棉鈴蟲Ⅰ型和Ⅶ型幾丁質酶基因表達規律的比較分析
2021-01-18 07:51:14羅生慧麥麥提艾力阿卜杜納斯爾白雪瑩呂國會劉小寧
環境昆蟲學報 2020年6期
羅生慧,隨 慧,包 靜,麥麥提艾力·阿卜杜納斯爾, 白雪瑩,呂國會,劉小寧
(新疆大學生命科學與技術學院,新疆生物資源基因工程重點實驗室,烏魯木齊 830046)
棉鈴蟲是主要的農業害蟲之一,是棉花蕾鈴期的主要害蟲,對其它農作物也具有巨大的危害性。因其具有廣泛寄主范圍以及轉移危害的習性,對農業經濟造成巨大的損失。化學殺蟲劑因為使用方便、作用快速而被作為主要控制害蟲的方法,但長期使用化學藥物導致抗藥性增加(Aktaretal., 2009)。因此,需要尋找一種環境友好型的害蟲防治方法。研究發現,幾丁質是構成昆蟲角質層、表皮、腸道及圍食膜的主要成分,而高等動植物體內不存在幾丁質,因此,可以通過幾丁質降解酶來破壞棉鈴蟲中幾丁質的代謝平衡,從而可以達到綠色防治棉鈴蟲的目的。
幾丁質(Chitin)是昆蟲外骨骼角質層、圍食膜、頭殼、體壁等部位的主要成分,用以保護昆蟲并維持昆蟲的體型,還可以限制昆蟲的生長。幾丁質是在昆蟲內表皮和外表皮或在新分泌的、非硬化的前表皮中發現的,且在昆蟲的整個生命過程中幾丁質的降解與合成是一直存在的(Lehane, 1997; 張嚴峻等,2000;相靜波,2004;張八生,2007;Hegedusetal., 2009)。在昆蟲的整個生命周期中,昆蟲必須降解舊的角質層同時合成新角質(Hataetal., 1996; Merzendorfer and Zimoch, 2003; Muthukrishnanetal., 2012),即蛻皮現象(Nijhoutetal., 1981)。而昆蟲的蛻皮過程是由幾丁質酶(chitinase)和β-N-乙酰葡糖胺糖苷酶(β-N-acetyglucosaminidase)協同完成的(Fukamizoetal., 1985)。
幾丁質酶(chitinase, CHT)是昆蟲生長發育的各個階段起著至關重要的作用,在蛻皮、圍食膜的降解以及消化等重要生理過程(Shietal., 2004; Zhuetal., 2008; Arakaneetal., 2010; Jasrapuriaetal., 2010; Quetal., 2014)均與幾丁質酶息息相關。幾丁質酶是一類具生物催化活性的水解酶,能特異性水解成形的幾丁質。