酸脅迫處理對鼠傷寒沙門氏菌抗酸性的影響
2021-01-20 06:50:00李琳瓊張愛靜王鵬杰高瑀瓏
食品科學 2021年1期
李琳瓊,洪 靜,張愛靜,王鵬杰,高瑀瓏
(南京財經大學食品科學與工程學院,江蘇省現代糧食流通與安全協同創新中心,江蘇高校糧油質量安全控制及深加工重點實驗室,江蘇 南京 210023)
沙門氏菌是一種常見的食源性致病菌,在自然界分布廣泛、種類繁多,幾乎所有的血清型沙門氏菌都具有致病性,在全球范圍內,由其引起的細菌性食物中毒常居食源性疾病首位[1-2]。沙門氏菌不僅威脅到人類健康,而且給食品制造業帶來巨大損失[3-5]。沙門氏菌廣泛寄生于動物腸道內,常常存在于新鮮的家禽肉和豬肉等肉制品中,能引起敗血癥和腸胃炎。在食品生產、貯藏、運輸和銷售過程中,沙門氏菌常遭受極端的酸、堿、滲透壓、溫度、壓力以及抗菌劑等不適生長條件的脅迫作用,誘發其產生一系列的適應性來應對變化的環境條件,這種適應性包含同源性保護作用和交叉性保護作用[6-7]:如李斯特菌經某種不利生存的因素長時間脅迫后,其生存能力增強,這種現象被Hill等[8]稱為同源性保護作用;沙門氏菌遭受某種不利生存的環境脅迫后,之后在其他不利環境條件下其生存能力增強,這種現象被He Shoukui等[9]稱為交叉性保護作用。這些應激反應(適應性反應)均有助于菌體的損傷修復,能夠增強其抵抗后續應激作用的能力。
環境脅迫是指環境因素接近或超過生物體、種群或群落的忍耐極限時造成的脅迫作用[10]。酸對微生物的脅迫作用一直是生物學領域研究的熱點,而微生物的抗酸性受其種類、生長階段和環境因素的影響[11]。在食品生產加工的過程中,微生物常遭受各種不適環境條件的脅迫,誘導出一系列自我保護機制,導致其對致死因素的抗性增加。其中,pH值是引起細菌細胞組成和生理變化的一個重要因素[12]。
在食品保藏過程中,有許多關于采用有機酸來控制、殺滅食源致病菌的研究報道,例如采用一定濃度的有機酸對鮮肉和蔬果等進行噴淋、浸泡等處理來殺滅其中的致病菌[13-14]。Foster等[15]首次提出誘導性耐酸應答的概念,認為在酸脅迫條件下處于亞致死狀態的細菌經過損傷修復之后,其生存能力會明顯增強。Zhang Yimin等[16]將單核增生李斯特菌置于添加了乳酸、醋酸和乙酰丙酸酸化的培養基中,發現其生長至穩定期后,再經過一定致死條件的膽鹽和酸處理后,其滅活率低于原野生對照菌株,說明單核增生李斯特菌經過酸處理后產生了抗酸性。Cebrián等[17]通過對金黃色葡萄球菌進行亞致死濃度的酸、堿、熱和H2O2處理后,發現其對這些脅迫因子的抗性與原野生對照菌株相比顯著增加。Martine等[18]研究發現,大腸桿菌受到酸性環境的脅迫作用后,其生理狀態會發生相應的變化,其中調控因子基因RpoS表達量上調并產生一定量的酸修復蛋白。但對于鼠傷寒沙門氏菌(Salmonella typhimurium)經不同種類和不同pH值的有機酸脅迫后,是否可誘導其產生抗酸性和同源性保護作用,目前尚不清楚。
