TGF-β1對人皮膚鱗狀細胞癌A431細胞增殖的影響及可能的機制*
2021-01-27 14:49:02何仁穎李航宇
檢驗醫(yī)學(xué)與臨床 2021年2期
何仁穎,何 威,李航宇,張 斌,4△
1.中國人民解放軍陸軍軍醫(yī)大學(xué)第二附屬醫(yī)院皮膚科,重慶 400037;2.重慶市人民醫(yī)院皮膚科,重慶 400014;3.中國人民解放軍陸軍軍醫(yī)大學(xué)第一附屬醫(yī)院燒傷科,重慶 400038;4.重慶市中醫(yī)院/重慶市第一人民醫(yī)院皮膚科,重慶 400011
皮膚鱗狀細胞癌是一種來源于表皮角質(zhì)形成細胞的常見惡性腫瘤,好發(fā)于顏面部,且隨著時間的推移容易發(fā)生轉(zhuǎn)移,嚴(yán)重影響患者的外貌及健康。隨著環(huán)境污染的加重,鱗狀細胞癌的發(fā)病率逐年升高。近年來研究表明,轉(zhuǎn)化生長因子-β1(TGF-β1)在調(diào)節(jié)細胞生長、分化、凋亡和在維持皮膚內(nèi)環(huán)境穩(wěn)定方面均發(fā)揮著至關(guān)重要的作用,是重要的調(diào)節(jié)者和參與者[1]。既往研究發(fā)現(xiàn),在人皮膚鱗狀細胞癌組織中有TGF-β/Smad信號轉(zhuǎn)導(dǎo)通路的存在,且其中幾種TGF-β1受體的表達存在變化[2]。然而,TGF-β1的表達是否與人皮膚鱗狀細胞癌A431細胞的增殖過程有關(guān),相關(guān)報道甚少。為此,本研究通過觀察A431細胞和HaCaT細胞在不同TGF-β1作用濃度和作用時間后,細胞的增殖情況和細胞中TGF-β受體mRNA表達的變化情況,來進一步探討其可能的機制。
1 材料與方法
1.1材料來源 人角質(zhì)形成細胞 HaCaT 細胞和人皮膚鱗狀細胞癌A431 細胞均購于中國醫(yī)學(xué)科學(xué)院上海細胞所。
1.2儀器與試劑 人重組 TGF-β1 (美國 PEPROTECH 公司)、胎牛血清(天津灝洋生物制品科技責(zé)任有限公司)、DMEM培養(yǎng)液(美國 Gibco公司)、胰蛋白酶和 Tripure Isolation Reagent(美國Roche公司)、SYBR?Premix Ex TaqTM和 PrimeScriptTMRT Reagent Kit(日本TaKaRa Bio公司)、DMSO(常州市科豐化工有限公司)、MTT試劑盒(南京凱基生物科技發(fā)展有限公司)。凝膠成像分析儀(美國Alpha Innotech公司)、熒光定量 PCR 反應(yīng)擴增儀和酶標(biāo)儀(Model 550 version 2.24,美國Bio-Rad公司)、DU800核酸/蛋白檢測儀(美國Beckman Coulter公司)。
1.3方法
1.3.1細胞培養(yǎng)及分組 將HaCaT 細胞和A431細胞以每孔5×103個接種于96孔板,在5% CO2,37 ℃的細胞培養(yǎng)箱中,用含10%胎牛血清的DMEM液培養(yǎng)24 h至貼壁。按以下分組要求向細胞培養(yǎng)液中添加TGF-β1后換液培養(yǎng):(1)濃度組,按TGF-β1終濃度0、5、10、20、100 ng/mL分為5組,持續(xù)作用48 h。(2)時間組,TGF-β1終濃度為20 ng/mL時,分別作用12、24、48、72 h。另設(shè)空白對照。每組設(shè)6個復(fù)孔,結(jié)果重復(fù)檢測3次。
1.3.2MTT法檢測TGF-β1對細胞增殖的影響 將50 μL MTT溶液加入各孔,繼續(xù)培養(yǎng)4 h后棄上清液。加入DMSO液150 μL,充分震蕩溶解后于490 nm處測定各孔吸光度值(A490 nm),計算細胞活性率。