隨著生物技術及生物信息學的不斷發展,通過對昆蟲幾丁質酶及其類似蛋白的催化域進行系統發育分析,可將昆蟲幾丁質酶劃分為十個(Ⅰ-Ⅹ)類型(group)(Arakaneetal., 2003; Zhangetal., 2011; 夏夢,2015;Liuetal., 2018),每個類型幾丁質酶在昆蟲發育過程中執行不同的生理功能,其中II型幾丁質酶包括4~5個催化域和4~7個CBD區主要與蛻皮發育相關(李大琪等,2011);III型幾丁質酶包含2個催化域和一個CBD區,在N端含有一個跨膜結構域,屬于膜結合蛋白,參與昆蟲蛹化后腹部的收縮和翅的伸展(范曉軍等,2013);IV型幾丁質酶主要由一個信號肽和一個催化域組成,大部分缺失CBD區,參與圍食膜的降解與更新,抵抗含有幾丁質的有害病原菌,對昆蟲起免疫保護作用(Khajuriaetal., 2010);V型幾丁質酶包含一個信號肽和一個催化域,但缺乏CBD區,可作為生長因子,來增強細胞的增值能力(Zhuetal., 2008);VI型幾丁質酶分子量較大,在C端有一個較長的富含Ser/Thr的高度糖基化無規則卷曲,可用來保護相似結構的幾丁質酶不被降解(Yasuyuki and Subbaratnam, 2010)。本文主要對I型幾丁質酶和VII型幾丁質酶的結構和不同時空表達規律進行了研究。Ⅰ型幾丁質酶(通常CHT5屬于I型幾丁質酶)主要參與昆蟲蛻皮過程中舊表皮的消化,目前已經在鱗翅目、雙翅目、鞘翅目、半翅目和直翅目昆蟲中進行研究。Ⅰ型幾丁質酶基因在昆蟲表皮中的表達量最大(仙笑笑等,2012),該類酶的結構域含有N-末端信號肽、催化域(catalic domain, CAD)、連接區(linker)以及C-末端的幾丁質結合結構域(chitinase binding domain, CBD)(張常,2014),CBD會增強酶對幾丁質的親和能力(夏夢,2015)。研究發現,當I型幾丁質酶被RNAi沉默時,會導致蛻皮失敗,甚至使昆蟲死亡,這表明I型幾丁質酶在蛻皮過程中參與新舊角質層的更新(Nohetal., 2018)。對于昆蟲赤擬谷盜Triboliumcastaneum而言,Ⅰ型幾丁質酶不正常的表達將導致其不能成功的從幼蟲變為蛹,表明Ⅰ型幾丁質酶對外骨骼或圍食膜幾丁質的降解有重要作用(Zhangetal., 2012;張常,2014;Nohetal., 2018)。Ⅶ型幾丁質酶包含一個信號肽和單一的CAD,沒有CBD,該類幾丁質酶的功能尚不明確,有待進一步研究(Liuetal., 2018)。
昆蟲幾丁質酶作為分解昆蟲體壁、中腸、圍食膜等組織中幾丁質的重要水解酶,在昆蟲的消化、變態等生理活動中發揮著至關重要的作用,幾丁質酶的過量或缺失都可影響到昆蟲的正常發育。在本課題組前期的研究中發現HaCHT I可以降解膠體幾丁質,而HaCHT VII對膠體幾丁質則沒有降解活性,但其可將寡聚糖降解成單糖,本研究以此為基礎對棉鈴蟲I型和Ⅶ型幾丁質酶的結構域及其基因表達規律進行了比較分析。為其功能的進一步探究奠定基礎,也為棉鈴蟲生物防治提供有利信息。
