豬蛔蟲全基因組微衛(wèi)星分子記開發(fā)與特征分析
2021-01-21 05:59:24何晶晶黃建華牛紅艷周春花吳小平
中國獸醫(yī)雜志 2020年9期
何晶晶,黃建華,牛紅艷,周春花,吳小平
(南昌大學(xué)生命科學(xué)學(xué)院,江西 南昌 330031)
豬蛔蟲(AscarissuumGoeze,1782年)隸屬于線蟲綱蛔蟲目,主要寄生于豬小腸,是生豬感染的最常見寄生蟲之一,給養(yǎng)豬業(yè)造成巨大的經(jīng)濟(jì)損失[1]。同時豬蛔蟲也可以感染人,危害人類健康[2]。因此,許多國家已將其納入公共衛(wèi)生學(xué)范疇[3]。微衛(wèi)星標(biāo)記又稱短串聯(lián)重復(fù)序列(Simple tandem repeat,STR),廣泛分布于原核和真核生物基因組中,由1~6 bp的重復(fù)單元串聯(lián)而成,以雙核苷酸重復(fù)最為常見[4]。與其他分子標(biāo)記相比,微衛(wèi)星分子標(biāo)記因其存在廣泛、數(shù)量多、分布均勻、多態(tài)性信息豐富、遺傳共顯性、具有較好的穩(wěn)定性和重復(fù)性、易于檢測等優(yōu)點,成為目前遺傳多樣性和遺傳結(jié)構(gòu)研究中的首選標(biāo)記之一[5]。但目前沒有關(guān)于豬蛔蟲的微衛(wèi)星分子標(biāo)記相關(guān)信息的報導(dǎo)。雖然有學(xué)者應(yīng)用微衛(wèi)星標(biāo)記研究豬蛔蟲,但這些標(biāo)記都是基于其近緣種人蛔蟲而開發(fā)[6-8]。近年來隨著測序技術(shù)的發(fā)展,從全基因組水平研究物種的微衛(wèi)星,一方面可以了解不同物種微衛(wèi)星的生物信息學(xué)特征,另一方面可以揭示微衛(wèi)星在遺傳多樣性等領(lǐng)域的應(yīng)用[9-10]。因此,近年來基于全基因組開發(fā)微衛(wèi)星分子標(biāo)記受到研究者青睞。豬蛔蟲全基因組測序已完成,其基因組微衛(wèi)星分布規(guī)律也有報道[11-12]。本試驗基于已公布的豬蛔蟲全基因組數(shù)據(jù),設(shè)計微衛(wèi)星引物,驗證其多態(tài)性,并進(jìn)一步檢測其在近緣種人蛔蟲中的擴(kuò)增情況,以期為豬蛔蟲和人蛔蟲的種群遺傳研究、流行病學(xué)、宿主特異性、遺傳結(jié)構(gòu)、交配模式提供有價值的研究工具和分子標(biāo)記資源。
1 材料與方法
1.1 材料 豬蛔蟲采自撫州市孝橋屠宰場,收集了25頭豬的腸內(nèi)容物,共198條豬蛔蟲。
1.2 DNA提取和PCR擴(kuò)增 取豬蛔蟲樣本,剪下一小段體壁組織(約5 mm)用雙蒸水洗去固定液(2~3 次),剪碎,采用Wizard?SV Genomic DNA Purification System 試劑盒提取DNA。本試驗豬蛔蟲全基因組序列從美國國家生物技術(shù)信息中心NCBI(http://www.ncbi.nlm.nih.gov/)下載,然后使用軟件MSDB v2.4.1搜索并統(tǒng)計微衛(wèi)星序列,根據(jù)SSR側(cè)翼序列的保守性使用Primer Premier 5.0設(shè)計引物,引物長度18~22 bp,GC含量在40%~60%,目的片段長度在100~400 bp。總共設(shè)計了104對豬蛔蟲微衛(wèi)星引物,由生工生物工程(上海)股份有限公司合成。利用上述104對引物進(jìn)行PCR擴(kuò)增,擴(kuò)增體系為20 μL:PremixTaqVersion 2.0(TaKaRa)10 μL,上下游引物和DNA模板各2 μL,M-13FAM熒光標(biāo)記引物1.6 μL,加水至20 μL。反應(yīng)條件為94 ℃預(yù)變性5 min,先進(jìn)性5個循環(huán)的高溫PCR:94 ℃ 變性45 s,退火30 s,72 ℃延伸1 min,再進(jìn)行30個低溫循環(huán):94 ℃變性45 s,退火30 s,72 ℃ 延伸1 min,最后72 ℃延伸7 min,4 ℃保存。用1%瓊脂糖凝膠電泳分離PCR反應(yīng)產(chǎn)物。
1.3 引物的篩選 104對微衛(wèi)星引物均先用6個DNA樣本擴(kuò)增,能全部擴(kuò)增出條帶的引物即被初選。然后將初選得到的引物用于豬蛔蟲種群的擴(kuò)增(n=30),瓊脂糖凝膠電泳后,將有條帶的擴(kuò)增產(chǎn)物送至生工生物工程(上海)股份有限公司進(jìn)行STR掃描。并將所有有多態(tài)性的引物用人蛔蟲(n=6)擴(kuò)增。
