19882018年滇池氮磷比的時空演變特征與原因解析*
2021-01-12 06:58:54程國微嚴長安
湖泊科學 2021年1期
關鍵詞:影響
高 偉,程國微,嚴長安,陳 巖
(1:云南大學生態與環境學院,云南省高原山地生態與退化環境修復重點實驗室,昆明 650091) (2:昆明市生態環境科學研究院,昆明市環境系統模擬與風險評估重點實驗室,昆明 650032) (3:生態環境部環境規劃院,國家環境保護環境規劃與政策模擬重點實驗室,北京 100012)
氮和磷是影響淡水生態系統初級生產力的兩種主要營養元素,氮、磷富集導致的富營養化是全球面臨的普遍水環境問題[1]. 氮磷比是水體氮、磷元素的質量或原子數量比值,是水體氮、磷相對水平的綜合體現. 作為影響藻類生長的重要化學計量指標之一[2],氮磷比變化對水域生態系統結構和功能具有重要影響,如低氮磷比與水體固氮藍藻的出現具有關聯性[3],氮磷比也被用于判識水體氮磷限制類型[4-5]. 現有研究表明氮磷比對藻類的影響較為復雜,藻類生長受氮磷比和氮、磷在水體中絕對濃度的共同影響[6],且在不同的水體中響應機制存在差異. 此外,水體氮磷比的變化不僅影響本地水體的生態系統健康,還可能對下游水體造成不利的生態影響[7]. 由于氮磷比的變化不僅取決于氮、磷的絕對濃度變化,還受到氮和磷相對濃度變化的直接影響,因此氮磷比的主要影響因素既是氮和磷濃度影響因素的集合,也與影響它們相對濃度的因素有關. 現有研究主要關注氮磷濃度的變化及其原因,但對它們相對濃度變化即氮磷比的驅動研究較少. 目前對湖泊氮磷濃度影響的定量研究主要包括相關性分析法、統計學建模和機理過程模型3種. 相關性分析法是基于相關系數(線性相關系數、非線性相關系數、灰色關聯度等)判斷因素之間的相關強度,如通過計算水質濃度與驅動因素之間的灰色關聯度識別影響湖泊水質指標變化的因素[8],該方法計算較為簡單,但只能識別單因素的獨立作用. 統計模型法是建立湖泊氮、磷濃度與多個驅動因子之間的回歸關系,通過統計顯著性篩選驅動因子,通過回歸系數判斷驅動強度,該方法在湖泊水質變化研究中得到廣泛應用[9-10],常用的統計方法有多元線性回歸[9]、隨機森林[11]和因素分解法. 機理模型法主要是采用湖泊模型建立不同輸入條件與湖體水質的定量關系,通過運行情景模擬出湖體水質變化的主要因子,常用的有EFDC水動力-水質模型[12]. 然而,機理模型建模的難度較高,對數據量和建模者使用經驗有較高要求,在實際應用中難度較高. 湖泊氮磷濃度的影響因素較為復雜,主要包括流域氮磷輸入負荷[12]、氣象條件[10]和氮磷削減工程措施[13]等. 統計模型能夠有效地處理多因素綜合影響效應問題,模型相對簡單,本研究使用統計模型分析滇池氮磷濃度和氮磷比時空變化的影響因素及其效應.
滇池的富營養化持續歷史較長,氮磷超標是滇池水環境治理長期面臨的難題. 隨著城市污水處理、環湖截污和生態補水等治理工程的實施,滇池的氮磷濃度發生了顯著變化[11],湖體氮磷比隨之改變,并對湖體優勢藻類產生了影響[14]. 目前針對滇池的氮磷比演變特征及其驅動力的研究相對較少,主要集中于氮磷比對藻類影響上[14]. 對滇池氮磷濃度變化與來源的研究相對較多[11-12, 15],但主要集中于影響相對重要性的識別和污染來源角度. 滇池氮磷比長期演變趨勢及其多種因素的綜合驅動影響尚缺乏相關研究. 本研究基于滇池1988-2018年的多年水質觀測數據,評估了滇池氮磷比在時間和空間上的變化趨勢;基于氮磷比的影響因素,定量解析了不同因素對滇池氮磷比變化的貢獻,從而識別滇池氮磷比的主控因子. 氮磷比變化與驅動效應研究對科學診斷湖泊生態系統狀態變化方向并制定調控方法具有重要意義,本研究可為湖泊氮磷比變化趨勢與驅動效應識別提供案例參考和方案借鑒.