1 材料與方法
1.1 材料實驗昆蟲
棉鈴蟲是由新疆生物資源基因工程重點實驗室飼養,飼養條件:溫度26±2℃,RH65%±5%,光周期16 h ∶8 h。
1.2 實驗材料與試劑
RNA提取試劑盒(Trans Zol Up Plus RNA Kit)購于北京全式金公司;Real master mix SYBR green PCR試劑盒購買于invitrogen公司;Taq DNA聚合酶購自大連TaKaRa公司;其余化學試劑均為國產分析純試劑;引物合成及測序均由上海生工完成。
1.3 結構域分析
通過利用在線工具SMART(http://smart.embl-heidelberg.de/)預測對本課題組前期克隆獲得的棉鈴蟲的I型和VII型幾丁質酶HaCHTⅠ、HaCHTⅦ的蛋白結構域,并標注信號肽、催化結構域、幾丁質酶保守區以及幾丁質結合結構域。
1.4 棉鈴蟲總RNA的提取及cDNA第一鏈的合成
為研究HaCHTI和HaCHTVII在棉鈴蟲生長發育過程的蛻皮階段的表達規律,本研究收集了棉鈴蟲不同發育階段蛻皮前后的中腸組織,同時收集了棉鈴蟲6齡幼蟲的頭殼、前腸、中腸、后腸、脂肪體、體壁,收集的所有組織均在-80℃冰箱保存備用。按照制造商的說明提取總RNA,將1 μg的總RNA合成為cDNA:首先,將Oligo d(T)18引物加入到總RNA中,并在70℃下溫育10 min,后在冰上快速冷卻5 min。然后加入MLV逆轉錄酶,在42℃孵育1 h,72℃孵育15 min。最后將反轉錄cDNA產物在冰上冷卻并儲存在-80℃以進行qRT-PCR。
1.5 棉鈴蟲HaCHT I和HaCHT VII 在不同發育階段蛻皮前后和不同組織中的表達量
根據引物設計原則,運用DNAMAN 6.0軟件和 primer 5.0軟件設計了引物,HaCHTI(GenBank accession number:AAQ91786.1)的正向引物:5′-ATGA GAGTGATACTAGCGACGTTGGC-3′;反向引物:5′-CTAAGGCGTCCTGTTCATGAGC CGG-3′。HaCHTVII(GenBank accession number:XP_021180780.1)正向引物:5′-ATGAAAGTTTTA AGTTTATTCTTTGTT-3′;反向引物:5′-CTACAA ATTCAACACAAAAGCTACAGAC-3′。
2 結果與分析
2.1 HaCHTⅠ和HaCHTⅦ結構域的比較
