香樟葉肉含晶細胞季節變化研究
2021-01-07 05:13:07尤揚趙明華張曉云王保全郎冬梅徐鈞齊玉杰尚潛
廣西植物 2021年12期
尤揚 趙明華 張曉云 王保全 郎冬梅 徐鈞 齊玉杰 尚潛
摘 要:? 為探討香樟(Cinnamomum camphora)葉肉含晶細胞超微結構的季節變化,闡明香樟葉肉中草酸鈣晶體在春夏秋冬的變化規律。該研究以多年生香樟(C. camphora)葉片為材料,分別于春夏秋冬四個季節露地取樣,制作超薄切片,用透射電子顯微鏡(TEM)觀察葉肉含晶細胞超微結構的變化。結果表明:春季時香樟葉肉中只有少數細胞有草酸鈣晶體,數量較少,晶體結構多為柱狀晶、方晶;夏季時香樟葉肉細胞中隨機分布于液泡的草酸鈣晶體明顯比春季的數量多、體積大、形態豐富,晶體多為柱狀晶、方晶、針晶、簇晶;秋季時香樟葉肉細胞草酸鈣晶體和夏季的類似,數量較多,形態多樣,以方晶和柱狀晶針晶為主,伴有晶簇;冬季時香樟葉肉含晶細胞晶體形態為柱狀晶、方晶、針晶,數量比夏季和秋季的數量略有減少。該研究結果表明在一年四季中香樟葉肉細胞液泡中均有草酸鈣晶體結構存在。
關鍵詞: 香樟, 含晶細胞, 草酸鈣, 超微結構
中圖分類號:? Q44.6
文獻標識碼:? A
文章編號:? 1000-3142(2021)12-1974-07
收稿日期:? 2020-06-13
基金項目:? 河南省科技計劃項目(162102110155);新鄉市科技創新發展專項(CXGG16029);河南省教育廳科學技術研究重點項目(13A180307);河南科技學院高層次人才科研啟動項目;河南科技學院大學生創業實踐計劃項目(2019CY026) [Supported by Science and Technology Project of Henan Province(162102110155); Special Project of Science and Technology Innovation and Development of Xinxiang (CXGG16029);Key Project of Science and Technology Research of Henan Education Department (13A180307); Launch Research Project for High-level Talents of Henan Institute of Science and Technology; Henan Institute of Science and Technology Student Entrepreneurship Practice Plan Project(2019CY026) ]。
作者簡介: 尤揚(1973-),博士,副教授,碩士研究生導師,從事植物逆境生理及解剖結構等研究,(E-mail)youyang1028@126.com。
通信作者
Seasonal changes of crystal idioblasts in mesophyll of Cinnamomum camphora
YOU Yang1,2*, ZHAO Minghua3, ZHANG Xiaoyun1,2, WANG Baoquan1,2, LANG Dongmei1,2, XU Jun1,2, QI Yujie1, SHANG Qian1
( 1. School of Horticulture Landscape Architecture, Henan Institute of Science and Technology, Xinxiang 453003, Henan, China; 2. Henan
Engineering Technology Research Center of Characteristic Horticultural Plants Development and Utilization, Henan Institute of Science
and Technology, Xinxiang 453003, Henan, China; 3. Forestry Workstation of Xinxiang, Xinxiang 453000, Henan, China )
Abstract:? In order to explore the seasonal changes of the ultrastructure of crystal idioblasts in mesophyll of? Cinnamomum camphora, and to elucidate the changing law of calcium oxalate crystals in spring, summer, autumn and winter, in this study with C. camphora as test materials, samples were taken from the open field in the four seasons and ultra-thin sections were made and the ultrastructural changes of mesophyll crystal idioblasts were observed with a transmission electron microscope (TEM). The results were as follows: Only a few mesophyll cells had calcium oxalate crystals in spring and the number was relatively small, which were mainly in shape of columnar crystals and square crystals;? Calcium oxalate crystals randomly distributed in the vacuoles of mesophyll cells in summer were obviously more in number, bigger in volume, and richer in morphology than those in spring, and the crystals are mostly columnar crystals, square crystals, needle crystals and cluster crystals; The calcium oxalate crystals of the C. camphora mesophyll cells in autumn were similar to those in summer, with a large numbers of? and various forms, mainly square and columnar needle crystals, accompanied by crystal clusters; The crystal form of C. camphora mesophyll crystal idioblast in winter was columnar crystals, square crystals and needle crystals, and the number was slightly less than those in summer and autumn. The crystal structure of calcium oxalate existed in the vacuole of C. camphora mesophyll cells throughout the year.