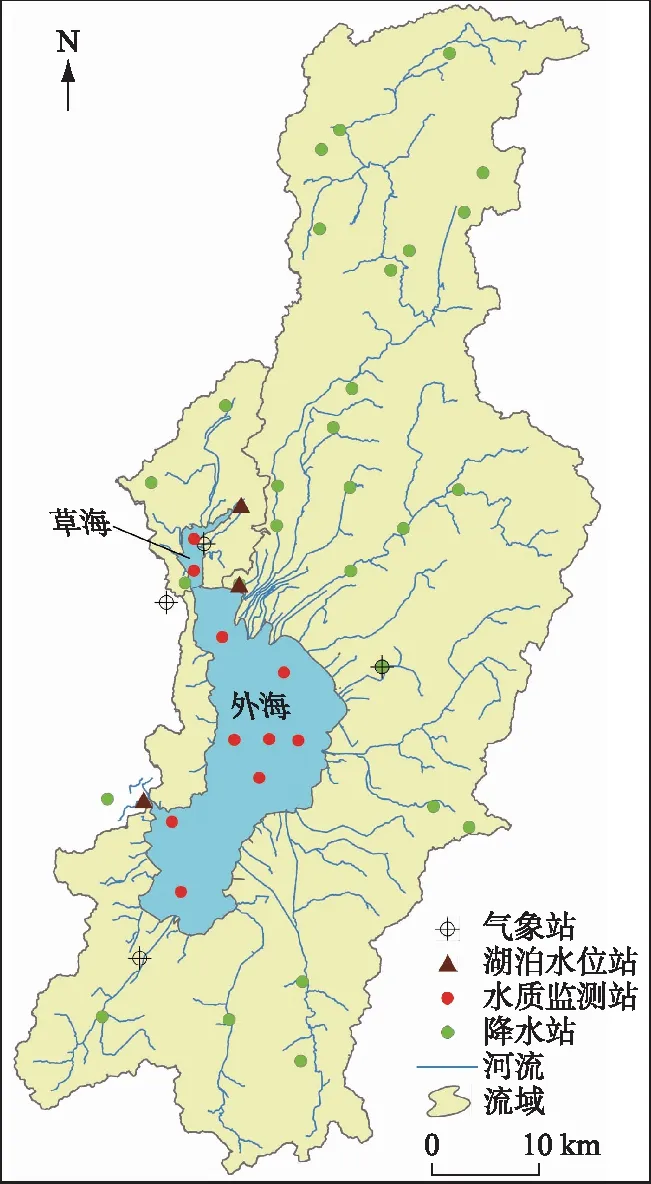
圖1 滇池流域及其氣象、水文與水質監測站點分布 Fig.1 Location of metrological, hydrological and water quality gauging stations of Dianchi Basin
1 研究區概況
滇池是我國第六大淡水湖泊,是云貴高原面積最大的湖泊,湖泊總面積309.5 km2(1887.4 m),湖容15.6億m3,平均水深5.3 m(圖1). 1996年滇池被海埂大壩分割為北部的草海和南部的外海兩個子湖,外海是滇池的主體,面積為289.2 km2. 草海由人工出口西園隧道出流,外海由海口河出流,兩者已沒有水量聯系. 滇池水質自1970s末開始惡化,成為我國水體富營養化嚴重的“三湖”之一. 為改善滇池的水環境,滇池流域自“八五”以來實施了一系列污染防治工程[16],水質得到有效改善[11]. 2013年12月底牛欄江-滇池補水工程實施,牛欄江調水成為滇池的重要水源,對滇池草海和外海開展了生態補水. 相對于巢湖和太湖,滇池在“三湖”中的富營養化程度最高[15]. 根據云南省2018年環境狀況公報[17],滇池草海的總氮為劣V類,總磷為IV類,外海總氮和總磷均為IV類. 雖然近年來水質有所好轉,特別是2009年以來滇池草海和外海的總氮與總磷濃度有顯著下降[15],但仍然未達到水環境功能區(草海IV類,外海III類)的要求,滇池仍然處于輕度富營養化狀態.
2 材料與方法
2.1 研究方法
2.1.1 氮磷比時間變化趨勢分析 指標時間變化趨勢判別是水環境質量評估的重要內容,目前主要采用主觀判斷和統計分析兩種方法. 主觀判斷是根據指標數值隨時間增長的變化方向判斷數據的趨勢,在湖泊污染指標變化趨勢分析中獲得廣泛應用[18],但存在主觀性較強且在數據波動較大時不確定性極高的問題. 統計分析方法是時間序列分析的另一主流方法,常用的有參數檢驗法和非參數檢驗法. 參數檢驗法最常用的是線性趨勢回歸檢驗[19];非參數檢驗法類型較多,常用的有Mann-Kendall檢驗法和Daniel檢驗法[8]. 本研究選擇線性趨勢檢驗和常用的Mann-Kendall非參數突變檢驗法對滇池氮磷比的時間變化趨勢與突變進行判別,其中Mann-Kendall突變檢驗具體方法參見文獻[20],該方法不要求時間序列的數據滿足正態分布.