本研究通過利用在線工具SMART(http://smart.embl-heidelberg.de/)預測HaCHTⅠ和HaCHT Ⅶ的信號肽序列、催化區域、連接區域和幾丁質結合結構域(圖1)。分析表明,HaCHTⅠ包括4個區域:N-末端信號肽、一個催化域、連接區以及C-末端幾丁質結合結構域,而HaCHTⅦ僅含有一個N-末端信號肽、一個催化域和一個連接區。
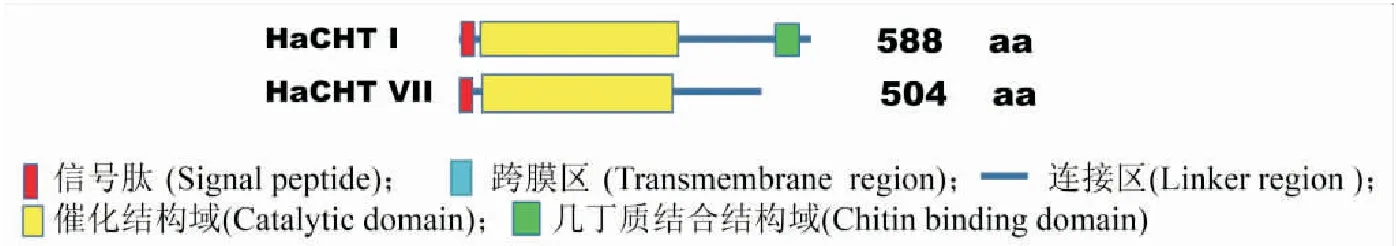
圖1 HaCHTⅠ和HaCHTⅦ結構域的比較Fig.1 Domain comparison between HaCHTⅠ and HaCHTⅦ
2.2 HaCHT I和HaCHT VII 在棉鈴蟲不同組織的表達分析
運用qRT-PCR檢測HaCHTI在棉鈴蟲不同組織的表達量,如圖2-A結果顯示HaCHTI在棉鈴蟲的所有組織部位均有表達,但在統計學上沒有顯著差異(P>0.05)。圖2-B結果顯示的是HaCHTVII在棉鈴蟲不同組織的表達量,HaCHTVII在棉鈴蟲所有組織部位也均有表達,其中在頭殼和體壁中的表達量顯著高于前腸、中腸、后腸和脂肪體(P<0.05);脂肪體中表達量顯著高于前腸、中腸、后腸(P<0.05);前腸、中腸、后腸中沒有顯著性差異(P>0.05)。
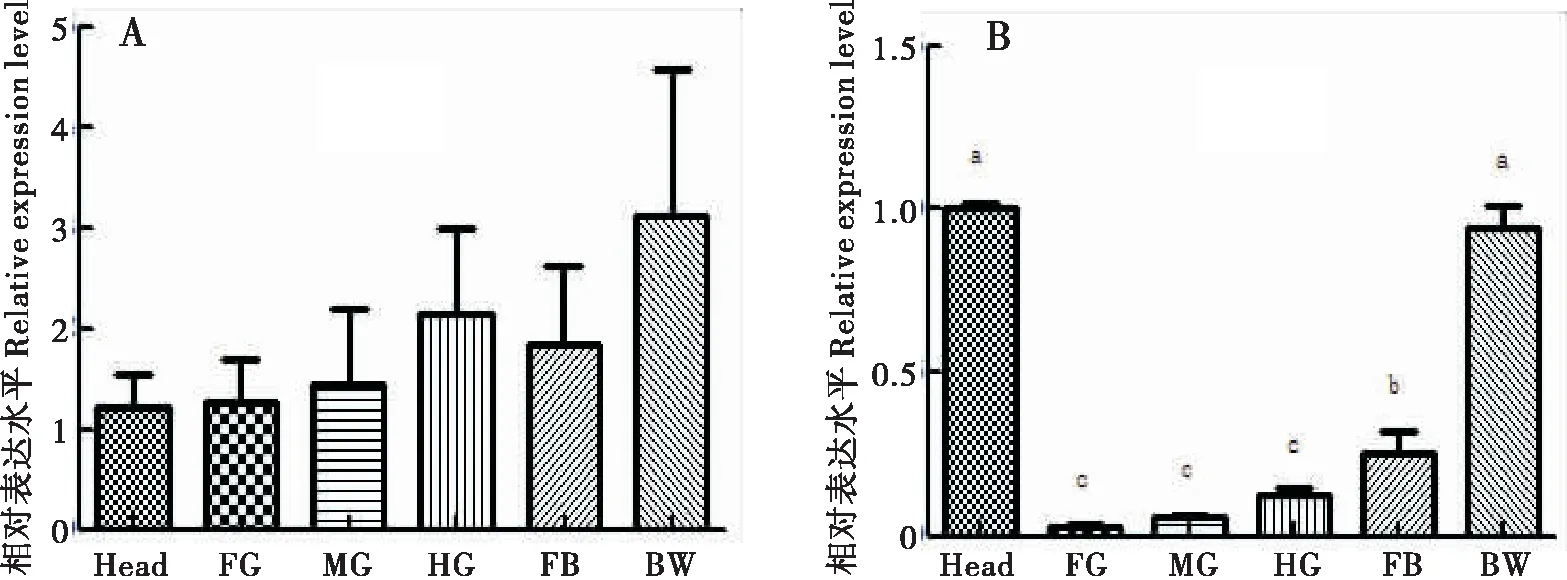
圖2 HaCHT I和HaCHT VII 在棉鈴蟲不同組織的表達量Fig.2 Relative expression level of HaCHT I and HaCHT VII in different tissue of Helicoverpa armigera注:Head, 頭殼; FG, 前腸; MG, 中腸; HG, 后腸; FB, 脂肪體; BW, 體壁。 A,HaCHTI在不同組織中的表達量;B,HaCHT VII 在不同組織中的表達量,qRT-PCR進行3個生物學重復,2個技術重復,數據使用單因素方差分析。Note: Head; FG, Foregut; MG, Midgut; HG, Hindgut; FB, Fat Bady; BW, Bady Wall. A, Expression amount of HaCHT I in different tissue; B, Expression amount of HaCHT VII in different tissue. Three biological replications and two technical replications were performed by qRT-PCR, and the data were analyzed by one-way ANOVA.
