(nèi)毒素(LPS)的涌入有關,也與肝內(nèi)的氧化應激反應有關,由于KCs本身的細胞特性,還與其極化狀態(tài)的改變有關,但具體機制尚未明確。
1 脂肪肝IRI的病理生理
隨著現(xiàn)代人飲食結構的改變,高脂高熱量飲食愈發(fā)普遍,這就導致非酒精性脂肪肝病(NAFLD)人數(shù)日益增多,在亞洲地區(qū)NAFLD的總體患病率為29.62%[5]。當肝臟發(fā)生脂肪變性,肝細胞內(nèi)的脂滴使細胞脹大,一方面導致肝血竇腔隙減小,血流不足,肝細胞缺氧壞死,另一方面保存器官時易發(fā)生脂肪固結,引起再灌注時肝細胞崩解。脂肪變性也改變了線粒體膜的脂質組成和流動性,使其易受到自由基的攻擊,同時,細胞器的主要抗氧化物質還原型谷胱甘肽(GSH)也發(fā)生下降。于是在IRI時,大量氧自由基生成,脂肪肝內(nèi)脂質過氧化,氧自由基直接損傷細胞組成,或產(chǎn)生炎性因子和黏附分子造成間接損害。KCs的激活是脂肪肝IRI加劇的另一原因,這一點將在下文著重介紹。
2 KCs的基本形態(tài)
KCs是一種肝固有巨噬細胞,來源于骨髓中的單個核細胞[6],占全身巨噬細胞總數(shù)的80%~90%,約占肝臟細胞總數(shù)的15%[7]。KCs體積較大,形態(tài)不規(guī)則,可伸出絲狀偽足依附于肝血竇內(nèi)皮細胞表面或經(jīng)窗孔伸入Disse間隙內(nèi),與肝細胞微絨毛交錯,這樣的結構方便其協(xié)調或影響其他細胞的功能。KCs含有多種溶酶體酶,其質膜表面可表達多種受體,如Toll樣受體(toll like receptor,TLR)、內(nèi)毒素受體、清道夫受體、Fc受體、補體受體、半乳糖胺受體等。KCs的結構及酶學特點均表明其具備吞噬、分泌、免疫調節(jié)與監(jiān)視的作用。在受到刺激后,KCs將釋放大量細胞因子、趨化因子、炎癥介質、活性氧中間產(chǎn)物和一些其他的因子[8]。
3 KCs的功能
KCs在多種肝臟疾病中發(fā)揮重要作用。發(fā)生急性肝衰竭時,損傷相關分子模式(DAMPs)釋放,激活固有免疫,KCs通過釋放炎癥因子和趨化因子產(chǎn)生炎癥環(huán)境;在NAFLD中,KCs被激活,TNF-α、IL等多種細胞因子,使NAFLD向非酒精性脂肪肝炎(NASH)進展;KCs與自然殺傷(NK)細胞相互作用可能是肝纖維化過程中的一個調控因素[9],這對預防肝硬化有積極意義。
KCs在肝移植的IRI中也有重要作用。在肝臟IRI的熱缺血期,由于肝臟處于相對缺氧狀態(tài),無氧酵解增多易引起代謝性酸中毒,同時ATP的減少導致細胞膜離子交換紊亂,在這些因素的影響下,細胞內(nèi)鈣離子濃度上升,形成鈣超載。增多的鈣離子,一方面使黃嘌呤脫氫酶轉化為黃嘌呤氧化酶,這一轉變將加速氧自由基的產(chǎn)生,另一方面也激活了KCs,釋放細胞因子損傷肝臟。此外,冷缺血期主要損害肝非實質性細胞,機制與熱缺血期類似,在這一階段,NF-κB受到應激,活性增強,使細胞因子基因表達增多[10]。在再灌注期,大量LPS隨血流進入肝臟,與KCs表面受體結合,釋放細胞因子,導致肝臟炎癥反應[11]。顯然,在整個肝移植IRI過程中,KCs都扮演著重要角色,因此,明確KCs的作用機制將有助于提高肝移植術后供體肝的存活率與功能。
4 KCs加重脂肪供肝IRI進程的機制
脂肪供肝一般來源于NAFLD患者,其肝臟內(nèi)的KCs處于激活狀態(tài)[4],這是NAFLD發(fā)展為NASH的重要原因,也就是說相較于正常供肝,脂肪供肝本身的基礎條件較差。當發(fā)生肝臟IRI,脂肪供肝內(nèi)的KCs進一步激活,這可能與大量LPS的涌入、氧化應激反應以及極化狀態(tài)的轉變有關。
4.1 加重脂肪供肝IRI時KCs的激活與大量LPS的涌入有關