食品中微生物在不適環境條件下的生長和失活可通過建立微生物預測模型來對其進行預測,為食品中微生物的控制提供科學的理論依據[19]。常用來描述微生物生長和失活的動力學模型有Linear模型[20]、Logistic模型[21]、Gompertz模型[22]、Baranyi模型[23]和Weibull模型[24]等。Linear模型操作簡便,但對數據預測的精確度不高;Logistic模型是數學中用來描述某一普通笛卡爾平面上S型曲線的模型,有人也將其用來研究細菌生長曲線,但因其不能擬合細菌延滯期的生長變化,所以不能完整地顯示S型曲線;Gompertz模型和Baranyi模型克服了這些缺點;Baranyi模型中的參數生物學意義更明確,能夠較好地描述微生物的生長曲線,目前應用較多。Juneja等[25]使用Baranyi模型、修正Gompertz模型和Huang模型等分別建立了蠟樣芽孢桿菌在四季豆中的一級生長模型,發現Baranyi模型為其最適生長模型;Weibull模型在材料工業中常用于對脆性材料強度的壽命預測,也可用于研究不同實驗條件對細菌失活的影響,適用于熱力殺菌[26]和非熱力殺菌[27]條件下細菌失活模型的擬合。張愛靜等[28]發現采用Weibull模型能較好地擬合經多次熱脅迫后產生抗性的大腸桿菌在80 ℃下的熱失活曲線。
本實驗采用食品工業中常用的具有較強殺菌能力的檸檬酸、乙酸、乳酸和蘋果酸等有機酸處理S.typhimurium,借助Weibull模型研究經不同有機酸誘導后其抗酸性的變化,以期為有機酸殺菌技術的廣泛應用提供理論參考,對保障食品加工貯藏安全、有效控制食源性致病菌、建立食品質量安全控制體系具有重要意義。
1 材料與方法
1.1 菌株、材料與試劑
鼠傷寒沙門氏菌(S.typhimurium)CGMCC 1.1190購于中國普通微生物菌種保藏管理中心。
胰蛋白胨大豆瓊脂(trypticase soy agar,TSA)培養基、胰蛋白胨大豆肉湯(trypticase soy broth,TSB)液體培養基、檸檬酸、乙酸、乳酸、蘋果酸、鹽酸、氯化鈉(均為分析純) 南京丁貝生物科技有限公司。
1.2 儀器與設備
LDZX-50FBS型立式壓力蒸汽滅菌器 上海申安醫療器械廠;GNP-9160型隔水式恒溫培養箱、SF-CF-2A型超凈工作臺 上海三發科學儀器有限公司;PHS-3G型pH計 上海儀電科學儀器股份有限公司;HZQ-F160型恒溫振蕩培養箱 太倉市實驗設備廠;ME55型精密電子天平 北京塞多斯天平有限公司;SK-1型快速混勻器金壇市杰瑞爾電器有限公司;A1正置熒光顯微鏡 德國蔡司公司。
1.3 方法
1.3.1 菌種活化與預處理
供試菌經活化后,接入pH 7.2的TSB培養基,于37 ℃、150 r/min搖床振蕩培養24 h,后續作為對照組,備用。
1.3.2 CGMCC 1.1190抗酸菌株的獲得
不同pH值TSB培養基的配制:分別用濃度0.05 mol/L檸檬酸(pH 2.2)、1 mol/L醋酸(pH 2.4)、1.7 mol/L乳酸(pH 2.5)、0.07 mol/L蘋果酸(pH 2.4)調節TSB培養基的pH值,得到12 種pH值分別為6.2、5.2和4.2的TSB培養基,121 ℃、20 min滅菌,冷藏備用。強酸基質培養基的配制:用濃度1 mol/L HCl溶液調節TSB培養基至pH 2.5,滅菌備用。