Key words: Cinnamomum camphora, crystal idioblast, calcium oxalate, ultrastructure
在植物界超過215科的植物中發現有草酸鈣晶體,依據其形狀特點可將草酸鈣晶體分為柱晶、簇晶、針晶、方晶、砂晶5種常見類型(Lersten & Horner,2006;嚴巧娣和蘇培璽,2006)。草酸鈣的生理作用主要集中在解毒作用(Franceschi & Nakata,2005)、鈣調節(Hudgin et al.,2003)、支持作用(Borchert,1986)、離子平衡(Volk et al.,2002)、防御功能(Volk et al.,2002)等方面。含晶細胞的晶體主要成分中占絕大多數的是草酸鈣(Finley,1999)。植物中沉積草酸鈣晶體的現象普遍存在,所有晶體來源均由環境中的鈣與生物體自身合成的草酸所形成(Nakata et al.,2003)。草酸鈣晶體大小和形態的多樣性、空間分布及分布優勢等,促成了許多有關植物細胞液泡內晶體功能的假說。事實上,含晶細胞在植物許多組織中都有發現,大部分聚集在特化的細胞及含晶細胞的液泡中(Nakata & Mccoonn,2003)。植物液泡中含晶細胞的存在可能暗示在某些方面具有重要的生理功能,這些功能與外界環境的變化可能有某些聯系。
香樟(Cinnamomum camphora)為樟科(Lauraceae)樟屬(Cinnamomum)常綠喬木,樹體高大,可達50 m;葉卵狀橢圓形,離基三出脈;圓錐花序;核果;花期5月, 果期9—11月(陳有民,2010)。香樟的研究大多集中在北引抗寒生理方面,自然降溫脅迫中5種樟樹抗寒性強弱為猴樟>芳樟>本樟>尾葉樟>沉水樟(王寧等,2014),用質量濃度為500 mg·L-1的多效唑(PP333)溶液噴布葉片能明顯提高香樟幼樹的抗寒性(尤揚等,2009)。鹽脅迫方面,SNP的加入對酸鋁脅迫下的香樟幼苗起到一定緩解作用且濃度在0.05~0.1 mmol·L-1之間緩解作用最大(李茹等,2017)。分子生物學方面,CcCBFa和CcCBFb均能被低溫、干旱、鹽及ABA強烈誘導(李勇鵬等,2016)。凋落物作用方面,香樟凋落葉分解初期可能釋放了不利于土壤硝化過程的物質,造成土壤硝態氮匱乏(陳洪等,2016)。有關人工低溫脅迫下香樟葉肉細胞中草酸鈣結晶偶有報道(尤揚等,2018a,b),而香樟不同季節葉肉細胞中含晶細胞的研究未見報道。香樟葉肉細胞中草酸鈣結晶可能在植物抗逆性,尤其是在抗低溫脅迫方面有某種重要的暗示作用。