2.3 HaCHT I和HaCHT VII在棉鈴蟲幼蟲蛻皮前后中腸中的表達分析
運用qRT-PCR檢測HaCHTI和HaCHTVII基因在棉鈴蟲幼蟲不同組織的表達量,結果表明HaCHTI和HaCHTVII基因在棉鈴蟲幼蟲3齡末至預蛹階段在中腸中均有表達。3齡末期HaCHTI表達量顯著高于4齡末期(P<0.05),4齡末期HaCHTI表達量顯著高于5齡、6齡末期及蛹期(P<0.05),5齡、6齡末期及蛹期沒有顯著性差異(P>0.05),總體分析HaCHTI在3齡、4齡、5齡、6齡末期及蛹期時的表達量呈遞減的趨勢(圖3-A);4齡、6齡初期HaCHTI基因的表達量均顯著高于5齡初期(P<0.05),總體分析HaCHTI基因的表達量在4齡、5齡、6齡初期時呈先減后增的趨勢(圖3-B)。而同一時期HaCHTVII和HaCHTI基因的表達量幾乎成相反關系。在3齡、4齡末期時HaCHTⅦ的表達量沒有顯著性差異,但均顯著低于5齡末期(P<0.05),5齡末期HaCHTVII表達量又顯著低于6齡末期與蛹期(P<0.05),6齡末期與蛹期沒有顯著性差異,總體分析HaCHTVII在3齡、4齡、5齡、6齡末期及蛹期時的表達量呈遞增的趨勢(圖3-C)。4齡、5齡初期HaCHTVII的表達量無顯著差異(P>0.05),但顯著高于6齡初期(P<0.05),總體分析HaCHTVII的表達量呈遞減的趨勢,與HaCHTI在不同齡期的初期表達量模式不同(圖3-D)。
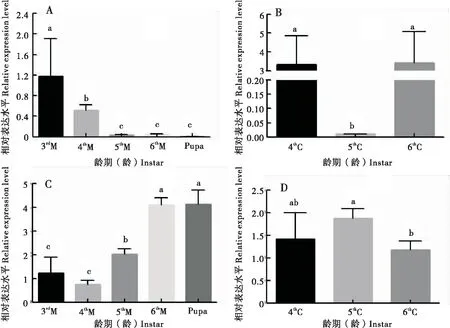
圖3 HaCHT I和HaCHT VII 在棉鈴蟲幼蟲發育階段的表達量Fig.3 Expression of HaCHT I and HaCHT VII in the developmental stage of Helicoverpa armigera larvae注:3rdM,3齡末期;4thM,4齡末期;5thM,5齡末期;6thM,6齡末期;Pupa,蛹期;4thC,4齡初期;5thC,5齡初期;6thC,6齡初期。A,HaCHT I在幼蟲3齡、4齡、5齡、6齡末期以及蛹期的表達量;B,HaCHT I在幼蟲4齡、5齡、6齡初期表達量;C,HaCHT VII 在幼蟲3齡、4齡、5齡、6齡末期以及蛹期的表達量;D,HaCHT VII 在幼蟲4齡、5齡、6齡初期的表達量。qRT-PCR進行3個生物學重復,2個技術重復,數據使用單因素方差分析。Note: 3rdM, 3 inster telephase; 4thM, 4 inster telephase; 5thM, 5 inster telephase; 6thM, 6 inster telephase; Pupa, Pupa stage; 3rdC, 3 inster initial; 4thC, 4 inster initial; 5thC, 5 inster initial; 6thC, 6 inster initial. A,Expression amount of HaCHT I at the end of the 3rd, 4th, 5th and 6th instar and the pupal stage of H. armigera; B,Expression amount of HaCHT I at the beginning of the 4th, 5th and 6th instar of H. armigera; C,Expression amount of HaCHT VII at the end of the 3rd, 4th, 5th and 6th instar and the pupal stage of H. armigera; D,Expression amount of HaCHT VII at the beginning of the 4th, 5th and 6th instar of H. armigera. Three biological replications and two technical replications were performed by qRT-PCR, and the data were analyzed by one-way ANOVA.
