抗磺胺二甲氧嘧啶VHH抗體噬菌體庫的構建和鑒定
2021-01-08 12:31:18賀東陽張齊張星星鐘發剛張小鶯王金泉
畜牧與獸醫 2021年1期
賀東陽,張齊,張星星,鐘發剛,張小鶯,3,王金泉
(1. 新疆農業大學動物醫學學院,新疆 烏魯木齊 830052;2. 新疆農墾科學院省部共建綿羊遺傳改良與健康養殖國家重點實驗室,新疆 石河子 832000;3. Department of Biomedical Sciences, Ontario Veterinary College, University of Guelph, Ontario N1G2W1,Canada)
磺胺二甲氧嘧啶(sulfadimethoxine, SDM)是一種磺胺類獸用抗生素,并具有一定毒性作用[1]。由于SDM抗菌譜廣且成本低廉,所以在治療和預防動物傳染病方面的應用極其廣泛,然而大量使用磺胺類獸用抗生素會導致動物體內以及食品中抗生素藥物殘留,過度攝入可能會引起過敏、造血系統功能障礙、急性溶血性貧血等不良癥狀,給食品安全帶來潛在的隱患[2]。世界各國針對SDM在不同的動物性食品中的最高殘留限量(maximum residue limit, MRL)給出了明確的規定,我國SDM在牛、山羊、豬、家禽肌肉中MRL限量為100 μg/kg,肝、腎和脂肪中為100 μg/kg,牛和羊乳中為25 μg/kg;歐盟(EU)設定了磺胺類藥物在肉類、牛奶和雞蛋等動物源性食品中的MRL為100 μg/kg[3];日本規定動物源性食品中單個磺胺類藥物的MRL不超過20 μg/kg;英、美等國家規定肉品中磺胺類藥物的MRL為100 μg/kg[4]。SDM在動物源性食品中的殘留問題與食品安全關系密切,并受到人們廣泛關注。
傳統的藥物殘留分析主要是利用色譜技術和免疫檢測等方法進行定性定量分析[5]。色譜方法結果準確、靈敏,但局限在于待檢樣品需經一系列的預處理,繁瑣費時;此外色譜儀器價格昂貴,只有專業人員才能應用,臨床難以推廣[6]。免疫學方法是一種基于抗原抗體特異性結合而建立起來的方法。目前,國內外檢測磺胺類抗生素殘留主要基于多克隆抗體或小鼠雜交瘤細胞的單克隆抗體。多克隆抗體操作規程相對繁瑣,耗時長,檢測靈敏度不高,有時容易出現假陽性結果,且由于多克隆抗體具有多個抗原表位,特異性較低,檢測過程中容易出現偏差;單克隆抗體雖然特異性較好,但是生產周期長,成本較高[7]。
近年來,基因工程抗體發展迅速,越來越受到科研工作者的青睞,抗體小型化是基因工程抗體的一個主要研究趨勢。駝源重鏈單域抗體(variable domain of heavy chain of heavy-chain antibody, VHH)為駱駝科(駱駝、羊駝或美洲駝)及鯊魚體內存在的天然缺失輕鏈的單鏈抗體,由3個互補決定區(complementarity determining regions, CDRs)構成,其晶體結構呈橢圓形,直徑為2.5 nm,長為4 nm[8]。納米抗體的優勢在于具有更長的CDR3區、體積小、穩定性高、水溶性好、可在微生物系統中大量合成表達,已被廣泛應用于食品安全檢測及臨床診斷[9]。本試驗采用人工合成的免疫原(sulfadimethoxine-albumin bovine serum, SDM-BSA)免疫駱駝,構建抗SDM駝源納米抗體庫,以期為動物源性食品中SDM的殘留檢測提供一種新的方法。
1 材料與方法
1.1 試驗動物和試劑