古有“樟不過江”的說法,如今,香樟的引種栽培已到達黃河南岸的鄭州、三門峽等地,甚至北上到了黃河北岸的新鄉。本研究以北引至黃河北岸的河南新鄉多年生香樟為材料,在自然狀況下,于不同季節分別對葉片取樣,用透射電子顯微鏡拍照,研究其葉肉含晶細胞在一年之中不同季節的變化,以期在細胞學水平上闡明香樟葉肉含晶細胞中草酸鈣的變化規律,為香樟的進一步北引及合理栽培奠定細胞學基礎,也為研究香樟在北方栽培環境中季節變化對草酸鈣結晶形態規律的影響提供一定的理論依據。
1 材料與方法
1.1 儀器和藥品
超凈工作臺、LEICA EM UC7超薄切片機、HITACHI-HT7700型透射電子顯微鏡;4%戊二醛(pH為7.0)、pH為7.0的0.1 mol·L-1的磷酸緩沖液(PBS)、1%四氧化鋨固定液、梯度乙醇(50%、70%、80%、90%、95%、100%)、Epon812樹脂、醋酸雙氧鈾染色液、檸檬酸鉛染色液等。
1.2 材料和處理方法
材料為從河南信陽引種到河南新鄉的多年生香樟截干苗木,干高3~3.5 m,胸徑5~8 cm,作為行道樹栽植于學校主干道兩側3 a。栽植地土壤堿解氮含量76.4 mg·kg-1,速效磷含量24.2 mg·kg-1,速效鉀含量162.5 mg·kg-1,有機質含量0.68%,土壤容重2.17 g·cm-3,土壤pH值8.2~8.3。
參照李和平(2009)和尤揚等(2018b)的方法(有改動)制備超薄切片。2019年春季[3月20日,15 ℃(取樣瞬時溫度),下同],當季平均溫度為12 ℃),夏季(6月20日,29 ℃,當季平均溫度為26.3 ℃),秋季(10月20日,20 ℃,當季平均溫度為22.6 ℃)和2020年冬季(1月5日,-6 ℃,當季平均溫度為9.3 ℃)的清晨7:00左右,隨機選取三株香樟,每株香樟分別選3個正常的、當年生且南向外圍枝條,取樣于枝條頂端向下數第3片或第四片功能葉中脈中部左右0.5 cm處,橫切(春季取樣為新生葉片,略紅色),分別用4%戊二醛進行前固定和1%四氧化鋨進行后固定。經磷酸緩沖液漂洗、脫水、環氧樹脂Epon812滲透、包埋、聚合后,用LEICA EMUC7型超薄切片機鉆石刀切片(厚度50~70 nm),經檸檬酸鉛-醋酸雙氧鈾雙重染色,用HITACHI-HT7700型透射電子顯微鏡拍照,每個超薄切片樣本拍照15個。