2.1.2 氮磷比驅動效應模型 驅動效應模型是用于解析影響因子對被影響變量變化貢獻的數學模型. 本研究使用多元線性回歸法,該方法是通過建立自變量和因變量的定量關系,基于回歸系數和變量數值測算自變量對因變量的貢獻. 該方法在污染源解析中應用較多[21],近年來在水文變化驅動力分析中也有應用[22]. 由于多元線性回歸模型具有結構清晰、操作簡便和應用范圍廣的特點,本研究選擇多元線性回歸模型對氮磷比驅動效應進行解析. 多元線性回歸模型對驅動因子的效應解析分為兩步,第一步是構建多元回歸方程,采用逐步回歸法識別出具有統計顯著性(P<0.05)的驅動因子及其回歸系數;第二步,基于回歸系數和變量均值測算各因子對因變量的貢獻. 具體公式為:
(1)
(2)
(3)
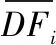
為進一步分析湖泊氮和磷濃度變化的影響因素,建立了氮、磷濃度與降水、氣溫、流域氮磷輸入等驅動因子的多元線性回歸模型,方程的形式與公式1相同. 為了消除數據偏度分布的影響,對自變量和因變量進行了取自然對數處理[10],對取值有0的數據進行Z值(歸一化為均值為0、標準差為1的Z值)轉換.
2.1.3 氮磷比驅動力計算模型 在本研究中,根據文獻調研和數據可得性,主要評估流域氮磷輸入、氣象條件和水文條件3類驅動因子的驅動效應. 其中流域氮和磷輸入分別采用NANI[23]和NAPI[24]模型計算,兩者均是基于物料平衡原理計算流域的人為氮、磷輸入量,NANI估算流域化肥施用輸入、大氣沉降輸入、食品與飼料凈輸入和固氮農作物種植輸入等來源的氮輸入量,NAPI估算流域化肥施用、食品與飼料凈輸入和非食品輸入3項磷輸入量,NANI和NAPI的具體模型與參數參見文獻[9, 25]. 氣象條件是基于逐日降水和逐日平均氣溫計算了滇池流域的平均降水、汛期降水和平均氣溫. 水文指標主要使用湖泊平均水位作為水文驅動因子. 污染物削減和調水數據分別采用污水處理廠削減量和牛欄江滇池補水工程調水量,表征污染控制措施.
2.2 數據來源
根據研究內容,使用的數據分為4類:滇池氮磷監測數據、氣象水文數據、人為氮磷輸入數據和污染控制措施(污水處理廠削減和外流域調水). 氮、磷數據包括滇池流域草海和外海10個點位(圖1)1988-2018年逐月總氮和總磷濃度數據(表1),數據來源于昆明市生態環境監測中心. 氣象數據主要收集了降水、湖泊水位和氣溫,其中降水數據收集了1988-2018年流域27個降水站(圖1)的逐日降水量,水文數據收集了滇池大觀樓、海埂、中灘站3個水位站的同期逐月平均水位(圖1),降水和水文數據來源于云南省水文水資源局;同期昆明、晉寧、太華山和呈貢4個國家氣象站(圖1)的逐日平均氣溫數據來源于資源環境數據云平臺(http://www.resdc.cn/Default.aspx). 滇池流域1988-2015年的NANI和NAPI數據來源于文獻[13],2016-2018年基于經濟社會統計數據和文獻相同的方法計算,統計數據來源于昆明市統計年鑒. 污染物削減和調水數據來源于昆明市生態環境保護局和昆明市水務局.