3 結論與討論
在昆蟲的生長發育過程中,由幾丁質組成的外骨骼及中腸的圍食膜限制了體內器官的生長,因此,昆蟲在生長發育過程中會出現蛻皮現象(Riddifordetal., 2016)。在昆蟲蛻皮過程中,幾丁質酶負責消化圍食膜和外骨骼中的幾丁質。昆蟲幾丁質酶是數量龐大且結構多樣化的酶家族(Merzendorfer and Zimoch, 2003)。近年來,由于其在幾丁質代謝中的重要生化功能而倍受關注。幾丁質酶是一種內切酶,可降解幾丁質為幾丁寡糖。隨著生物信息學的不斷發展,通過對昆蟲幾丁質酶及其類似蛋白的催化域進行系統發育分析,可將昆蟲幾丁質酶至少劃分為10種類型(Arakaneetal., 2003; Zhangetal., 2011; 夏夢,2015;Liuetal., 2018),每種類型具有各自的結構,其中有些幾丁質酶具有一個共同的結構域,即幾丁質結合結構域(CBD區)。CBD是昆蟲幾丁質酶的一個重要結構,具有6個保守的半胱氨酸,可能形成了3個二硫鍵(Sinuetal., 2010)。其中CBD結構域可增強幾丁質酶與不溶性底物之間的親和性,所以含有CBD結構域的幾丁質酶可能對降解多聚幾丁質比降解幾丁寡糖更加有效(Yasuyukietal., 2003)。
本研究對棉鈴蟲幾丁質酶HaCHTⅠ和HaCHTⅦ以及前期所獲得的HaCHTⅣ的結構域進行比較,發現HaCHTⅠ和HaCHTⅣ在結構上都具有CBD區,而HaCHTⅦ則沒有CBD區。本課題組前期對HaCHTⅠ和HaCHTⅣ的底物探索發現,兩者均具有降解膠體幾丁質的作用(古新蓉等,2019;麥麥提艾力等,2019)。在幾丁質酶降解幾丁質的過程中,幾丁質酶的CBD就會錨定膠體幾丁質,增加對底物的親和能力(Zhuetal., 2008)。當幾丁質酶沒有CBD結構域時,也可降解幾丁質,但是其活性很低(Gentaetal., 2009)。赤擬谷盜體內,異常表達的Ⅰ型幾丁質酶則會導致昆蟲無法順利的從幼蟲蛻變為蛹,說明Ⅰ型幾丁質酶在參與角質層幾丁質降解或圍食膜的更換中起重要作用。大多數Ⅳ型幾丁質酶在中腸或脂肪體中表達,還有少數Ⅳ型幾丁質酶具有特異的表達時期(Zhuetal., 2008),Ⅳ型幾丁質酶的結構域組成以及表達方式的特殊,說明其對昆蟲具有十分重要的生理功能。有相關研究報道,當沉默歐洲玉米螟Ostrinianubilalis體內的Ⅳ型幾丁質酶(OnCHT)時,其中腸圍食膜的孔徑變小而且對食物的消化速率有明顯的下降,最終會影響昆蟲的生長發育,甚至會抑制昆蟲的生長(Khajuriaetal., 2010)。本文推測HaCHTⅠ和HaCHTⅣ可能對圍食膜均有影響,這可能與它們有相同的CBD有關。HaCHTⅦ對膠體幾丁質就沒有幾丁質降解活性,卻能將殼二糖降解成單糖(麥麥提艾力等,2019),分析可能與其沒有CBD有關,這就為HaCHTⅦ在棉鈴蟲體內起到將寡聚糖降解為單糖(N-乙酰基葡糖糖)提供了有力的證據,并為新幾丁質的合成提供起始底物,參與昆蟲的生長發育過程。