2 結果與分析
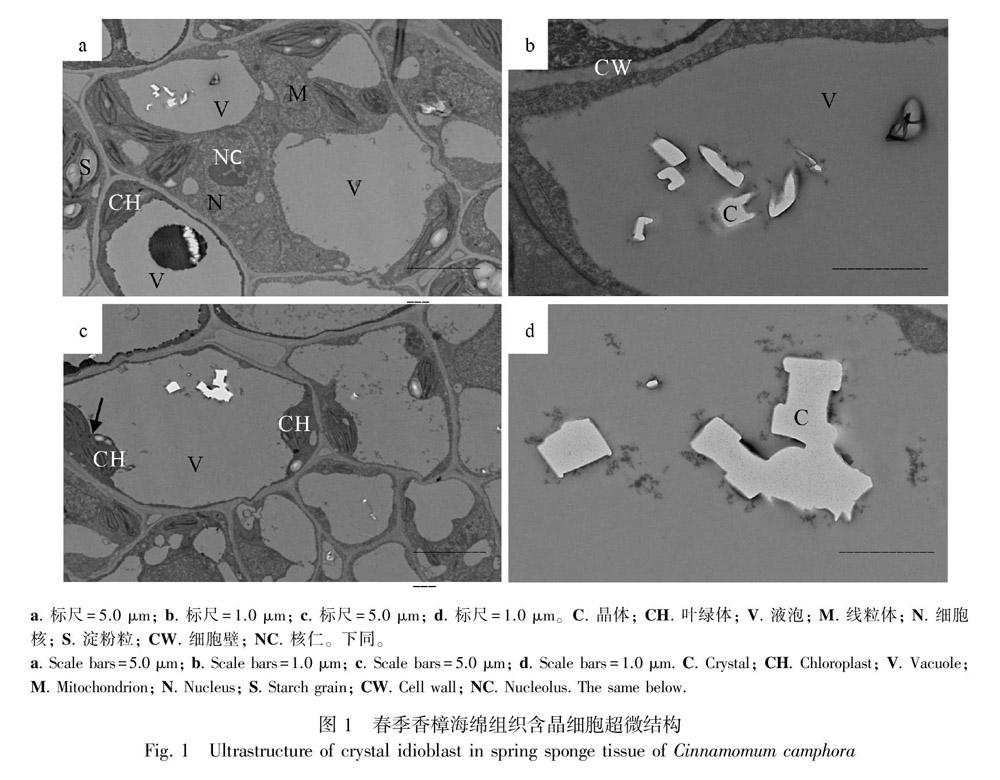
2.1 春季香樟葉肉含晶細胞的超微結構
如圖1所示,所切部位為葉肉海綿組織,春季香樟葉肉細胞的細胞壁與質膜清晰可見。細胞核為圓球形,核仁清晰地位于細胞核的中央(圖1:a)。葉綠體沿細胞質膜內側分布,基粒片層和基質片層清晰可辨,由于是春季剛萌發的新葉,可以看到葉綠體的縊縮增殖現象(圖1:c箭頭所示),觀察視野中單個細胞葉綠體的數量為2~10個不等。葉綠體縊縮增殖的生理意義在于增加了葉綠體的數量,為更多光合產物的形成提供基礎保證。部分葉綠體中含有少許淀粉粒(圖1:a,c)。線粒體呈圓球形毗鄰于葉綠體。部分細胞的中央大液泡尚未形成(圖1:a),這表明細胞還未發育成熟;而部分細胞則已經發育成熟,液泡把細胞器擠向細胞的邊緣(圖1:c)。有少許電子密度較低的草酸鈣晶體零星隨機散布于液泡中,這些晶體從觀察視野來看為針晶、方晶或柱狀晶等(圖1:b,d分別為圖1:a,c的細部特寫照片)。
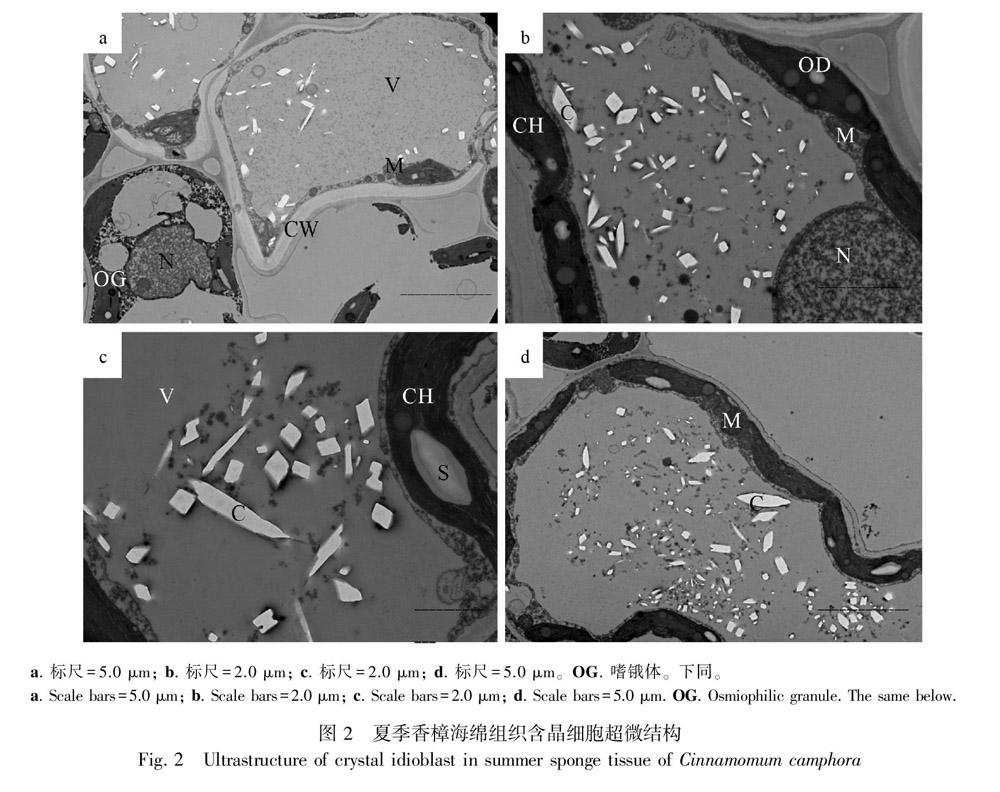
2.2 夏季香樟葉肉含晶細胞的超微結構
夏季香樟含晶細胞的超微結構所切部位是海綿組織,細胞質均勻(圖2:a)。細胞核為圓球形,由于中央大液泡的擠壓,故分布于細胞的邊緣,核質均勻,核仁明顯(圖2:a,b)。葉綠體呈“鐵餅”狀分布于細胞質膜內側,電子密度較高,為黑色;葉綠體上有電子密度較低的白色淀粉顆粒及黑色的油滴(圖2:b,c);觀察視野中單個細胞中葉綠體的數量為1~12個。線粒體緊鄰葉綠體沿細胞壁的內側分布,呈長橢圓形(圖2:b)和球形(圖2:d),內嵴明顯。中央大液泡已經形成,表明細胞已經發育成熟,占據細胞空間較大。液泡中明顯可見許多電子密度較低的白色草酸鈣晶體結構,這些晶體結構數量多、形態不一、大小不等,且隨機散布于液泡中,從觀察結果來看多為柱狀晶、方晶、針晶、簇晶。這些葉肉細胞的晶體數量遠比春季葉肉細胞中晶體數量要多,形態要豐富。
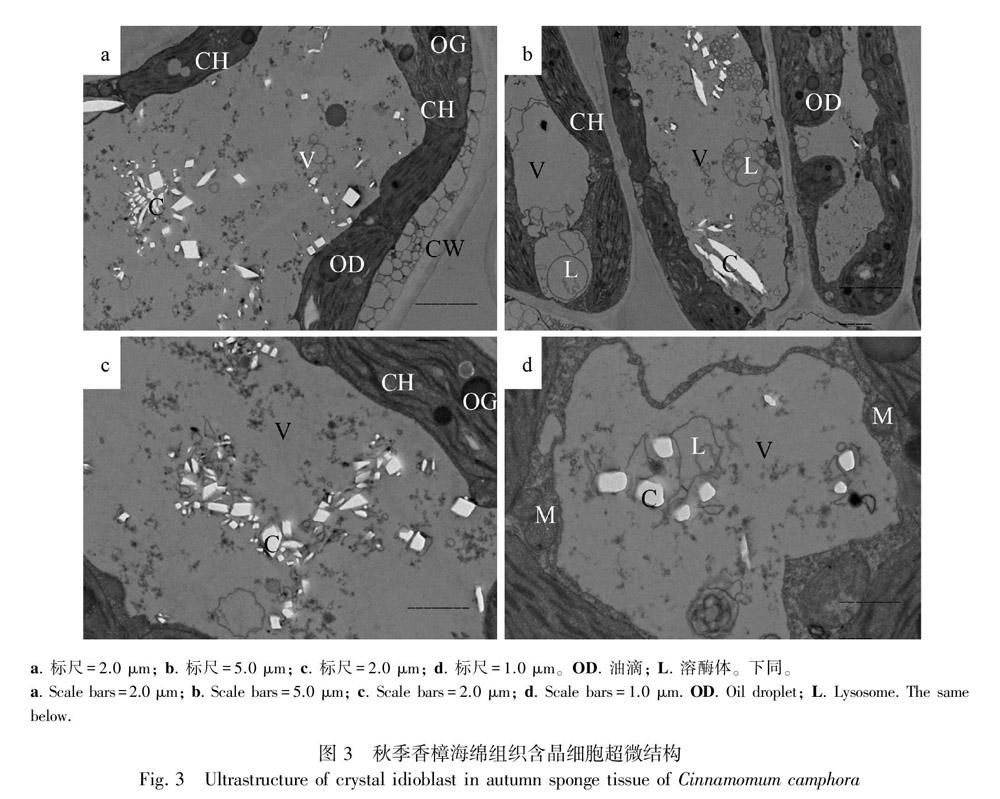
2.3 秋季香樟葉肉含晶細胞的超微結構