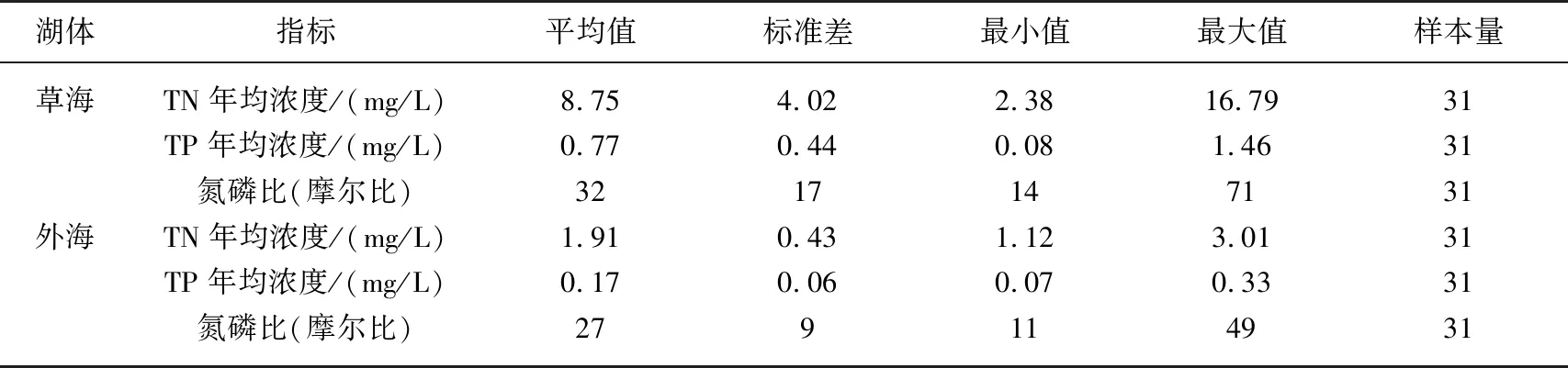
表1 1988-2018年滇池氮、磷濃度基本統計特征
3 結果與討論
3.1 滇池氮磷比的時間演變趨勢
基于滇池草海和外海的多年氮磷監測數據,得到兩個子湖泊氮磷比的逐年變化. 1988-1997年草海的氮磷比平均為21.3±11.2,到2009-2018年增長到51.3±13.7;外海從1988-1997年的20.9±5.10上升到近10年的32.8±4.9. 線性趨勢檢驗結果表明,1988-2018年滇池草海和外海的氮磷比均呈現顯著線性上升趨勢,其中草海年均上升1.3(P<0.01),外海年均上升0.7(P<0.01),草海氮磷比上升速度是外海的2倍(圖2). 草海和外海氮磷比的Mann-Kendall趨勢檢驗的統計值分別為4.62和3.98,均呈顯著上升趨勢(P<0.01),與線性趨勢檢驗的結果一致. 然而,值得注意的是,雖然草海和外海的氮磷比均呈現上升趨勢,但草海、外海的氮磷比線性相關系數僅為0.32,低于0.05的顯著性水平,說明兩者的環境驅動因素可能存在差異. 從Mann-Kendall的突變檢驗結果看,研究期間滇池草海和外海的氮磷比均存在顯著突變. 其中,草海在2008年發生了上升突變,外海在2004年也發生了上升突變,兩湖的突變方向一致,但時間上草海滯后于外海(圖3).
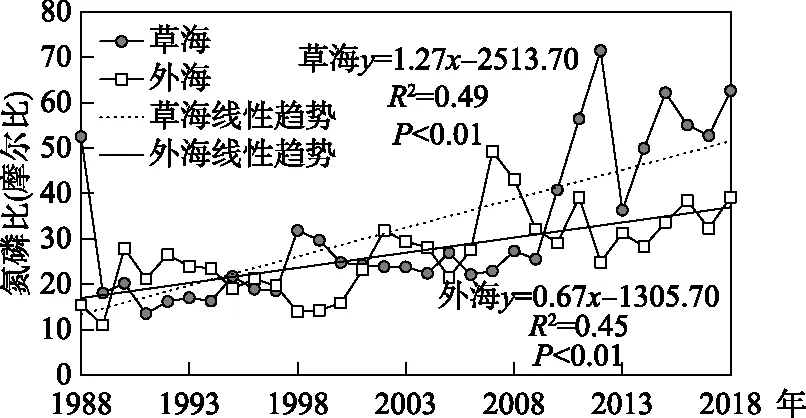
圖2 1988-2018年滇池氮磷比變化Fig.2 Change of nitrogen to phosphorus ratio of Lake Dianchi from 1988 to 2018
3.2 滇池氮、磷濃度的變化特征
根據氮磷比的突變時間,對突變前后的湖體氮、磷濃度變化進行了分析(圖4). 對于草海,突變前后的氮、磷濃度存在相反的變化趨勢,1988-2008年,草海的TN和TP濃度均呈現顯著的上升趨勢(P<0.01),2008-2018年則轉變為顯著下降(P<0.01). 從兩個階段的斜率看,突變前的TN和TP年均濃度分別上升0.49 和0.04 mg/L,相對于突變前期(1988-2008年)均值分別提高0.05和0.04倍,TN濃度的相對增長速度較TP快,因此導致前期氮磷比呈現上升趨勢;突變后期(2008-2018年)草海TN和TP年均濃度分別下降
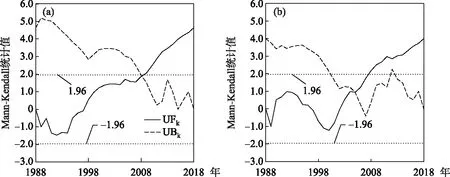