門靜脈血中含有許多微生物相關抗原,主要為革蘭氏陰性菌細胞壁成分LPS。正常情況下,外周靜脈血中的LPS含量極低,無法測得,但在門靜脈血中高達1.0 μg/L。在肝移植過程中,隨著再灌注的開始,門靜脈血流開放,KCs被進入肝臟的LPS進一步激活。KCs質膜表面可表達多種受體,LPS一方面與LPS受體CD14結合,激活KCs釋放炎癥介質,另一方面與TLR4結合,通過引起KCs內(nèi)鈣超載激活NF-κB[12],大量釋放炎性因子、氧自由基及趨化因子。正常肝臟內(nèi)的KCs通過上調抗炎基因精氨酸酶1和IL-10來抑制LPS介導的炎癥反應,實現(xiàn)LPS耐受,但當肝臟存在脂肪變性時,這種保護機制受到了損害[13],所以在脂肪肝中,LPS涌入激活KCs后造成更重的炎癥損傷。
LPS激活KCs分泌多種炎性因子,包括TNF-α、INF、IL等。TNF-α是主要的炎性因子之一,低水平的TNF-α是肝細胞生長分化與再生所必需的調節(jié)因子,而高水平的TNF-α主要誘導肝細胞的凋亡與壞死。正常狀態(tài)下TNF-α可以調控細胞的增殖、炎癥、凋亡等一系列病理生理過程;而在脂肪肝IRI中,大量釋放的TNF-α則導致肝臟損傷。已有研究表明,TNF-α的水平與供肝的損傷程度呈正相關,高濃度TNF-α是LPS引起肝臟炎癥反應以及肝細胞凋亡的主要原因[14]。IL在傳遞信息、激活調節(jié)免疫細胞及介導炎癥反應中起重要作用,其中IL-1β、IL-18等促炎因子主要來源于NLRP3炎癥小體。NLRP3被KCs激活后發(fā)生寡聚化并招募適配器蛋白,然后與caspase-1結合,形成炎癥小體,導致caspase-1的裂解和活化,促進了IL-1β和IL-18的成熟和分泌[15]。INF是一種主要用于抗病毒的糖蛋白,同時也有免疫調節(jié)的作用,它與TNF-α以及IL-2、IL-12都能增強NK細胞活性,引起更強的移植物免疫排斥反應。所有這些細胞因子,除了介導炎癥反應,還會導致細胞間黏附分子(intercellular adhesion molecule,ICAM)和內(nèi)皮細胞黏附分子(endothelial cell adhesion molecule,ECAM)表達的上調,同時,血管內(nèi)皮細胞產(chǎn)生了PAF[16]。因此,大量白細胞和血小板聚集于肝血竇區(qū),引發(fā)“無復流”現(xiàn)象,使肝臟缺血缺氧,間接對肝臟造成了損害。
趨化因子是一類可溶性細胞因子,它作為趨化劑引導細胞,特別是免疫細胞的遷移[17]。KCs經(jīng)LPS激活后產(chǎn)生的趨化因子CCL2(也稱為MCP1)是肝內(nèi)主要的促纖維化細胞,它可以聚集淋巴細胞和中性粒細胞浸潤肝組織,釋放蛋白水解酶,損傷內(nèi)皮細胞和肝實質細胞。同時,大量細胞的聚集加劇了前文所述炎癥因子引起的肝血竇區(qū)“無復流”現(xiàn)象,使得肝內(nèi)血流灌注進一步減少,加重肝組織損傷[18]。
4.2 加重脂肪供肝IRI時KCs的激活與氧化應激有關
KCs的氧化應激是早期損傷的表現(xiàn),此時KCs產(chǎn)生和釋放活性氧(reactive oxygen species,ROS),包括超氧陰離子和過氧化氫,對肝臟造成急性損傷[19]。ROS一方面可以損傷呼吸鏈中的酶復合物,引起細胞凋亡,另一方面可以上調促炎因子,造成炎癥損傷。當發(fā)生IRI時,由炎癥小體介導的無菌性炎癥反應過程被啟動,其中NLRP3炎性小體發(fā)揮主要作用[20],而KCs產(chǎn)生的大量ROS可通過各種途徑激活NLRP3炎性小體,最終產(chǎn)生炎癥反應。在脂肪供肝中,會產(chǎn)生更多的ROS,加重了肝臟IRI[21]。Yang等[22]認為,其中的機制可能與以下有關:(1)脂肪變性導致細胞體積增大,肝血竇減小,脂肪肝本就處在相對缺血缺氧的環(huán)境;(2)線粒體氧化不足,生成的ATP無法滿足缺血再灌注過程中的能量需求,就會產(chǎn)生更多的ROS;(3)脂肪肝內(nèi)KCs處于應激狀態(tài),非IRI狀態(tài)下也會生成炎癥因子。也有研究指出,脂肪供肝在IRI過程中線粒體損傷加重,線粒體氧化應激和脂質過氧化作用增加,除了ROS的增多,抗氧化防御機制也受到損害,因此受到更重損傷。
4.3 脂肪供肝IRI時KCs的激活與極化狀態(tài)的轉變有關