健康新疆雙峰駱駝,由新疆農墾科學院省部共建綿羊遺傳改良與健康養殖國家重點實驗室提供;弗氏完全/不完全佐劑(FCA/FIA)(Sigma公司,美國);牛血清白蛋白(BSA)、卵清蛋白(OVA)、N-羥基琥珀酰亞胺(NHS)、1-(3-二甲氨基丙基)-3-乙基碳二亞胺鹽酸鹽(EDC)、四甲基聯苯胺(TMB)、二甲基亞砜(DMSO)和RNA Simple Total RNA Kit (天根生物科技公司,北京);膠回收試劑盒、Axygen Mini Prep質粒小量抽提試劑盒(Omega生物科技有限公司,美國);RNA反轉試劑盒(諾唯贊生物科技公司,南京);pCANTAB-5E噬菌體載體、限制性內切酶SfiⅠ、T4連接酶、DNA Marker、輔助噬菌體M13K07(NEB公司,美國);TRYPTONE、YEAST EXTRACT(OXOID公司,英國);SDM(國家標準物質中心);HRP標記抗M13抗體(Sigma公司,美國)。
1.2 引物設計
參考雙峰駝VHH基因序列,利用Primer Premier 5.0軟件設計引物,引物序列如表1所示。
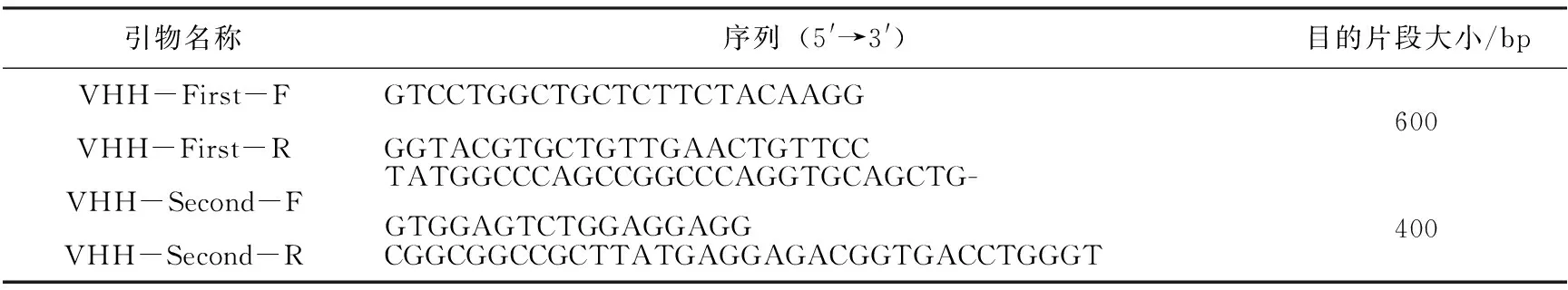
表1 文庫構建所用引物
1.3 磺胺二甲氧嘧啶人工抗原的合成及動物免疫
①A液制備:稱取50 mg BSA溶于2 mL的1 mol/L Na2CO3中冰浴;②B液制備:稱取33.2 mg SDM溶于3 mL 1 mol/L HCl中,完全溶解后冰浴15~30 min;③C液制備:稱取10 mg亞硝酸鈉溶于1 mL蒸餾水中冰浴;④D液制備:將C液逐滴加入B液中,搖勻并置冰浴反應5 min,淀粉KI試紙進行檢驗,若變紫,則加少量尿素,直至不再變紫為止;⑤將D溶液逐滴加入A溶液中,反應過程中保持pH值為9,在4 ℃攪拌反應6 h后,用0.01 mol/L PBS透析3 d,每天換液2次,即制備得SDM-BSA。同理,按此方法制備(sulfadimethoxine-ovalbumin, SDM-OVA),合成的抗原經紫外光譜掃描鑒定后于-20 ℃保存[10]。將合成的人工抗原SDM-BSA與弗氏佐劑等比例混勻并充分乳化(初次免疫使用FCA,之后免疫使用FIA),采用背部皮下多點注射的方式免疫駱駝,共5次,免疫間隔時間為2周。
1.4 噬菌體庫的構建與庫容測定
分離駱駝外周血淋巴細胞,提取總RNA,反轉錄為cDNA,然后用套式PCR擴增出駱駝VHH基因片段。首先使用VHH-First-F、VHH-First-R上下游引物,以cDNA為模板進行PCR擴增,通過凝膠電泳切膠回收擴增片段,然后以回收產物為模板,用VHH-Second-F、VHH-Second-R為上下游引物進行PCR擴增,得到VHH基因片段。分別用SfiⅠ,NotⅠ限制性內切酶進行酶切獲得的VHH片段和pCANTAB-5E噬菌體載體,用T4連接酶將VHH片段和pCANTAB-5E載體置于16 ℃連接過夜。將重組質粒轉化到TG1大腸桿菌感受態細胞中,取少量菌液涂布于氨芐抗性的LB固體培養板,37 ℃培養過夜。次日計算庫容量,隨機挑選單克隆,進行PCR擴增,用1%瓊脂糖凝膠電泳鑒定噬菌體展示文庫的重組率。然后將從平板上刮下來的菌落用LB培養基重懸,此為噬菌體抗體庫,噬菌體庫經實驗室前期鑒定,其庫容為1.08×105CFU,陽性率為96.6%;挑單克隆生長的菌液測序,剩余菌液加入50%終濃度的甘油,分裝凍存于-80 ℃。
1.5 生物淘選與噬菌體克隆的鑒定
1.5.1 SDM噬菌體抗體庫生物淘選
接種至少10倍庫容量的活細胞于100 mL 2×YT-GA培養基的三角瓶中,37 ℃、220 r/min振蕩培養至OD600約為0.4~0.6。按細胞數∶輔助噬菌體=1∶20的比例加入M13K07輔助噬菌體,37 ℃靜置15 min后,37 ℃、220 r/min繼續振蕩培養30 min,將上述培養物4 500 r/min離心15 min,收集菌體,用100 mL 2×YT-AK培養液重懸菌體,30 ℃,250 r/min振蕩培養8 h;次日取出,10 000 r/min,4 ℃,離心15 min,收集上清,加入1/5體積的PEG/NaCl溶液,混勻后置于冰上放置8~10 h至絮狀沉淀出現。