圖3 1988-2018年滇池(a:草海;b:外海)氮磷比Mann-Kendall突變檢測(UFk和UBk分別為Mann-Kendall正向和逆向序列的統計檢驗值)Fig.3 Abrupt change of nitrogen to phosphorus ratio of Lake Dianchi (a: Lake Caohai; b: Lake Waihai) from 1988 to 2018 based on Mann-Kendall test (UFk and UBk are statistics of Mann-Kendall)
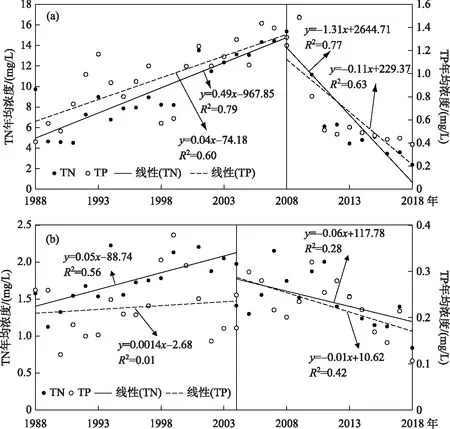
圖4 1988-2018年滇池氮、磷濃度變化(a:草海;b:外海)Fig.4 Temporal change of nitrogen and phosphorus concentrations of Lake Dianchi from 1988 to 2018 (a: Lake Caohai; b: Lake Waihai)
1.31 和0.11 mg/L,相對于突變后期均值分別下降0.18和0.26倍,TN濃度的下降速度慢于TP,從而導致氮磷比上升. 對于外海,突變前期(1988-2004年)僅TN濃度有顯著上升趨勢(P<0.01),因此突變前期氮磷比上升是TN濃度升高導致的結果;突變后期(2004-2018年)的TN和TP濃度均呈現顯著下降趨勢(P<0.01),由于TN濃度較TP的相對下降速度慢(年均下降量相對于突變后期均值分別為0.03和0.04),導致氮磷比呈現上升趨勢. 綜上,滇池草海和外海的氮磷比在突變前期與后期均呈現上升趨勢,但原因有所差異. 滇池草海在突變前是由于TN濃度上升相對速度較TP快,突變后是由于TN濃度下降速度較TP慢;外海在突變期是TN濃度上升單一因素導致的,突變后是TN濃度下降速度慢于TP導致的.
綜合現有文獻[8,10-11,13,25],可將湖體氮、磷水平的影響因素歸納為流域氣象、水文、污染負荷和治理水平等. 氣溫可通過影響氮磷相關化學過程和微生物活動改變湖體的氮、磷水平[26],進而對氮磷比可能造成影響;降水是湖泊水量的直接來源之一,同時也是陸域非點源入河的關鍵影響因素,因此可通過改變湖體水文和污染入湖量影響氮、磷濃度水平[10];研究表明,風速是湖泊浮游植物生物量的重要影響因素,成為部分湖泊藍藻水華暴發的重要驅動力[26];水位是湖泊水量的直接表征指標,水位變化可通過影響水環境容量改變湖體氮、磷濃度[13];人為氮、磷輸入是流域最大的污染來源,可直接改變湖體的氮、磷水平[11, 13];調水量對湖體氮、磷的影響主要是通過水文和氮、磷濃度變化兩個途徑[13];此外,流域的污染減排措施可降低氮、磷的入湖水平,對湖體的氮、磷濃度及其相對水平產生影響[13].