1.3.3RT-PCR測定TGF-β1對細胞中TGF-β受體mRNA表達的影響 分別將培養(yǎng)好的人皮膚鱗狀細胞癌A431細胞和HaCaT細胞進行RNA提取,并對各自RNA樣本的純度和濃度進行檢測。按照PrimeScriptTMRT Reagent Kit說明中的具體操作步驟進行反轉(zhuǎn)錄反應(yīng)。嚴(yán)格按照試劑盒操作說明配制相應(yīng)的反應(yīng)體系。擴增條件:95 ℃ 10 s,95 ℃ 5 s、60 ℃ 30 s,72 ℃ 45 s,共40個循環(huán)。采用2-ΔΔCt法進行分析。
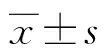
2 結(jié) 果
2.1相同作用時間下不同濃度的TGF-β1對細胞增殖的影響 在作用時間相同的情況下,不同濃度的TGF-β1對HaCaT細胞的增殖均能產(chǎn)生顯著的抑制作用,見圖1,與對照組相比,差異有統(tǒng)計學(xué)意義(P<0.05)。而在不同濃度的TGF-β1對A431細胞的增殖無明顯抑制作用,與對照組相比,差異無統(tǒng)計學(xué)意義(P>0.05),這提示TGF-β1對A431細胞產(chǎn)生的抑制作用不明顯。
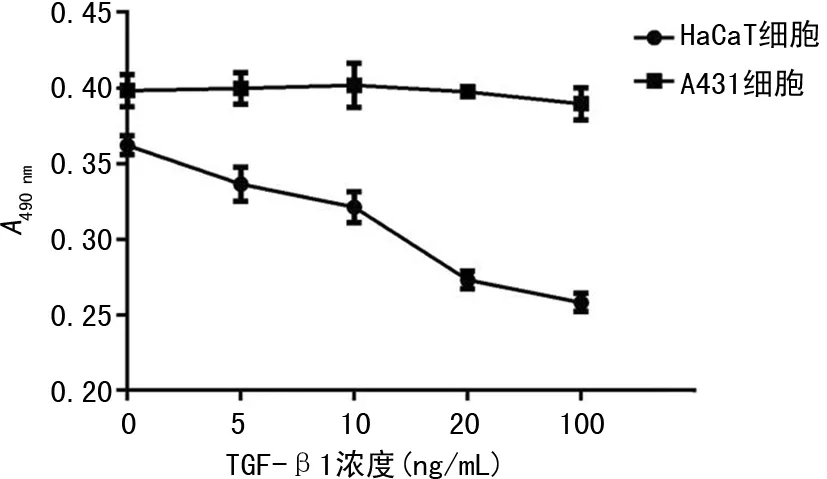
圖1 不同濃度TGF-β1對HaCaT細胞和A431細胞增殖的影響(MTT法)
2.2相同濃度的條件下TGF-β1不同作用時間對細胞增殖的影響 在TGF-β1濃度均為20 ng/mL的情況下,分別作用不同時間,TGF-β1對HaCaT細胞的增殖產(chǎn)生了明顯的抑制作用,見圖2,與對照組細胞相比,差異有統(tǒng)計學(xué)意義(P<0.05)。而TGF-β1對A431細胞增殖產(chǎn)生的抑制作用不明顯,與對照組細胞相比,差異無統(tǒng)計學(xué)意義(P>0.05),見圖3。
圖2 TGF-β1不同作用時間對HaCaT細胞增殖的影響
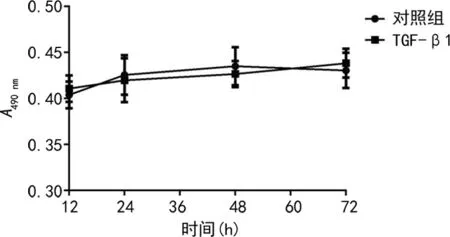
圖3 TGF-β1不同作用時間對A431細胞增殖的影響
2.3TGF-β1對HaCaT細胞TGF-β受體mRNA表達的影響 在相同的作用時間下(均為48 h),在一定濃度范圍內(nèi),隨著TGF-β1濃度的增加,HaCaT細胞的TGF-β受體Ⅰ mRNA(TGF-βRⅠ mRNA)、TGF-β受體Ⅱ mRNA(TGF-βRⅡ mRNA)的表達呈下降趨勢,見表1。在TGF-β1濃度均為20 ng/mL時,HaCaT細胞TGF-βRⅠ mRNA、TGF-βRⅡ mRNA的表達隨著TGF-β1作用時間的延長呈下降趨勢,見表2。
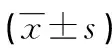
表1 不同濃度TGF-β1對HaCaT細胞TGF-β受體mRNA表達的影響
表2 TGF-β1不同作用時間對HaCaT細胞TGF-β受體mRNA表達的影響
2.4TGF-β1對A431細胞TGF-β受體mRNA表達的影響 在作用相同48 h條件下,在一定濃度范圍內(nèi),隨著TGF-β1濃度的增加,A431細胞的TGF-βRⅠ mRNA、TGF-βRⅡ mRNA的表達逐漸下降,見表3。在TGF-β1濃度均為20 ng/mL的條件下,A431細胞中TGF-βRⅠ mRNA的表達在一定時間范圍內(nèi),隨著TGF-β1作用時間的延長逐漸下降,見表4。
表3 不同濃度TGF-β1對A431細胞TGF-β受體mRNA表達的影響
表4 TGF-β1不同作用時間對A431細胞TGF-β受體mRNA表達的影響
3 討 論
TGF-β信號傳導(dǎo)通路十分復(fù)雜,主要由TGF-β受體介導(dǎo)。TGF-β受體分為Ⅰ型受體(TGF-βRⅠ)、Ⅱ型受體 (TGF-βRⅡ) 和Ⅲ型受體(TGF-βRⅢ)[3]。經(jīng)典的Smads信號通路主要是由TGF-β與TGF-βRⅡ的胞外段相結(jié)合后磷酸化激活TGF-βRⅠ,通過下游的Smad4蛋白完成TGF-β信號由細胞質(zhì)轉(zhuǎn)入細胞內(nèi)的過程[4]。TGF-βRⅢ因結(jié)構(gòu)上不含激酶活性區(qū),故不直接參與信號轉(zhuǎn)導(dǎo),主要起調(diào)節(jié)的作用[5]。