1.4 數(shù)據(jù)處理 根據(jù)掃描結(jié)果,利用CERVUS V2.0計算等位基因數(shù)(Number of observed alleles,NA)、觀測雜合度(Observed heterozygosity,HO)、期望雜合度 (Expected heterozygosity,HE)和多態(tài)信息含量(Polymorphism information content,PIC),利用Micro-Checker ver. 2.2.3計算無效等位基因頻率(Frequency of null alleles,F)。利用Popgene1.32進(jìn)行近交系數(shù)(Inbreeding coefficient,FIS)和哈迪-溫伯格平衡(Hardy-Weinberg equilibrium,HWE)檢測。
2 結(jié)果
2.1 遺傳多態(tài)性分析 本試驗基于豬蛔蟲全基因組,根據(jù)微衛(wèi)星側(cè)翼序列的保守性共設(shè)計了104對引物,經(jīng)擴(kuò)增篩選共獲得38對能穩(wěn)定擴(kuò)增并具有清晰條帶的引物,經(jīng)再次篩選后擴(kuò)增成功的有28個位點,初步測試顯示所有位點也能在人蛔蟲中擴(kuò)增(表中數(shù)據(jù)未顯示)。在篩選到的28對引物中有25對具有多態(tài)性(表1),擴(kuò)增成功率26.9%。在25個有多態(tài)性的位點中,每個位點擴(kuò)增得到的等位基因數(shù)(NA)在2~25個,平均8.2個;觀測雜合度(HO)范圍為0~1,平均值0.442;期望雜合度(HE)范圍為0.042~0.945,平均值0.602;多態(tài)信息含量(PIC)范圍為0.040~0.921,平均值0.558,其中有15個微衛(wèi)星位點顯示高度多樣性(PIC>0.5),5個位點顯示中度多態(tài)性(0.5>PIC>0.25),其他位點多態(tài)性較低(PIC<0.25)(表1);無效等位基因頻率范圍為0~0.293 3,ASM46、ASM132、ASM213、ASM252、ASM535、ASM551、ASM573、ASM597、ASM598、ASM615、ASM632、ASM650這12個位點沒有出現(xiàn)無效等位基因,其他位點均存在無效等位基因。
2.2 哈迪-溫伯格平衡檢測 Hardy-Weinberg檢測顯示,篩選獲得的25個多態(tài)性位點中有15個符合哈迪-溫伯格平衡(P>0.05),有10個位點或多或少偏離哈迪-溫伯格平衡(圖1)。此外,通過Popgene1.32計算的FIS變化區(qū)間為-0.198 5~1,其中21個位點FIS>0(圖1)。
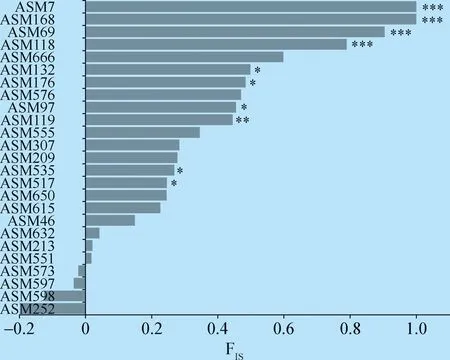
圖1 豬蛔蟲微衛(wèi)星位點的FIS值統(tǒng)計Fig.1 FIS value of microsatellite loci in A.suum注:負(fù)值表示雜合子過剩, 正值表示純合子過量; *:偏離哈迪-溫伯格平衡的位點(*:0.01 HWE檢測顯示,多數(shù)位點偏離哈迪-溫伯格平衡。這與人蛔蟲和Nicrophorusvespilloides開發(fā)的微衛(wèi)星分子標(biāo)記類似[6,13],同時烏干達(dá)西南部人蛔蟲的遺傳多樣性研究也出現(xiàn)了這種情況[14],這很可能是蛔蟲種群的普遍現(xiàn)象。而造成這種結(jié)果的原因可能是由于這些位點的純合子過量或近親繁殖,但也可能是由于這些標(biāo)記物中存在無效等位基因引起的,此次試驗也發(fā)現(xiàn)無效等位基因確實存在,而且無效等位基因存在的多數(shù)位點偏離HWE,其中ASM7、ASM69、ASM118、ASM168這4個位點顯著偏離HWE(P≤0.001),同時這些位點無效等位基因頻率相比于其他位點也較高。由此推斷無效等位基因的存在可能是導(dǎo)致偏離HWE的原因之一。 此外,F(xiàn)IS檢測顯示21個位點FIS>0,且多數(shù)位點觀測雜合度小于期望雜合度,這與人蛔蟲及其他線蟲相似[6,15],表現(xiàn)為雜合子缺失,種群個體間親緣關(guān)系較近。試驗結(jié)果表明,多數(shù)位點雜合子缺失,這種現(xiàn)象可能是由宿主內(nèi)個體近交和/或Wahlund效應(yīng)造成。而Criscione等在人蛔蟲遺傳多樣性分析中也發(fā)現(xiàn)了雜合子缺失的多數(shù)位點也偏離哈迪-溫伯格平衡,并解釋這種現(xiàn)象可能是由于Wahlund效應(yīng),而非宿主內(nèi)隨機(jī)交配[6]。綜上所述,本試驗認(rèn)為多數(shù)位點偏離哈迪-溫伯格平衡可能是由無效等位基因和/或雜合子缺失造成的。 本試驗基于已公布的豬蛔蟲全基因組,進(jìn)行了豬蛔蟲微衛(wèi)星分子標(biāo)記的開發(fā)和特征分析,篩選獲得了25個能穩(wěn)定擴(kuò)增并具有較高多態(tài)性的微衛(wèi)星位點。這些標(biāo)記可用于蛔蟲遺傳多樣性、遺傳結(jié)構(gòu)、譜系鑒定和種間雜交等的研究。 表1 豬蛔蟲25個微衛(wèi)星位點特征描述Table 1 Characteristics of the 25 microsatellite loci for A. suum 續(xù)表13 討論