以氮、磷濃度為因變量,以流域年均氣溫、年降水、汛期降水、年均風速、年均水位、NANI、NAPI、年調水量、流域TN削減量、流域TP削減量10個變量為自變量,對1988-2018年滇池氮、磷濃度進行多元線性回歸,結果表明,草海的TN濃度主要受人類活動凈氮輸入量、流域TN削減量和滇池水位控制,其中人類活動凈氮輸入與草海TN濃度呈正相關,而流域TN削減量和滇池水位與草海TN濃度呈負相關. 草海的TP濃度受到人類活動凈磷輸入量、流域TP削減量、牛欄江調水和風速的綜合影響,其中調水量和流域TP削減量表現出顯著的負相關,說明近年來的調水和污染治理工程對滇池草海TP濃度的降低有顯著作用. 對于外海,影響TN濃度的主要變量是流域人類活動凈氮輸入和外流域調水. 從可決系數看,外海TN濃度對這些變量的線性響應關系不如草海,可決系數低于草海,但仍然能夠達到0.613;外海TP濃度回歸方程的可決系數較低,僅0.356,只有調水量一個指標進入方程(表2),說明牛欄江調水對滇池外海的TP濃度產生了較顯著的抑制作用,但是滇池外海TP濃度變化過程較為復雜,可能存在復雜的非線性過程和底泥的源匯作用[27]. 總體來看,滇池氮磷濃度受陸域氮磷輸入負荷、跨流域調水、流域氮磷削減和氣象水文條件影響,其中流域氮輸入負荷對草海和外海的總氮有顯著影響,磷輸入負荷對草海總磷有影響,這與基于隨機森林統計模型得
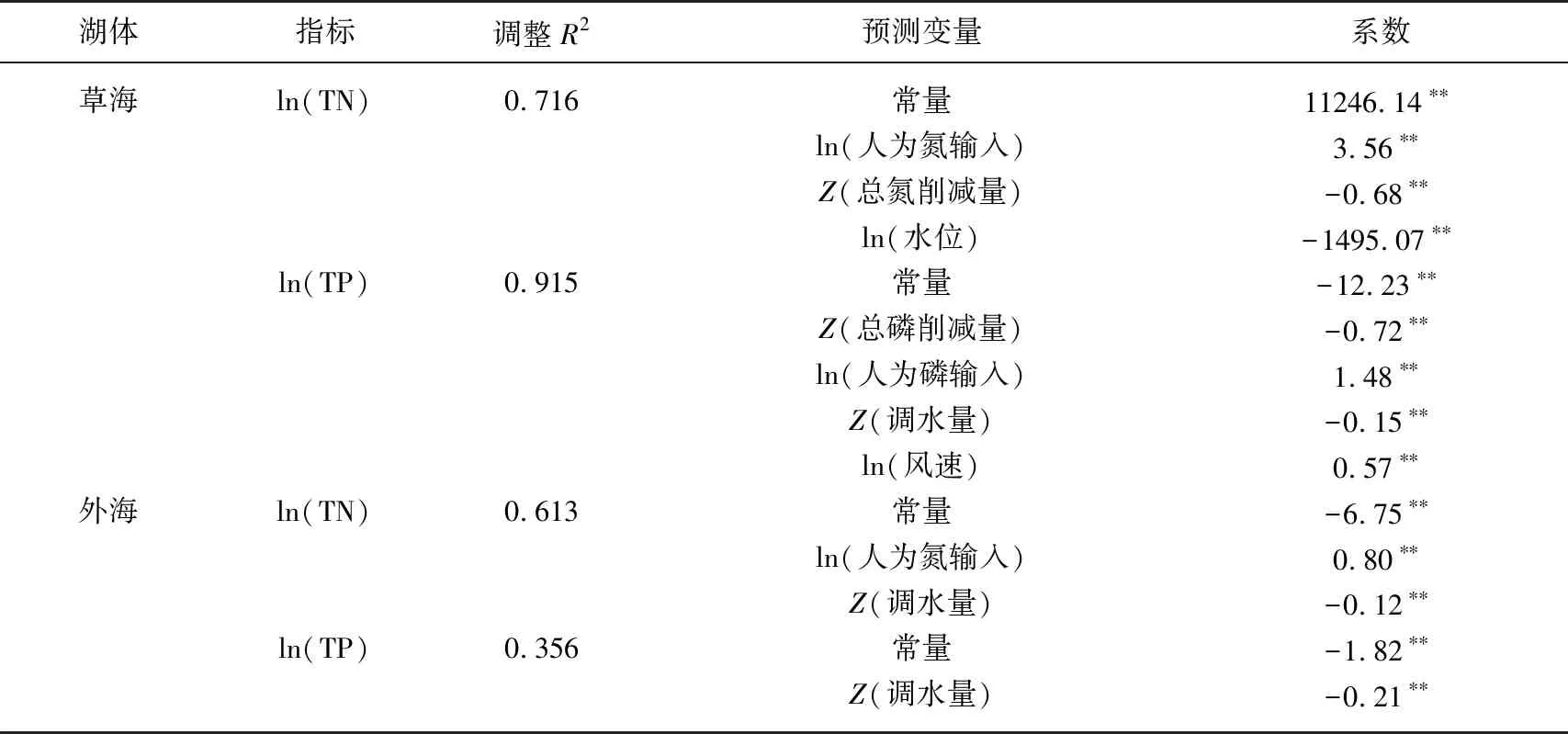
表2 滇池氮、磷濃度的多元線性回歸模型
出的結論較為一致[11],流域氮、磷削減對草海氮、磷濃度影響較大,跨流域調水量對外海的氮、磷濃度和草海的總磷濃度影響較大.