KCs有兩種表型,M1型為經(jīng)典激活型,參與機體炎癥、防御反應,此時主要釋放TNF-α等促炎因子;M2型為替代激活型,參與機體抗炎、修復作用,此時IL-10等抗炎因子顯著增多。當環(huán)境發(fā)生改變,KCs誘導分化為不同表型,發(fā)揮不同的功能,在適當條件下兩種表型還可相互轉化[23]。正常狀態(tài)KCs為M2型,當肝臟發(fā)生脂肪變性,分解出的一些促炎脂肪酸將募集單核細胞,并在脂肪組織內(nèi)將其轉化為M1型的KCs,在IRI過程中,TLR的激活以及ROS的產(chǎn)生,也會將KCs轉化為M1型。所以在肝移植過程中,脂肪供肝內(nèi)的KCs偏向M1型,其炎癥反應也強于正常供肝[24]。
5 KCs減輕脂肪供肝IRI的機制
除了損傷作用,KCs對肝臟也存在著保護作用,前文提到替代激活的M2型KCs參與機體抗炎,并能修復組織,這主要通過產(chǎn)生一些生物活性物質或上調表達一些酶類來實現(xiàn)。M2型KCs可以產(chǎn)生抑制性細胞因子IL-10,使T細胞失功能或無功能,或是表達凋亡相關因子配體(FasL)和腫瘤壞死因子相關凋亡誘導配體(TRAIL),引發(fā)T細胞凋亡,從而實現(xiàn)肝臟的免疫耐受[25],這將減輕缺血再灌注損害后移植物的排斥反應。另外,KCs可以生成可誘導一氧化氮合酶(inducible nitric oxide synthase,iNOS),KCs激活iNOS后,促進了NO的生成。NO是一種重要的血管擴張劑,有研究證實NO因其抑制血小板聚集、擴張血管的功能,在肝的缺血再灌注損傷中發(fā)揮了保護作用[26]。同時,KCs會生成血紅素加氧酶-1(hemeoxygenase-1,HO-1),主要分布于KCs的內(nèi)質網(wǎng)與核周外模,具有抗氧化、抗炎、抗凋亡以及抑制血小板聚集的作用[27]。Zeng 等[11]證明,HO-1增加后,大鼠血清中多種炎性因子的含量下降,從而減輕IRI。還有研究證明,外源性上調HO-1表達后,可以顯著保護肥胖Zucker大鼠的脂肪化肝臟,減少脂肪肝IRI[28]。HO-1還可以通過降解血紅素的產(chǎn)物CO來發(fā)揮保護作用。CO可以使KCs生成的TNF-α和IL-6減少,也可以下調因LPS激活KCs產(chǎn)生的炎癥反應,這些都改善了肝臟IRI。
總的來說,LPS與KCs表面受體CD14及TLR4結合后激活NF-κB,一方面釋放大量炎癥因子,包括TNF-α、INF、IL-2、IL-12,在造成直接肝臟損傷的同時,導致ICAM與ECAM表達上調,造成間接肝臟損害;同時也生成更多的ROS,激活NLRP3炎癥小體生成IL-1β和IL-18,引起炎癥反應,加重肝臟IRI。另一方面釋放IL-10及HO-1,HO-1還能生成CO,這些細胞因子都可以抑制炎癥因子,減輕炎癥反應;而其釋放的iNOS可以生成血管擴張劑NO,對肝臟IRI也有緩解作用(圖1)。
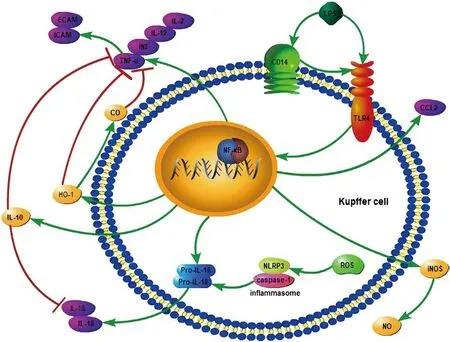
圖1 LPS激活KCs引起雙向作用:LPS與KCs表面受體CD14及TLR4結合后激活NF-κB,一方面釋放大量炎癥因子,包括TNF-α、INF、IL-2、IL-12,導致ICAM與ECAM表達上調;同時生成更多ROS,激活NLRP3炎癥小體生成IL-1β和IL-18。另一方面釋放抗炎因子,如IL-10、HO-1及iNOS,HO-1生成CO,iNOS生成NO。
6 小結
IRI是肝移植術后供體肝功能受損和失功能的重要原因,是多因子共同作用造成損傷的持續(xù)過程,KCs在其中起到重要作用。對于脂肪肝供肝,肝實質細胞脂肪變性及肝非實質細胞激活狀態(tài)使其處在一個相對缺血缺氧且促炎的環(huán)境。所以當IRI發(fā)生,LPS涌入后激活KCs釋放大量細胞因子,脂肪肝表現(xiàn)出更差的耐受力,而脂肪變性導致的氧化應激加劇以及KCs極化狀態(tài)向促炎型改變也使脂肪肝在IRI中受到更重的損傷。除了損傷作用,KCs對肝臟也存在著保護作用,激活的M2型KCs參與機體抗炎,并能修復組織。當前,脂肪供肝在肝移植術中應用增多是無法逆轉的趨勢,研究如何減輕脂肪肝IRI具有重要意義,而KCs顯然是一個重要的靶點。