從圖3可看出,所切部位為葉肉海綿組織(圖3:a,c,d)和柵欄組織(圖3:b)。秋季香樟葉肉的細胞壁結構完整,層次清晰(圖3:a,b)。葉綠體基本沿著細胞壁的內側分布于細胞的邊緣,其結構完整,基粒片層和基質片層清晰。部分葉綠體有腫脹甚至破損的現象,這可能是由于早秋的低溫所致。部分葉綠體與細胞壁之間有類似“墊襯物”存在(這些“墊襯物”有什么生理意義還不甚明了,有待進一步研究),并且電子密度較低(圖3:a),這種現象在春季和夏季葉肉細胞中均沒有發現。葉綠體上有大量的球形油滴和電子密度較高的黑色嗜鋨體(圖3:a,b,c)。線粒體圓球狀數量較多,集中分布于葉綠體的附近(圖3:b,d)。這些隨機散布于中央大液泡中電子密度較低的白色草酸鈣晶體盡管數量較夏季有了一定的減少,但比春季的晶體數量要多。從圖3還可看出,這些晶體多數成團聚集在一起,有部分隨機分布于液泡中。液泡中的細胞質較濃,有大量溶酶體分布(圖3:b,d)。
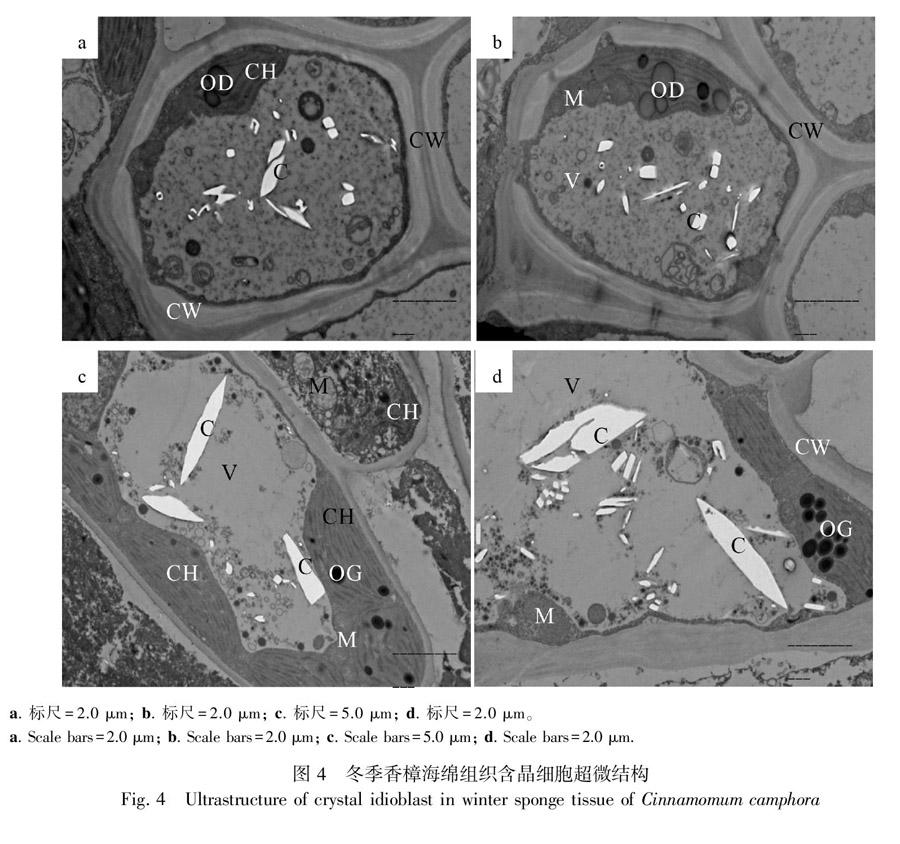
2.4 冬季香樟葉肉含晶細胞的超微結構
如圖4所示,所切部位是導管(圖4:a,b)、柵欄組織(圖4:c)和海綿組織(圖4:d)細胞,冬季香樟葉肉細胞結構完整。導管細胞細胞壁的層次清晰,可見胞間層、初生壁、次生壁(圖4:a,b)。由于次生壁的增厚不均勻,因此形成了紋孔(圖4:b),在細胞壁上還有胞間連絲(圖4:b)。部分葉綠體腫脹呈半球形,甚至有部分葉綠體崩潰解體(圖4:c)。之所以導管中的葉綠體數量較少,而海綿組織和柵欄組織的數量較多,這可能是由于導管細胞是一種逐步成熟趨死的細胞,在成熟的導管形成之前,導管內的部分細胞器逐漸減少直至消失的緣故。電子密度較高的黑色嗜鋨體和球形油滴大量聚集在葉綠體上(圖4:a-d)。圓球形的線粒體聚集在葉綠體的附近(圖4:b-d),其生理意義在于冬季低溫時,由于產能部位的線粒體和需能部位的葉綠體之間近,因此縮短了能量運輸的距離,減少了能量的損耗和運輸時間,從而有利于抵御外界的低溫。導管細胞中線粒體的數量較少,而柵欄組織和海綿組織的線粒體數量較多,呈聚集狀。導管細胞中細胞質較濃,液泡電子密度較高,呈淺黑色。中央液泡中隨機分布有電子密度較低的白色草酸鈣晶體,其形狀多樣,大小不等,多為針晶、方晶、柱狀晶。
3 討論與結論
本研究結果發現,一年四季香樟葉肉細胞液泡中均有草酸鈣晶體結構,這些晶體多為柱狀晶、方晶、針晶、晶簇。就春季而言,此時期的草酸鈣晶體只存在于少數細胞中,數量較少,晶體結構多為柱狀晶、方晶。觀察視野中只有少數細胞有草酸鈣晶體結構,推測這可能是春季正處在香樟葉片發育時期,葉綠體尚未發育成熟,部分葉綠體正在通過縊縮增殖;眾多小液泡的彼此融合逐漸形成中央大液泡,而草酸鈣晶體結構的形成尚需要一個過程。