3.3 滇池氮磷比的外在驅動因子及其變化特征
湖泊氮磷比是湖體總氮和總磷濃度的相對數量關系,因此直接取決于湖體的氮、磷水平. 基于氮、磷驅動的主要影響因素,本研究使用11項指標,分別表征流域氣象條件、水文條件、污染負荷和污染治理水平. 滇池流域的氮、磷驅動因子變化如圖5所示.
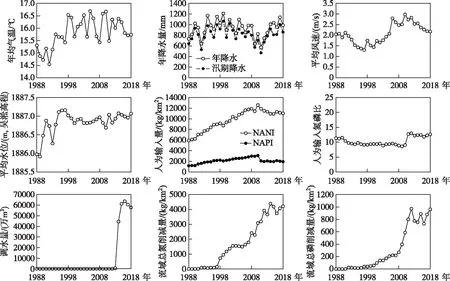
圖5 1988-2018年滇池氮磷比驅動因子變化Fig.5 Driving forces of nitrogen to phosphorus ratio in Lake Dianchi from 1988 to 2018
3.4 滇池氮磷比變化的驅動因子及其效應解析
以湖體氮磷比為因變量,以驅動因子為自變量,分別構建草海和外海的多元線性回歸模型. 基于逐步回歸法,得到草海和外海的氮磷比回歸方程(表3). 影響草海氮磷比的主要驅動因素是流域總磷削減量、人為輸入氮磷比和平均氣溫,3個因子可解釋草海氮磷比變化的80.4%,且均為正向驅動效應. 相對于草海,影響外海氮磷比具有顯著性的驅動因子只有NANI和平均氣溫,回歸模型的解釋力度為0.676,低于草海,表明外海氮磷比的影響因素和氮、磷變化過程更為復雜. 平均氣溫對外海氮磷比的影響是負向驅動,即氣溫越高,氮磷比越低,這與草海的效應相反,說明溫度在外海和草海的氮、磷循環中的作用可能存在差異. 已有研究結果表明[28],氣象因子(包括溫度)對外海TN濃度的影響高于TP,且主要表現為負向效應,這與本研究較為一致,氣溫越高,氮濃度越低,氮磷比呈現下降趨勢. 根據驅動因子貢獻的解析公式,得到滇池草海和外海各驅動因子對氮磷比的影響(圖6). 對草海,年平均氣溫是氮磷比變化的最主要影響因素,其次是流域的人為輸入氮磷比,兩者對草海氮磷比的貢獻為53%,流域的磷削減量對氮磷比的貢獻較小,僅為2%,此外還有45%的氮、磷是由其他未識別因素貢獻的,可能為草海存量;溫度同樣是外海氮磷比的最主要影響因素,占46%,其次是流域人為氮輸入,占14%,剩余40%的貢獻來自于其他未識別因子. 由于底泥是影響滇池氮、磷平衡的重要因素,未識別因子可能主要來自于底泥. 在已有的滇池污染源解析研究中,根據未識別源的貢獻特征將其定義為底泥內源[28]. 湖泊的氮、磷濃度受控因素較多,生物因素也是影響氮、磷的重要因素. 本研究在文獻綜述基礎上選擇了這些非生物因素,未考慮生物因素的可能影響,主要原因是缺乏長時間序列生物觀測資料. 此外,生物因素的影響也會與非生物因素耦合且有時互為因果,如氮、磷濃度變化可改變浮游植物數量,浮游植物數量反過來又改變氮、磷濃度. 現有研究一般將氮、磷濃度變化作為浮游動植物變化的驅動,而不是結果. 綜上,本研究主要考慮非生物因素的影響.
