hsa_circ_087631在原發性膽汁性膽管炎中的表達和作用機制*
2021-01-06 08:26:52張吟眉郝新杰鄭佳佳
中國病理生理雜志 2020年12期
關鍵詞:檢測
張吟眉,郝新杰,鄭佳佳
(北京大學第三醫院檢驗科,北京 100191)
原發性膽汁性膽管炎(primary biliary cholangi?tis,PBC)是一種漸進性發展的慢性自身免疫性膽汁淤積性肝病,以淋巴細胞性膽管炎,肝內小膽管逐漸破壞,導致纖維化為主要特征,并可能通過并發癥導致肝硬化[1]。PBC 發病機制復雜,目前臨床尚缺乏PBC 早期診斷理想的分子標志物以及監測PBC 疾病進展以及治療效果的有效指標[2-3]。
環狀RNA(circular RNA,circRNA)是一類共價閉合的非編碼RNA 分子,具有較高的核糖核酸酶R(ribonuclease R,RNase R)抗性和較長的半衰期,并以組織和發育階段特異性方式表達[4-5]。circRNA 具有生物調控功能,可以作為微小RNA(microRNA,miRNA,miR)的分子海綿以調控miRNA 對靶基因表達的干預,是一種潛在的理想的疾病診斷標志物和治療靶標[6-7]。我們前期分析了PBC 患者circRNA 表達譜芯片,發現22 條差異表達的circRNA,hsa_circ_087631 是其中豐度較高且表達均一的circRNA 之一[8]。通過芯片生物信息學分析獲得與circRNA 結合的miRNA,并對miRNA 靶基因進行預測,顯示在hsa_circ_087631 與miR-346 上存在結合位點。為揭示hsa_circ_087631 和miR-346 是否參與PBC 的發病機制,我們觀察了hsa_circ_087631 在PBC 患者中的表達,并初步探討了其作用機制。
前期我們通過文獻檢索聯合芯片和RT-qPCR 檢測,證實了miR-346在PBC 患者外周血T淋巴細胞的表達顯著低于健康對照組。有研究表明,濾泡輔助性T 細胞(follicular helper T cells,Tfh)和濾泡調節性T細胞(follicular regulatory T cells,Tfr)的比例失衡可以導致多種自身免疫性疾病[9-10]。本課題組的前期研究發現,PBC患者相對于健康者Tfh細胞比例顯著升高,Tfr 細胞比例下降[11]。有文獻報道,Tfh17 和Tfh2 能夠通過分泌白細胞介素21(interleukin-21,IL-21)輔助B 細胞產生免疫球蛋白[12];又有文獻表明,miR-346能通過抑制Bcl-6,調控自身免疫性疾病中循環CD4+CXCR5+Tfh 細胞的比例[13]。因此,我們試圖研究在PBC 患者中是否存在hsa_circ_087631/miR-346/Bcl6 或hsa_circ_087631/miR-346/IL-21 的 信號轉導,通過調控CD4+CXCR5+Tfh 細胞參與PBC 的進展,以及對預測的circRNA-miRNA-mRNA 通路的潛在功能進行探索,為探索PBC 新的靶向治療方法提供依據。
材料和方法
1 隊列收集
實驗對象為2018 年~2019 年北京大學第三醫院消化科門診和住院PBC 患者30 例,以及性別、年齡匹配的健康對照者30 例。所有入組個體均進行肝功能、抗線粒體抗體(antimitochondrial antibodies,AMA)、免疫球蛋白及肝臟B 超檢測,并排除HBV 和HCV 感染,隊列的臨床特征見表1。所有患者及健康對照者全部填寫知情同意書。所有個體采集外周靜脈抗凝血進行后續檢測。