春季葉綠體的縊縮增殖增加了葉綠體的數量,為積累更多的光合產物提供了基礎保障。從夏季香樟葉肉細胞草酸鈣晶體結構和春季比較來看,夏季時葉肉細胞晶體明顯比春季的數量要多、體積要大、形態要豐富。這可能是夏季香樟葉片發育階段完成(為功能葉),草酸鈣晶體在液泡內的形成也已完成。秋季香樟葉肉細胞草酸鈣晶體結構多為柱狀晶、方晶、針晶和晶簇,和夏季的類似,比春季的數量多、體積大,多團聚在一起;秋季香樟葉肉細胞在細胞壁和葉綠體之間有1~4層大小不等的“墊襯物”存在,這些“墊襯物”電子密度較小,呈白色,在其他季節中并未發現。“墊襯物”出現在秋季有什么樣的生理意義,目前還不清楚,推測是秋季氣溫逐步降低,細胞壁內側出現的這些“墊襯物”對抵御外界的低溫脅迫,以及保護細胞內部細胞器多了一層保護“裝置”。冬季時香樟葉肉含晶細胞中細胞質較濃;部分葉綠體有腫脹崩潰現象,葉綠體上富集了電子密度較高呈黑色的油滴和嗜鋨體;線粒體毗鄰葉綠體呈球形,數量較多,且形態變化不明顯;晶體形態為柱狀晶、方晶、針晶,數量比夏季和秋季略有減少。晶體數量的減少可能是冬季低溫條件下,與草酸鈣晶體的分解有一定關聯。草酸鈣晶體分解的生理意義在于提高了香樟葉肉細胞的滲透勢,這對于降低細胞內溶液的冰點有益,進而有利于香樟葉片抵御外界的低溫傷害。草酸鈣晶體不僅對香樟抵御低溫有益,而且還可能會提高堇菜(Viola verecumda)忍耐干旱脅迫的能力(蘇志孟等,2019)。徐靜靜等(2012)對天津鹽漬化生境54種植物鈣晶體的研究表明草酸鈣晶體在落葉喬木、灌木抵御鹽分脅迫中發揮著重要作用。越來越多的文獻表明,草酸鈣在緩解植物逆境脅迫方面的確有一定的作用。
香樟在遭受外界人工低溫脅迫時,隨著脅迫溫度的逐步降低,草酸鈣晶體的數量會逐步減少,體積也隨之變小,晶體類型也由柱狀晶、方晶轉變成針晶(尤揚等,2018a)。而本研究結果發現,在觀察視野內,在一年四季中均有草酸鈣晶體隨機分布于香樟葉肉細胞的液泡中,并且冬季草酸鈣晶體的體積與夏季、秋季相比,并未減少許多,以方晶、柱狀晶為主,伴以少量針晶。這可能是由于人工低溫脅迫和自然降溫的冬季低溫脅迫有一定的差異而造成。
參考文獻:
BORCHERT R, 1986. Calcium acetate induces calcium uptake and formation of calcium-oxalate crystals in isolated leaflets of Gleditsia triacanthos L.? [J]. Planta, 168: 571-578.