酸脅迫處理:將材料中經活化的CGMCC 1.1190菌懸液以體積分數1%轉接至上述12 種不同pH值的TSB培養基中,搖床振蕩培養(37 ℃、150 r/min)。對不同pH值的TSB培養基中CGMCC 1.1190菌株定期取樣進行菌落計數,不同pH值TSB培養基選取不同時間間隔進行取樣,時間間隔的選取根據CGMCC 1.1190菌株在不同pH值TSB培養基中的生長情況決定,其生長情況通過分光光度計測定的光密度確定。按文獻[29]的方法使用TSA平板計數法進行菌落計數。不同pH值的TSB培養基中CGMCC 1.1190菌株的生長量通過計算每毫升TSB培養基中活菌的數量(CFU/mL)確定,以(lg(CFU/mL))表示。
抗酸菌株的獲得:取1.3.1節中pH 7.2的TSB培養基與上述12 種pH值分別為6.2、5.2、4.2的TSB培養基中培養至穩定期的CGMCC 1.1190菌株培養液10 mL混合,離心(4 ℃、5 000 r/min,10 min),棄上清液,收集的沉淀用質量分數0.85%無菌生理鹽水洗滌兩次,再以等體積的生理鹽水制成菌懸液,分裝于無菌離心管,備用。
1.3.3 生長模型的建立
采用Baranyi模型擬合CGMCC 1.1190菌株在不同種類、不同pH值的酸脅迫環境下的生長曲線,建立CGMCC 1.1190菌株在不同pH值的TSB培養基中生長的一級模型,菌株的生長量參照公式(1)計算。
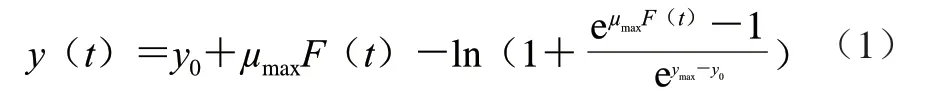
式中:t為培養時間/h;y(t)為t時刻CGMCC 1.1190菌株的生長量(lg(CFU/mL));y0為t=0時CGMCC 1.1190菌株初始菌數的對數(lg(CFU/mL));ymax為菌體最大生長量的對數(lg(CFU/mL));μmax為最大比生長速率/h-1;F(t)為調整函數,表示CGMCC 1.1190菌株在生長過程中,從遲滯期到對數期的過渡時期(調整生理狀況和對新環境的適應期)所對應的生長遲滯時間。
F(t)按照公式(2)、(3)計算。
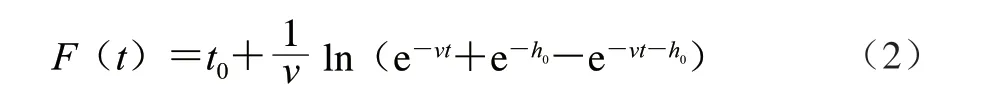
式中:v為菌體在生長過程中的增長速率/h-1,相當于μmax;h0為菌體的初始生理狀態參數,用來描述其適應新環境的能力。

式中:λ為遲滯時間/h。
1.3.4 生長模型的驗證
生長模型的驗證采用決定系數R2、準確因子Af和偏差因子Bf。其中,R2表示模型的精確度和可靠性,R2越接近1,模型的擬合度越高;Af是衡量預測值與實測值之間的接近程度,Af越接近1,模型的擬合度越好、越精確;Bf>1表示模型預測值比實測值高,Bf<1表示模型預測值比實測值低,Bf越接近1,模型擬合度越高[30]。
CGMCC 1.1190菌株在經檸檬酸、乳酸、醋酸和蘋果酸調節pH值分別至6.2、5.2和4.2的TSB培養基中生長,在每種pH值條件下,在不同于模型建立的時間點進行取樣驗證,每次實驗重復3 次。R2、Af和Bf分別按照公式(4)~(6)計算。

式中:n為測定值的個數。
1.3.5 CGMCC 1.1190菌株抗酸性的測定
利用Weibull模型來擬合CGMCC 1.1190菌株在pH 2.5的TSB培養基中的酸致死曲線,Weibull模型的規模參數δ/min和形態參數ρ按公式(7)計算。

式中:N0為初始CGMCC 1.1190菌數量;Nt為酸處理t時刻后存活的CGMCC 1.1190菌數量;t為酸處理時間/min。