為此本研究采用qRT-PCR方法對棉鈴蟲幾丁質酶基因HaCHTI在棉鈴蟲不同組織及幼蟲中的時空表達特性進行分析,發現HaCHTI在棉鈴蟲的各個組織中均高表達,在整個發育過程中均有一個基礎水平的表達,表明HaCHTⅠ在棉鈴蟲的生長發育階段的蛻皮過程中起著至關重要的作用,在蛻皮前后的表達量變化很大,尤其是4齡和6齡蛻皮后的表達量很高,有可能此期發揮著更重的作用。據相關報道顯示赤擬谷盜的Ⅰ型幾丁質酶基因TcCHT5在幼蟲-成蟲蛻皮過程中高表達(Zhuetal., 2008),但桔小實蠅Bactroceradorsalis中Ⅰ型幾丁質酶基因BdCHT5在幼蟲-蛹和蛹-成蟲階段均有高表達,推測BdCHT5在幼蟲-蛹和蛹-成蟲變態過程中調控幾丁質的降解(Liuetal., 2018);家蠶Bombyxmori的I型幾丁質酶BmCHT5在3齡幼蟲蛻皮前開始高表達,并且在蛻皮過程以及蛻皮后也高表達,蛻皮12 h后開始消失(Kimetal., 1998),后期研究發現,BmCHT5在5齡幼蟲蛻皮前后以及化蛹期間表達較高,推測BmCHT5主要在該階段參與昆蟲的蛻皮(Zhangetal., 2016);美國白蛾Hyphantriacunea的I型幾丁質酶基因HcCHT5在4齡幼蟲蛻皮前(60 h)開始高表達,并在蛻皮過程以及蛻皮后也高表達,蛻皮12 h后開始降低(Kimetal., 1998)。棉鈴蟲、赤擬谷盜、家蠶、美國白蛾均是全變態昆蟲,綜合分析可以得出,HaCHTI可能在棉鈴蟲蛻皮過程中發揮著降解表皮幾丁質成分的作用,以滿足棉鈴蟲幼蟲的不斷生長發育,這與前期對Ⅰ型幾丁質酶功能的研究與報道是一致的,對棉鈴蟲幾丁質酶基因HaCHTVII在棉鈴蟲不同組織及幼蟲中的時空表達特性進行分析,HaCHTVII在頭殼和體壁中表達顯著高于其他組織,蛻皮前后表達量的變化較大,尤其是6齡末期和預蛹期的表達量相對很高,且本研究在飼養棉鈴蟲的過程中發現,由6齡末期到預蛹的過程中棉鈴蟲的表皮和頭殼的變化較大,因此推測從幼蟲到預蛹階段HaCHTⅦ可能對頭殼和體壁的變化發揮重要作用。
本研究通過對棉鈴蟲I型和Ⅶ型幾丁質酶HaCHTⅠ、HaCHTⅦ的結構域進行比較分析后,發現兩者具有相似的結構域,唯一不同點在于I型比Ⅶ型多了一個CBD。兩種幾丁質酶基因的時空表達也存在明顯的差異,其中HaCHTI在4齡初期和6齡末期的表達量顯著高于其他齡期,在各個組織中均有表達且沒有顯著性差異,因此推測HaCHTI參與其角質層幾丁質降解的更換過程,從而促進舊表皮的降解及新表皮的生長,以滿足棉鈴蟲發育和變態需求;HaCHTVII在6末及預蛹時高表達,在頭殼和體壁中高表達,說明HaCHTVII可能對棉鈴蟲外骨骼系統中幾丁質的形成具有重要作用。
在本研究中對幾丁質酶基因HaCHTI和HaCHTVII只進行了初步的比較分析,結合本課題組前期對這些酶底物研究的結果,發現HaCHTⅠ可以降解膠體幾丁質,而HaCHTⅦ對降解膠體幾丁質沒有降解活性,但其可將寡聚糖降解成單糖,可能為新的幾丁質的合成提供起始底物,推測這可能與其沒有結構域CBD有關。若對HaCHTⅠ和HaCHTⅦ進行RNA干擾、抑制、突變等手段破壞酶活性,檢測其對圍食膜的形態結構、昆蟲的生長發育過程的影響,最終探明兩種幾丁質酶的功能差異,為其進一步的開發利用奠定基礎。