CHEN H, MA GL, WANG GJ, et al., 2016. Limitation of allelopathic Cinnamomum camphora leaf litter on nitrogen nutrition in Capsicum annuum and soil [J]. Acta Bot Boreal-Occident Sin, 36(1): 106-115.? [陳洪, 馬光良, 王光劍, 等, 2016. 香樟凋落葉分解對辣椒及土壤氮營養的限制作用? [J]. 西北植物學報, 36(1): 106-115.]
CHEN YM, 2010. Landscape dendrology? [M]. Beijng: Forestry Press of China: 371-372. [陳有民, 2010. 園林樹木學? [M]. 北京: 中國林業出版社: 371-372.]
FINLEY DS, 1999. Patterns of calcium oxalate crystals in young tropical leaves: A possible role as an anti-herbivory defense? [J]. Rev Biol Trop, 47: 27-31.
FRANCESCHI VR, NAKATA PA, 2005. Calcium oxalatein plant: Formation and function [J]. Ann Rev Plant Biol, 56(1): 41-71.
HUDGIN JW, KREKLING T, FRANCESCHI VR, 2003. Distribution of calcium oxalate crystals in the secondary phloem of conifers: A constitutive defense mechanism? [J]. New Phytol, 159 (3): 677-690.
LERSTEN NR, HORNER HT, 2006. Crystal macropattem development in Prunus serotina (Rosaceae, Prunoideae) leaves? [J]. Ann Bot, 97(5): 723-729.