將1.3.1節中制備的對照組和酸脅迫CGMCC 1.1190菌株生理鹽水的菌懸液分別離心(4 ℃、5 000 r/min,10 min),棄上清液,重懸于10 mL的pH 2.5 TSB培養基中進行酸處理,每隔10 min取樣,于TSA培養基37 ℃培養72 h后,進行菌落計數,其存活率按公式(7)計算。利用Weibull模型對酸脅迫培養的CGMCC 1.1190菌株在pH 2.5的TSB培養基中的酸致死曲線進行非線性擬合。
1.3.6 菌體形態的觀察
對1.3.1 節p H 7.2 的T S B 培 養 基 中 培 養 的CGMCC 1.1190原始對照菌種和分別經pH 6.2、5.2和p H 4.2 的檸檬酸脅迫培養至穩定期獲得的3 種CGMCC 1.1190抗酸性菌株,在TSA培養基培養48 h后,以無菌接種環挑取單菌落,用結晶紫和碘液簡單染色,顯微鏡觀察CGMCC 1.1190菌株的個體形態。
1.4 數據處理與分析
實驗數據通過DMFit(www.combase.cc)軟件和JMP 10.0軟件進行模型擬合,采用SPSS 22.0軟件進行數據處理,實驗數據以平均值±標準差表示,差異性顯著分析采用Duncan's新復極差法,P<0.05表示差異顯著。
2 結果與分析
2.1 CGMCC 1.1190菌株在不同pH值TSB培養基中的生長曲線
CGMCC 1.1190菌株在pH值分別為6.2、5.2和4.2 TSB培養基中的Baranyi模型生長曲線及其失活情況見圖1。
除了經乳酸、醋酸和蘋果酸調節的pH 4.2的TSB培養基外,CGMCC 1.1190菌株在其他幾種不同pH值TSB培養基中的生長量皆均勻地分布在生長曲線兩側,說明利用Baranyi模型能很好地擬合CGMCC 1.1190菌株在pH 7.2和有機酸酸化處理后不同pH值TSB培養基中的生長曲線,CGMCC 1.1190菌株的生長曲線均呈S型,擬合度較高(R2>0.995),都有典型的延滯期、對數期和穩定期,pH值越低,遲滯期越長。在pH 4.2檸檬酸酸化處理的TSB培養基中,CGMCC 1.1190菌株的生長也呈典型的S型生長曲線;而在經乳酸、醋酸和蘋果酸3 種有機酸酸化處理pH 4.2的TSB培養基中,隨著培養時間的延長,CGMCC 1.1190菌體數量快速降低,分別在12、10 h和8 h后低于檢測限,推斷其在經乳酸、醋酸和蘋果酸3 種有機酸酸化處理pH 4.2的TSB培養基中不生長并開始失活。與醋酸、乳酸和蘋果酸相比,CGMCC 1.1190菌株在檸檬酸酸化的TSB培養基中的生存能力較強,對檸檬酸的耐受性、抗酸性作用最強。
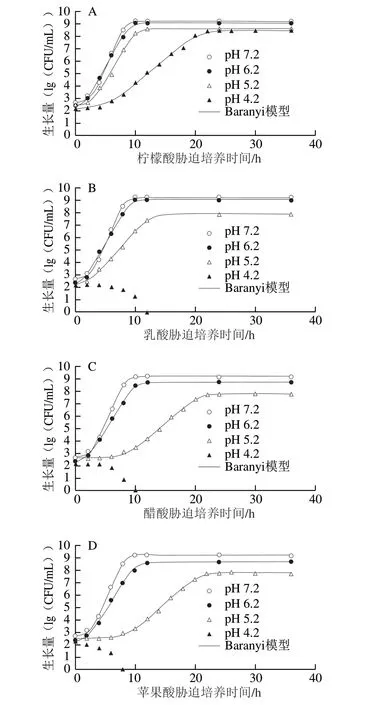
圖1 S.typhimurium CGMCC 1.1190菌株在不同pH值TSB培養基中的生長曲線及其失活情況Fig.1 Growth curves and inactivation of S.typhimurium CGMCC 1.1190 in TSB acidified at different pH
在不同pH值的TSB培養基中,CGMCC 1.1190菌株生長的一級Baranyi模型曲線的參數估計值見表1。