TGF-β/Smad信號途徑異常與多種上皮來源的惡性腫瘤密切相關(guān),在乳腺癌、胃癌、前列腺癌、膀胱癌、子宮內(nèi)膜癌及宮頸癌等腫瘤中均存在TGF-β的過度表達;在人的頭頸部鱗狀細胞癌(HNSCC)、食管鱗狀細胞癌(ESCC)和皮膚鱗狀細胞癌中也存在TGF-β1過表達的情況[6]。TGF-β在腫瘤細胞中過度表達,不僅可以直接影響腫瘤細胞的浸潤和轉(zhuǎn)移,還能刺激血管生長,起到促進腫瘤發(fā)展的作用[7]。TGF-β 對腫瘤的直接影響可以通過Smads依賴途徑或干擾Smads依賴途徑介導(dǎo)完成,除經(jīng)典的Smads信號通路外[8-9],還有其他非經(jīng)典的不依賴于Smads蛋白信號通路的交互作用,如MAPK信號通路,它包括 ERK、JNK、p38、MAP 激酶等[10-11]。
對于大部分上皮細胞來源的惡性腫瘤而言,TGF-β1主要通過抑制腫瘤細胞的增殖,誘導(dǎo)細胞生長停滯和凋亡來發(fā)揮作用[3]。但也有研究者觀察到,部分腫瘤細胞對TGF-β的抑制作用不敏感,可以逃避TGF-β介導(dǎo)的生長抑制作用,這可能與腫瘤細胞表面TGF-β受體的表達異常有關(guān),同時,TGF-β 介導(dǎo)的抑制作用與TGF-βRⅠ或者TGF-βRⅡ的表達密切相關(guān),在人類ESCC中,約53.8%的患者表現(xiàn)出TGF-βRⅠ的表達降低,這與腫瘤的侵襲深度、轉(zhuǎn)移和病理分期有關(guān)[12]。在人小細胞肺癌細胞系中,HOUGAARD等[13]發(fā)現(xiàn)TGF-βRⅡ的表達明顯降低。此外,編碼TGF-β受體和Smads的基因耗竭或突變會導(dǎo)致小鼠模型自發(fā)性腫瘤的發(fā)生,且與人類癌癥的不良存活率相關(guān)[14]。如果兩種受體的表達存在缺陷,或者其中一種存在表達缺陷,就可能直接導(dǎo)致其抑制增殖的作用喪失。在HNSCC患者中也發(fā)現(xiàn),在細胞惡性轉(zhuǎn)化過程中,TGF-βRⅠ發(fā)生突變的概率較小,但70%以上的患者會出現(xiàn)TGF-βRⅡ表達的降低或消失[15]。TGF-βRⅡ耗竭可以使內(nèi)源性TGF-β1的表達增加,由此產(chǎn)生的TGF-β1過度表達可能會增加血管生成和炎性反應(yīng),從而促進HNSCC中腫瘤的進展[14]。TGF-βRⅡ發(fā)生突變的概率較大,較多見。研究表明,在口腔鱗狀細胞癌(OSCC)患者中TGF-βRⅡ表達出現(xiàn)降低或消失,在晚期OSCC患者中,TGF-βRⅡ的E221V/N238I突變可增強TGF-β信號傳導(dǎo)、導(dǎo)致更具侵入性的表型改變[16]。TGF-βRⅡ的缺陷與SCC的病情進展密切相關(guān)。
本研究也發(fā)現(xiàn),在一定劑量和作用時間范圍內(nèi),TGF-β1對HaCaT細胞增殖的抑制作用十分明顯;而在A431細胞中,TGF-β1的抑制作用卻不顯著。當(dāng)TGF-β1作用后,在A431細胞和HaCaT細胞中均出現(xiàn)了TGF-β受體mRNA表達水平的下降,說明與HaCaT細胞相比,TGF-β1對A431細胞的作用更加明顯。TGF-β受體是TGF-β信號傳導(dǎo)的必要條件,如果TGF-β受體表達水平下降或缺失,會導(dǎo)致TGF-β信號下傳受阻。由此推測,A431細胞可能通過誘導(dǎo)TGF-β受體的表達下調(diào),使得TGF-β信號下傳受到阻礙,其抑制作用失效,使得表皮內(nèi)環(huán)境穩(wěn)態(tài)遭到破壞,導(dǎo)致皮膚腫瘤細胞的抑癌基因功能喪失,癌基因被激活,最終導(dǎo)致腫瘤的形成。
猜你喜歡
中學(xué)生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
鴨綠江(2021年35期)2021-04-19 12:24:18
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學(xué)學(xué)報(2015年3期)2015-11-11 17:20:00