采用固相淘選方法對已構建的噬菌體展示納米抗體庫進行4輪淘選。具體方法如下:1~4輪包被抗原濃度依次分別為:150、75、37.5和18.75 μg/mL,以2% BSA作為第1、3輪封閉液,2% OVA作為第2、4輪封閉液,加入100 μL噬菌體抗體庫至已封閉的酶標板孔中(滴度約1×1011CFU);PBST(含0.5% Tween-20)洗滌10次,PBS洗滌10次;再以50 μL Gly-HCl溶液(pH=2.2)洗脫特異性結合的噬菌體,加入50 μL Tris-HCl (pH=8.5)中和洗脫物。吸取10 μL洗脫物進行滴定,余下的洗脫物擴增后用于下一輪淘選。
1.5.2 噬菌體克隆的鑒定
采用phage ELISA方法鑒定篩選的克隆是否能特異性地結合SDM小分子。取10 μL第4輪洗脫庫,用2-YT培養液梯度稀釋后侵染TG1,然后涂布于氨芐抗性的LB固體培養板上,37 ℃培養過夜。次日,隨機挑取一定數量的單菌落接種LB (Amp)液體培養基,37 ℃、220 r/min培養8 h,以1∶100的比例轉接,繼續培養至OD600為0.4~0.5,再加入M13KO7輔助噬菌體,37 ℃靜置30 min,再將其置于37 ℃、220 r/min培養45 min。再加卡那霉素(終濃度為10 ng/mL),30 ℃恒溫搖床,培養過夜,5 000 r/min離心5 min,收集上清液。將SDM-OVA包被原稀釋成5 mg/mL,100 μL/孔,4 ℃包被過夜,PBST洗滌3次,PBS洗滌3次,隨后加入2% BSA,100 μL/孔, 37 ℃恒溫箱封閉2 h,再加入噬菌體上清液,100 μL/孔,37 ℃孵育40 min,加入HRP標記的抗M13抗體,37 ℃孵育40 min,TMB顯色液孵育10 min,50 μL 2 mol/L H2SO4終止,測定450 nm處吸光度。
2 結果與分析
2.1 PCR擴增VHH基因片段
提取駱駝外周血淋巴細胞總RNA,經1%瓊脂糖凝膠電泳分析可見2條清晰條帶28S rRNA和18S rRNA(圖1A)。
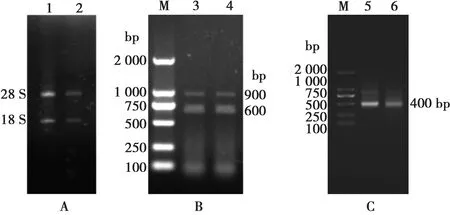
M. 2000 bp DNA Marker;1~2. 28S rRNA和18S rRNA;3~4. PCR第一輪擴增產物; 5~6. PCR第二輪擴增產物圖1 套式PCR擴增VHH基因
以總RNA為模板,反轉錄為cDNA,用VHH-First-F、VHH-First-R上下游引物,以cDNA為模板,進行PCR擴增,通過瓊脂糖凝膠電泳驗證得到600 bp和900 bp的基因片段(圖1B)。切膠回收600 bp基因片段并以此為模板,用VHH-Second-F、VHH-Second-R為上下游引物進行PCR擴增,得到400 bp VHH基因片段(圖1C)。
2.2 噬菌體抗體庫質量的鑒定
對隨機挑取的11個轉化后平板上的單菌落進行測序,用DNAMAN軟件分析測序結果,序列之間顯著不同,且序列符合VHH的典型特征(圖2)。利用進化樹系統對11個單菌落高變區氨基酸序列進行分析,其中VHH8噬菌體克隆菌落氨基酸序列與其他噬菌體克隆菌落氨基酸序列具有明顯差別,氨基酸序列枝長各不相同,相似度各異(圖3),證明了噬菌體抗體庫具有良好的多樣性。