LI HP, 2009. Technique of microscopic plant? [M]. 2nd ed. Beijing: Science Press.? [李和平, 2009. 植物顯微技術 [M]. 2版. 北京: 科學出版社. ]
LI R, WEI J, LI TZ, et al., 2017. Effects of exogenous NO Cinnamomum camphora seedlings under acid-aluminum stresses? [J]. J NW For Univ, 32(1): 30-36.? [李茹, 韋潔, 李桃禎, 等, 2017. NO對酸鋁脅迫下香樟根系形態及葉綠素熒光特性研究? [J]. 西北林學院學報, 32(1): 30-36.]
LI YP, ZHANG LW, ZHANG JJ, et al., 2016. Cloning and expression profilling of CcCBF genes in Cinnamomum camphora? [J]. J NE For Univ, 44(8): 34-40.? [李勇鵬, 張力維, 張佳佳, 等,? 2016. 香樟CcCBFs 基因的克隆及表達模式? [J]. 東北林業大學學報, 44(8): 34-40.]
NAKATA PA, KOSTMAN TA, FRANCESCHI V R, 2003. Calreticulin is enriched in the crystal idioblasts of Pistia stratiotes [J]. Plant Physiol Biochem, 41: 425-430.
NAKATA PA, MCCOONN MM, 2003. Calcium oxalate crystal formation is not essential for growth of Medicago truncatula [J]. Plant Physiol Biochem, 41: 325-329.
SU ZM, ZHANG XM, MA L, et al., 2019. Relationship between calcium oxalate crystals and water maintenance in leaves of Viola verecumda [J]. Guihaia, 39(6): 720-728.? [蘇志孟, 張習敏, 馬琳, 等, 2019. 堇菜葉片草酸鈣晶體與水分維持的關系 [J]. 廣西植物, 39(6): 720-728.]
VOLK GM, LYNCH-HOLM VJ, KOSTMAN TA, et al., 2002. Therole of druse and raphide calcium oxalate crystals in tissue calcium regulation in Pistia stratiotes leaves [J]. Plant Biol, 4: 34-45.
WANG N, YAO F, YUAN ML, et al., 2014. Analysis on cold tolerance of five camphor tree species during the natural drop in temperature process [J]. J Henan Agric Univ, 48(3): 302-309. [王寧, 姚方, 袁美麗, 等, 2014. 自然降溫過程中5種樟樹的抗寒性分析 [J]. 河南農業大學學報, 48(3): 302-309.]
XU JJ, CI HC, HE XD, et al., 2012. The calcium crystals and calcium component characteristics of 54 plants in salinization habitat in Tianjin [J]. Chin J Appl Ecol, 23(5): 1247-1253. [徐靜靜, 慈華聰, 何興東, 等, 2012. 天津鹽澤化生境54種植物鈣晶體與鈣組分特征 [J]. 應用生態學報, 23(5): 1247-1253.]
YAN QD, SU PX, 2006. Crystal idioblasts in plant: A review of their structure and function [J]. Plant Physiol Comm, 42(4): 761-766 . [嚴巧娣,? 蘇培璽, 2006. 植物含晶細胞的結構與功能 [J]. 植物生理學通訊, 42(4): 761-766.]
YOU Y, CHEN SJ, ZHANG XY, et al., 2018a. Effects of low temperature on the mesophyal crystal idioblasts and lysosomes of Cinnamomum camphora [J]. Plant Physiol J, 54(7): 1239-1244.? [尤揚, 陳仕均, 張曉云, 等, 2018a. 低溫對香樟葉肉含晶細胞和溶酶體的影響 [J]. 植物生理學報, 54(7): 1239-1244.]
YOU Y, JIA WQ, ZHANG XY, et al., 2018b. Effect of low-temperature stress on the ultrasturcture of mesophyll cells in Cinnamomum camphore [J]. J Chin Electr Microsc Soc, 37(3): 282-288. [尤揚, 賈文慶, 張曉云, 等, 2018b. 低溫脅迫對香樟葉肉細胞超微結構的影響 [J]. 電子顯微學報, 37 (3): 282-288.]
YOU Y, YUAN ZL, WU RS, et al., 2009. Effect on spraying leaves with PP333 on cold resistance in sapling of Cinnamomum camphora [J]. Henan Sci, 27(2): 169-171. [尤揚,? 袁志良,? 吳榮升,? 等,? 2009. 葉面噴施PP333對香樟幼樹抗寒性的影響 [J]. 河南科學,? 27(2): 169-171.]
(責任編輯 蔣巧媛)