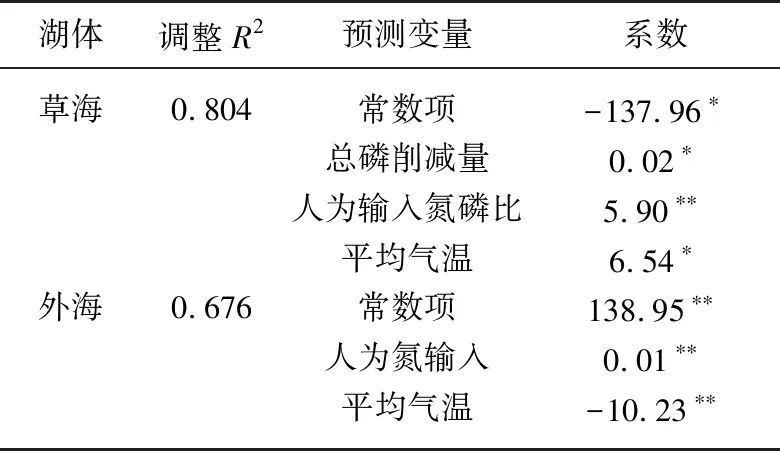
表3 滇池氮磷比多元線性回歸模型參數
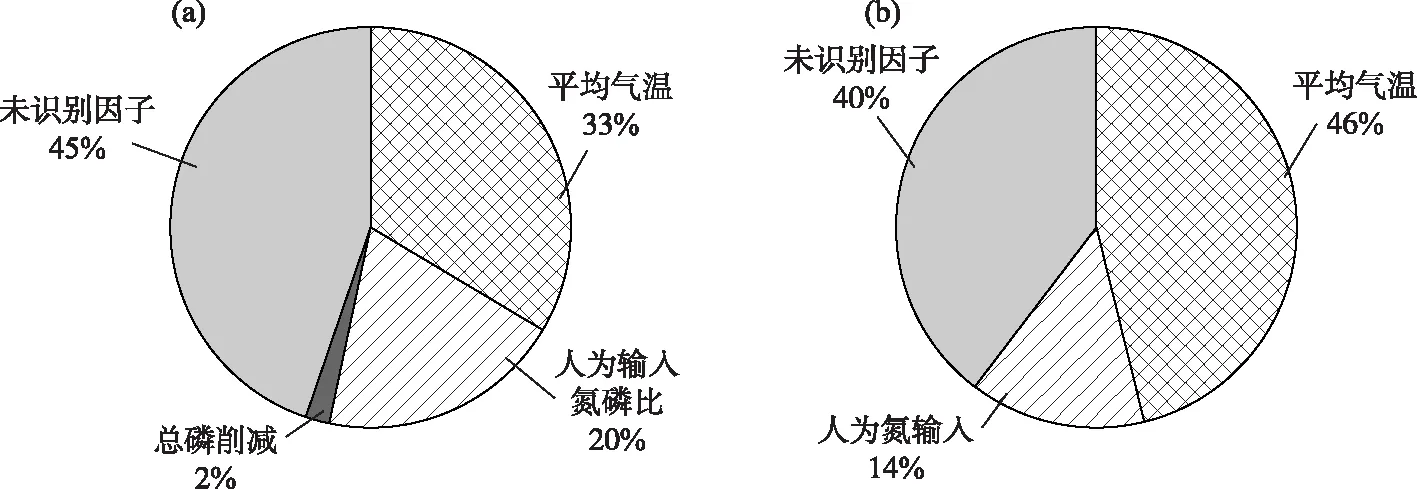
圖6 流域驅動因子對滇池氮磷比變化貢獻(a:草海;b:外海)Fig.6 Contribution of driving forces to N/P ratio in Lake Dianchi (a: Lake Caohai; b: Lake Waihai)
4 結論
基于1988-2018年氮、磷序列濃度觀測數據,對滇池草海和外海的氮磷比時間變化趨勢、突變特征和驅動力進行了定量解析,得到的主要結論如下:
1)1988年以來滇池的氮、磷濃度變化趨勢經歷了從上升到下降的反向逆轉,但氮磷比始終呈現出顯著的上升趨勢,草海和外海兩個子湖的氮磷比平均分別上升1.3和0.7 a-1,草海的氮磷比增速高于外海.
2)草海和外海均發生了氮磷比上升突變,且突變前后氮磷比上升的原因存在差異,突變前氮磷比變化的主要原因是湖體TN濃度上升快于TP,突變后是由于TN濃度下降速度慢于TP下降速度. 滇池的氮、磷濃度受流域氮磷輸入負荷、跨流域調水、流域氮磷削減、風速和湖泊水位的綜合影響,且在草海和外海存在差異.
3)滇池的氮磷比受控因子與氮、磷濃度受控因子不完全重疊. 草海氮磷比的主要控制因子為陸域輸入氮磷比、污水處理廠TP削減量、氣溫,外海氮磷比的主要控制因子為陸域氮輸入和氣溫. 但氣溫對兩個子湖氮磷比的作用相反.
氮磷比是湖泊生態系統重要的化學計量指標,本研究主要從年際變化的角度對其進行趨勢和驅動力解析,由于數據不足,未對氮磷比的季節性變化展開分析,未來有必要進一步研究湖泊季節性氮磷比變化與驅動特征.
猜你喜歡
英語文摘(2022年4期)2022-06-05 07:45:18
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