注:黑色區域和灰色區域表示框架區域相同的氨基酸序列,方框中的氨基酸序列分別為CDR1、CDR2和CDR3可變區氨基酸序列圖2 噬菌體展示納米抗體氨基酸序列比對

圖3 進化樹分析噬菌體文庫多樣性
2.3 噬菌體庫生物淘選
經過4輪生物淘選,噬菌體的回收率呈現增長的趨勢,雖然在淘選過程中,第3輪的回收率有所波動,但結果表明噬菌體陽性克隆得到了富集,每輪淘選滴定結果如表2所示;每輪淘選都對洗脫庫進行陽性克隆的鑒定,其結果如圖4所示。對第4輪淘選的洗脫庫進行滴定,挑取93個噬菌體克隆,采用phage ELISA方法進行鑒定,其中3、4、6、8、9、22、24、32、38等編號的噬菌體克隆對SDM具有明顯的結合力(圖5)。

表2 4輪生物淘選噬菌體庫庫容滴定
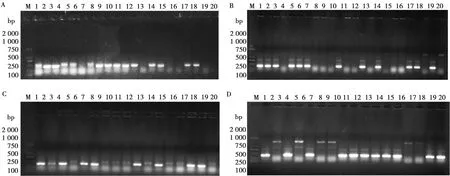
A.一級洗脫庫;B.二級洗脫庫;C.三級洗脫庫;D.四級洗脫庫;M. 2000 bp DNA Marker;1~20.各輪生物淘選隨機挑取單個的噬菌體克隆菌落圖4 4輪洗脫庫陽性克隆PCR鑒定

圖5 噬菌體陽性克隆phage ELISA鑒定
3 討論
本研究利用SDS-BSA人工合成抗原免疫雙峰駱駝,擴增了駱駝重鏈抗體VHH基因, 建立噬菌體展示VHH免疫文庫。本實驗室前期初步測定庫容量大小在1.08×105CFU,陽性率為96.6%。抗體庫的庫容量并不高,試驗過程中在優化載體與目的基因片段連接的摩爾比時發現,在連接體系中有酶蛋白和緩沖液鹽離子的存在,可能會影響和干擾連接產物的轉化,使轉化效率大大降低。盡可能減少對連接產物的干擾和基因片段的損耗,才能提高轉化效率,使抗體庫的質量得以明顯的改善[11]。對于轉化試驗,目前應用最多的是電轉化法和化學轉化法,都是通過外部附加條件,改變細胞膜的通透性,實現轉化的過程[12]。宋歡等[13]建庫時優化連接體系,采用電轉化的方法構建了針對豬繁殖與呼吸綜合征病毒(PRRSV)非結構蛋白2 (Nsp2)的VHH噬菌體文庫,其庫容為6.5×107CFU;何雷等[14]針對中東呼吸綜合征冠狀病毒采用電轉化法構建的噬菌體展示文庫,其庫容達到1.31×108CFU。相對于本試驗所構建的抗體庫,其庫容提高了102~103倍,由此也印證了電轉化法效率比化學轉化法效率要高很多。盡管如此,仍然有許多因素會影響轉化效率,VHH基因的多樣性、感受態細胞的質量、轉化時轉化條件的優化等都會對構建抗體庫的質量產生重大的影響[15]。
本研究采用了固相淘選方法對構建的SDM駝源納米抗體庫進行篩選。每輪篩選時,封閉液的交替使用、噬菌體投入量、洗脫液中Tween-20的含量、洗脫結合的時間及洗脫的次數等眾多因素都有可能影響淘選結果[16]。抗體庫中包含的特異性噬菌體克隆的親和力不盡相同,在整個淘選過程中,逐輪增加洗滌的次數及Tween-20的含量,并逐次降低包被抗原的濃度,從而逐輪增加淘選的嚴苛度,以期獲得高特異性的重組噬菌體。但是也并非嚴苛度越高,就能篩選到高特異性的噬菌體克隆,這樣可能會造成特異性的噬菌體克隆的丟失,所以合理地控制淘選的嚴苛度,才能保障篩選出高親和力的噬菌體。Alshamat等[17]構建的納米抗體庫庫容為5×108CFU,陽性率為95.6%,進行3輪生物淘選使陽性克隆噬菌體得到明顯富集;而本試驗則采用了4輪生物淘選。生物淘選的輪數視試驗的具體情況而定,確保陽性克隆噬菌體得到有效、更好地富集。
4 結論
本研究構建了抗SDM駝源納米抗體庫并對所構建的抗體庫進行4輪生物淘選,最終獲得了對SDM具有明顯結合力的陽性克隆噬菌體。試驗不僅為下游產品的開發奠定了良好的基礎,同時也為SDM在動物源性食品中的殘留檢測方法提供了相應的參考,其應用前景十分廣闊。