Baranyi模型是近年來運用廣泛的一種微生物模型[31],其既表達了微生物生長量與時間的關系,也較為準確地評估微生物λ、μmax及ymax等生長參數[32]。微生物生長的遲滯期是指微生物接種到新的培養基后,由于要適應新環境,微生物數目不會發生變化的一段時間。由表1可知,在CGMCC 1.1190菌株的生長曲線中,對于同一種有機酸酸化處理的TSB培養基,pH值越低,λ越長。CGMCC 1.1190菌株生長的遲滯時間與有機酸種類有關,在pH 4.2檸檬酸酸化處理的TSB培養基中,CGMCC 1.1190菌株的生長有典型的S型生長曲線(圖1A),λ為5.32 h,而在經乳酸、醋酸和蘋果酸酸化處理的pH 4.2的TSB培養基中,隨著時間的延長,CGMCC 1.1190菌株的數量明顯降低,表現為不生長、被抑制而失活,可能是因為CGMCC 1.1190菌體的核酸、蛋白質、細胞膜等的生物大分子物質對這3 種酸較敏感,在酸脅迫下發生了變性,細胞的生理代謝功能急劇下降,造成菌體死亡。說明CGMCC 1.1190菌株抗酸性的強弱與有機酸的種類有關,其對檸檬酸的抗性大于乳酸、醋酸和蘋果酸。
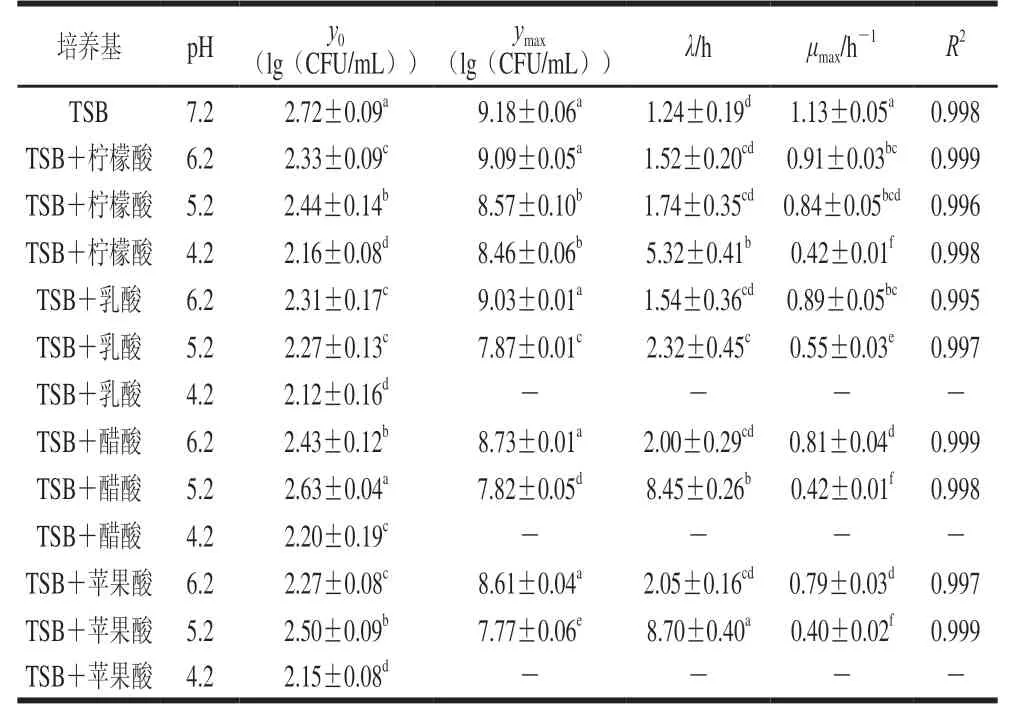
表1 S.typhimurium CGMCC 1.1190菌株生長的一級Baranyi模型曲線的參數估計值Table 1 Parameter estimates of Baranyi primary model for the growth of S.typhimurium CGMCC 1.1190
在經pH 5.2的乳酸、醋酸和蘋果酸酸化處理的TSB培養基中,CGMCC 1.1190菌株生長的λ分別為2.32、8.45 h和8.70 h,皆明顯長于經檸檬酸酸化處理pH 5.2的TSB培養基中λ(1.7 h);在經醋酸和蘋果酸酸化處理的pH 6.2的TSB培養基中,λ分別為2.00 h和2.05 h,明顯長于在經檸檬酸酸化處理的pH 6.2的TSB培養基中的λ(1.5 h)。研究結果表明,在相同pH值條件下,CGMCC 1.1190菌株對檸檬酸產生的抗性最強。
由圖1 與表1 可知, 不同有機酸脅迫培養CGMCC 1.1190菌株的對數期差異顯著(P<0.05),在經pH 5.2檸檬酸酸化處理后,CGMCC 1.1190菌株生長的對數期約為8 h,在經pH 5.2的乳酸、醋酸和蘋果酸酸化處理后,對數期分別為10、16 h和16 h,表明CGMCC 1.1190菌株在不同種類的酸脅迫下的生長速率不同,檸檬酸脅迫生長速率最快,醋酸和蘋果酸脅迫生長速率最慢;CGMCC 1.1190菌株的μmax也反映了菌體生長過程中對不適環境抵抗能力的強弱,μmax越大,抵抗能力越強,在pH 5.2的檸檬酸、乳酸、醋酸和蘋果酸酸化處理的TSB培養基中,μmax分別為0.84、0.55、0.42 h-1和0.40 h-1,說明CGMCC 1.1190菌株對這4 種酸的抵抗能力依次減小,其中,經檸檬酸酸化處理的TSB培養基中μmax顯著大于在其他3 種酸化處理的μmax(P<0.05),也說明同一pH值下CGMCC 1.1190菌株在檸檬酸酸化處理的TSB培養基中抗性最強。
在4 種酸脅迫條件下,y0在2.12~2.72(lg(CFU/mL))之間,彼此相差小于1(lg(CFU/mL))。進入穩定期后,在pH 7.2和4 種有機酸調節至pH 6.2的TSB培養基中ymax均可達到108~109CFU/mL,無顯著性差異(P>0.05),表明在pH 6.2時,CGMCC 1.1190菌株生長所能達到的最大菌濃度并不受有機酸種類的影響。在以4 種酸分別調節至pH 5.2的TSB培養基中,CGMCC 1.1190菌株的ymax分別為8.57、7.87、7.82、7.77(lg(CFU/mL)),呈顯著降低趨勢(P<0.05);當檸檬酸調節pH 4.2時,ymax為8.46(lg(CFU/mL)),顯著高于在經乳酸、醋酸和蘋果酸酸化處理pH 5.2 TSB培養基中的ymax(P<0.05)。結果表明,CGMCC 1.1190菌株抗酸性大小與有機酸的種類相關,其中檸檬酸>乳酸>醋酸>蘋果酸。
2.2 一級Baranyi生長模型的驗證結果
CGMCC 1.1190菌株一級Baranyi模型的驗證結果見表2。對CGMCC 1.1190菌株生長的實測值及一級模型的預測值進行相關性分析,決定系數R2=0.999 7、Af=1.008 3、Bf=1.005 3,表明本實驗建立的Baranyi模型擬合效果好,且有效、可靠,可以利用Baranyi模型對酸脅迫培養的CGMCC 1.1190菌株的生長情況進行預測。
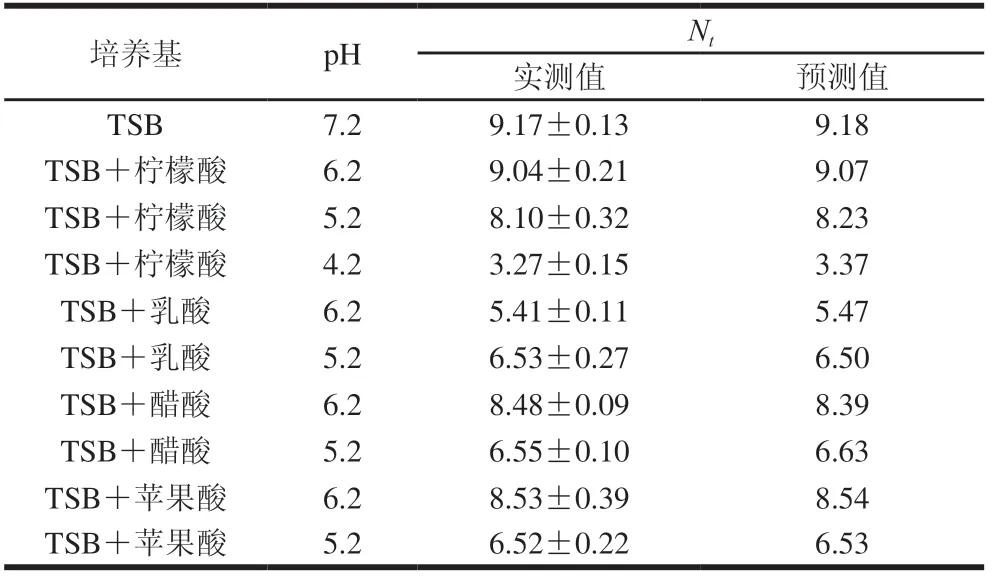
表2 S.typhimurium CGMCC 1.1190菌株生長一級Baranyi模型的驗證Table 2 Validation of Baranyi model for S.typhimurium CGMCC 1.1190
2.3 抗酸性菌株在pH 2.5 TSB培養基中的酸致死Weibull模型曲線
1.3.2 節中經檸檬酸、乳酸、醋酸和蘋果酸4 種有機酸調節pH值分別至6.2、5.2的TSB培養基中脅迫培養獲得的8 種抗酸性CGMCC 1.1190菌株與pH 7.2的TSB培養基中培養的對照組CGMCC 1.1190菌株,在pH 2.5 TSB培養基中的酸致死曲線分別見圖2、3。
由圖2 可知,采用W e i b u l l 模型能較好地擬合CGMCC 1.1190菌株經4 種有機酸脅迫培養后獲得的抗酸菌株(pH 6.2)在pH 2.5 TSB培養基中的失活曲線,擬合程度較高(R2>0.991 0)。CGMCC 1.1190菌株經酸脅迫培養后,再置于pH 2.5的強酸性環境下,隨著酸處理時間的延長,菌體存活率均呈下降趨勢,但與對照組相比,其存活率明顯增大,表明經4 種有機酸酸脅迫(pH 6.2)培養至穩定期的4 種CGMCC 1.1190酸脅迫菌株之間抗酸性存在差異,說明對酸的抗性作用與脅迫培養的有機酸的種類相關,其中脅迫菌株抗酸性從大到小依次為檸檬酸>乳酸>醋酸>蘋果酸。
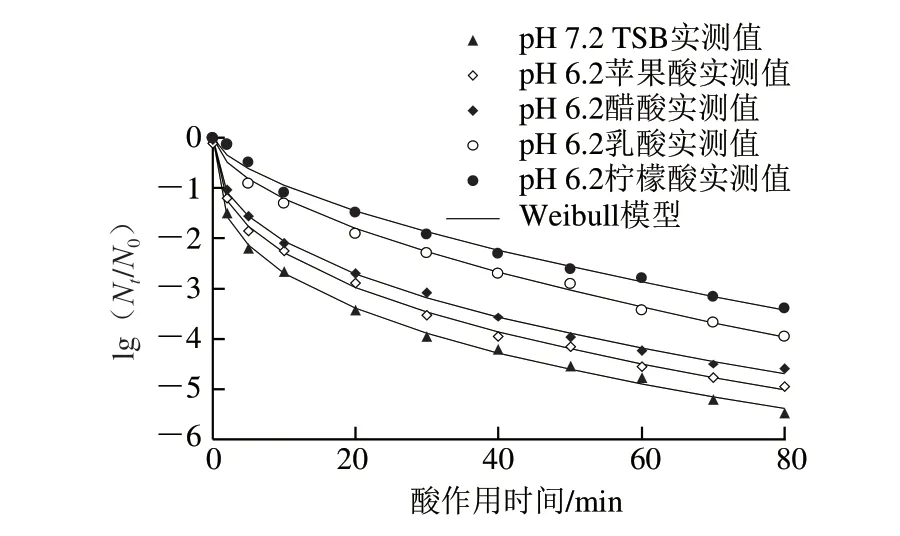
圖2 pH 6.2的TSB培養基中脅迫培養獲得的S.typhimurium CGMCC 1.1190抗酸菌株在pH 2.5的TSB培養基中的致死曲線Fig.2 Inactivation curves of S.typhimurium CGMCC 1.1190 cultured in TSB at pH 6.2 when exposed to pH 2.5
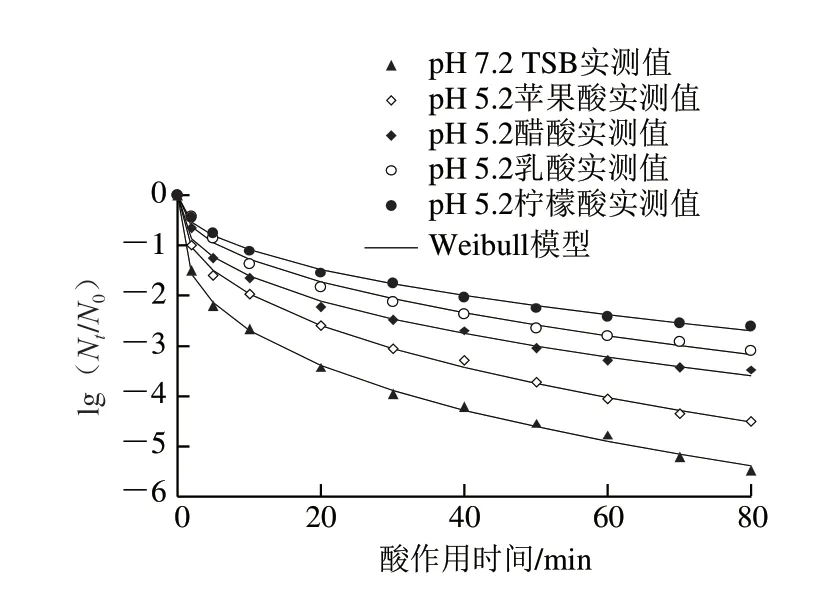
圖3 pH5.2的TSB培養基中脅迫培養獲得的S.typhimurium CGMCC 1.1190抗酸菌株在pH 2.5的TSB培養基中的致死曲線Fig.3 Inactivation curves of S.typhimurium CGMCC 1.1190 cultured in TSB at pH 5.2 when exposed to pH 2.5
由圖3可知,采用Weibull模型也能較好地擬合CGMCC 1.1190菌株經4 種有機酸脅迫培養后獲得的抗酸菌株(pH 5.2)在pH 2.5 TSB培養基中的失活曲線,擬合程度較高(R2>0.992 6)。研究發現,經4 種有機酸酸脅迫培養至穩定期的4 種CGMCC 1.1190酸脅迫菌株抗酸性存在差異,這4 種CGMCC 1.1190酸脅迫菌株(pH 5.2)對pH 2.5的強酸的抗性大小也與有機酸的種類相關,其中脅迫菌株抗酸性從大到小依次為檸檬酸>乳酸>醋酸>蘋果酸。
利用Weibull模型對本實驗獲得的9 種CGMCC 1.1190抗酸性菌株與對照組在pH 2.5的TSB培養基中的致死實驗數據進行回歸擬合,以規模參數δ來評價CGMCC 1.1190菌株的抗酸性,通過JMP軟件內部自動運算所得的模型參數δ和ρ見表3。
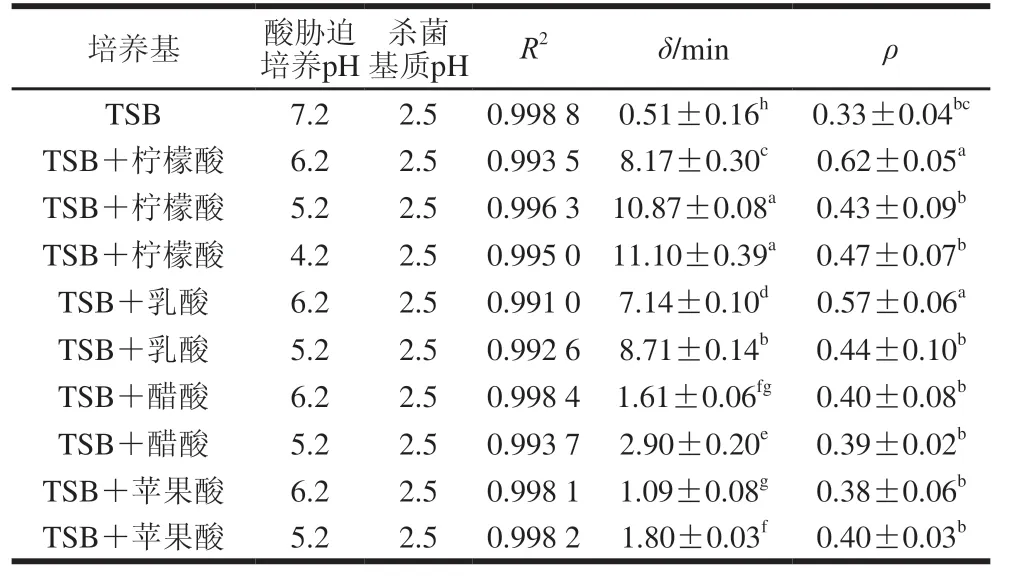
表3 S.typhimurium CGMCC 1.1190抗酸性菌株在pH 2.5的TSB培養基中酸致死模型曲線的擬合度及參數Table 3 Fitness and parameters of inactivation models for S.typhimurium CGMCC 1.1190 cultured in medium acidified with different organic acids when exposed to pH 2.5
ρ參數代表以Weibull模型擬合CGMCC 1.1190菌株的酸致死曲線的形狀,當ρ=1時,曲線為直線;當ρ>1時,曲線向下彎曲,為凸型曲線;當ρ<1時,曲線向上彎曲,為凹型曲線[33]。δ表示CGMCC 1.1190菌體數量第一次減少10 倍所用的時間/min,抗酸性大小與δ成正比。隨著脅迫培養的pH值的降低,CGMCC 1.1190菌株的抗酸性呈上升趨勢,用檸檬酸脅迫培養pH值分別為6.2、5.2、4.2 TSB培養基中的CGMCC 1.1190菌株,δ分別為8.17、10.87 min和11.10 min,顯著大于對照組pH 7.2 TSB培養基中CGMCC 1.1190菌株的δ(0.51 min)(P<0.05)。同樣,經乳酸、醋酸和蘋果酸3 種酸脅迫培養后,CGMCC 1.1190菌株的δ均呈現相同的變化趨勢。pH 6.2和pH 5.2檸檬酸脅培養的δ分別是蘋果酸脅迫培養的7.5 倍和6.0 倍,也表明在相同pH值下檸檬酸脅迫培養誘導CGMCC 1.1190菌株產生的抗酸性最強,蘋果酸脅迫培養誘導產生的抗酸性最弱。經過酸脅迫培養后的CGMCC 1.1190菌株比對照組菌種更加耐酸,對照組菌種于pH 2.5的TSB培養基中處理80 min后菌體數量下降約5.5 個對數,大部分菌體死亡,經檸檬酸調節pH 4.2 TSB培養基中脅迫誘導培養的CGMCC 1.1190菌株于pH 2.5 TSB培養基中處理80 min后,菌體數量僅下降了約2.4 個對數,CGMCC 1.1190菌株的存活率明顯提高。實驗結果ρ<1,表明CGMCC 1.1190菌株的酸致死曲線向上彎曲,為凹型曲線,進一步說明模型曲線隨著酸處理時間的延長,其下降趨勢減緩,菌株失活率下降趨勢減緩。綜上,以4 種有機酸脅迫培養CGMCC 1.1190菌株至穩定期均可誘導其產生抗酸性,在強酸作用下其致死率降低,抗酸性強弱與有機酸種類和pH值有關,對于同一種有機酸,pH值越低,脅迫誘導CGMCC 1.1190菌株產生的抗酸性越強。
2.4 檸檬酸脅迫前后CGMCC 1.1190菌株個體形態的變化
CGMCC 1.1190菌株原始對照菌種與分別經過pH 6.2、5.2和pH 4.2檸檬酸脅迫培養至穩定期獲得的3 種耐酸菌株在顯微鏡下的個體形態見圖4。
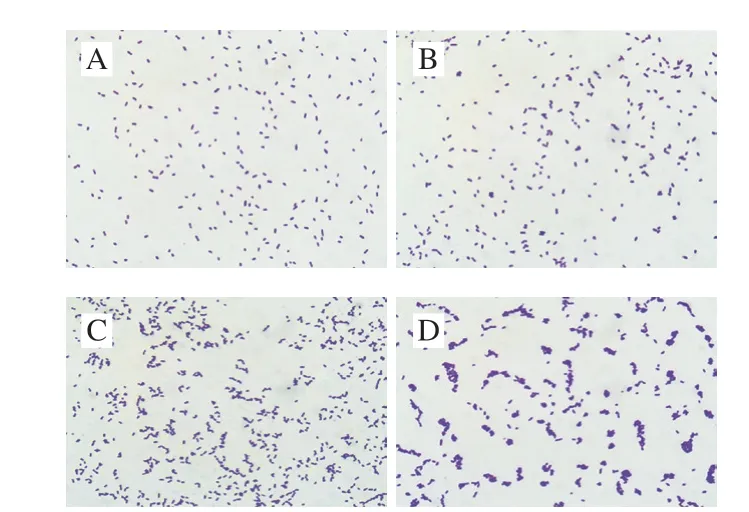
圖4 S.typhimuriumCGMCC 1.1190菌株個體形態圖(×1 000)Fig.4 Cellular morphology of S.typhimurium CGMCC 1.1190 (× 1 000)
CGMCC 1.1190對照菌株酸脅迫培養48 h后,在顯微鏡下呈單個分散狀態的短桿狀,單桿菌兩端為鈍圓(圖4A);經pH 6.2的檸檬酸脅迫培養后,個體形態略有變化,有極少數個體聚集并呈雙桿狀或八字形排列(圖4B);經pH 5.2的檸檬酸脅迫培養后,大部分菌體聚集并呈鏈狀或八字形排列(圖4C);經pH 4.2的檸檬酸脅迫培養后,幾乎全部菌體聚集并成鏈狀、柵狀或重疊狀排列(圖4D)。說明檸檬酸的脅迫pH值越低,CGMCC 1.1190菌體的聚集狀態越明顯。個體間聚集程度的變化有助于其避免在強酸環境下產生損傷,這可能與其抗酸性能力的增加有一定的關系。劉艷霞等[34]研究發現,用某些藥物對雙歧桿菌作用后,其個體出現絲狀體生長現象,這種個體形態的適應性變化有利于其抵抗不適的外界環境。目前,對于CGMCC 1.1190菌株酸脅迫導致其抗酸能力增加的相關機理仍在研究。本實驗發現酸脅迫培養對S.typhimuriumCGMCC 1.1190菌株產生了同源性保護作用。
3 結 論
本實驗研究了S.typhimuriumCGMCC 1.1190菌株分別在經檸檬酸、乳酸、醋酸和蘋果酸調節pH值的TSB培養基中脅迫培養,結果表明,在相同pH值條件下CGMCC 1.1190菌株的抗酸性從大到小依次為檸檬酸>乳酸>醋酸>蘋果酸。篩選出經脅迫培養的耐酸性菌株,發現在相同pH值下脅迫誘導,檸檬酸脅迫菌株抗酸性最強;對于同一種有機酸,pH值越低,脅迫培養誘導CGMCC 1.1190菌株產生的抗酸性越強,個體形態聚集程度越明顯。綜上,一定濃度的有機酸前期脅迫培養有利于保護CGMCC 1.1190菌株后期對強酸處理的同源性保護作用。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
光學精密工程(2016年6期)2016-11-07 09:07:19
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