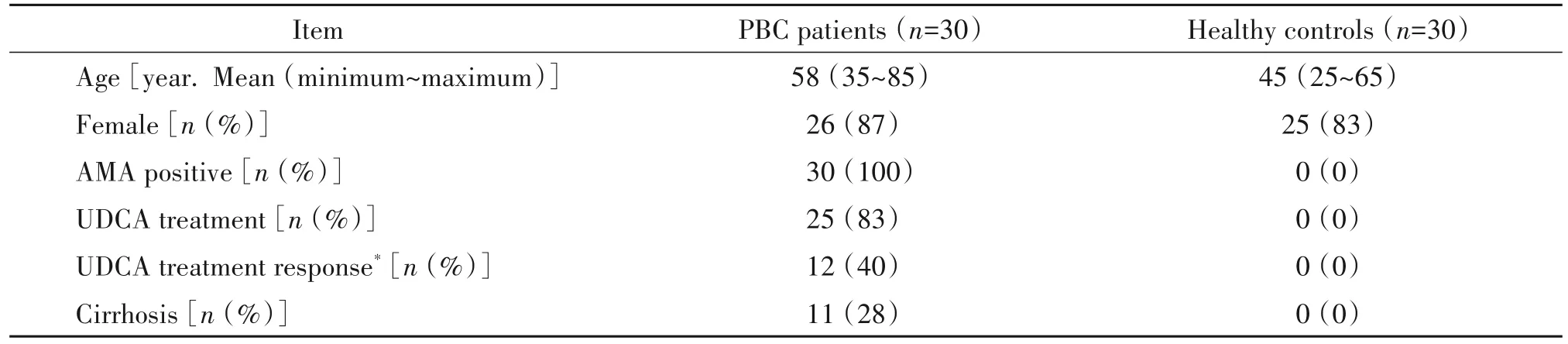
表1 PBC患者和健康對照者的臨床特征Table 1.Clinical characteristics of PBC patients and healthy controls
2 主要試劑
TRIzol 和Lipofectamine 2000(Invitrogen);逆轉錄試劑盒、SYBR Green Mix、SDS loading buffer 和RNase free water(TaKaRa);DMEM/F12培養基、胎牛血清、Opti-MEM?I Reduced Serum Medium 和0.25%EDTA-胰蛋白酶(Gibco);Polyethylenimine MAX(Polysciences);雙螢光素酶報告基因檢測試劑盒(Promega);所用引物由上海生工公司合成,序列見表2。

表2 RT-qPCR引物序列Table 2.The sequences of the primers for RT-qPCR
3 主要方法
3.1 構建過表達hsa-miR-346 的慢病毒pLenti-EF1a-EGFP-F2A-Puro-CMV-MCS 將hsa-miR-346 模板DNA 定向插入慢病毒pLenti-EF1a-EGFP-F2A-Pu?ro-CMV-MCS 載體(記為H145)中,構建過表達hsamiR-346 的慢病毒 pLenti-EF1a-EGFP-F2A-Puro-CMV-MCS(記為H9113),并轉染到293TN細胞中。
3.2 RT-qPCR 實驗 采用TRIzol 法從細胞中提取總RNA,取1.0 μg 總RNA,用oligo(dT)和random primers 逆轉錄出cDNA 用于檢測hsa-miR-346、hsa_circ_087631、Bcl-6 mRNA 和IL-21 mRNA 表達水平。以GAPDH 為內參照。在ABI PRISM?7500 Se?quence Detection System(Applied Biosystems)進行PCR和數據分析。以2?ΔΔCt法計算相對表達水平
3.3 雙螢光素酶報告基因系統檢測hsa_circ_087631 與hsa-miR-346 的結合 參照已報道方法構建包含hsa-miR-346 潛在結合序列的hsa_circ_087631 重組螢光素酶報告質粒pGL3-hsa_circ_087631-761-781 及包含突變了結合序列的重組質粒pGL3-hsa_circ_087631-761-781-Mut。將293T細胞按70%的匯合度接種到96 孔板,24 h 后轉染螢光素酶報告基因質粒和miRNA,每個樣品設置6 個復孔,轉染48 h后,棄去培養基,用PBS緩沖液洗滌細胞1次。每孔加入50 μL 的1× Passive Lysis Buffer 裂解液,常溫條件搖床晃動15 min;裂解液轉移到白色不透光的96 孔酶標板中,加入100 μL 預先混好的LAR II,設置熒光發光儀程序,檢測螢火蟲螢光素酶的活性;向檢測管中加入100 μL Stop&Glo?試劑,檢測海腎螢光素酶活性。兩種螢光素酶活性比值的變化可反映hsa_circRNA_08763與hsa-miR-346結合情況。
4 統計學處理
計量資料以均數±標準差(mean±SD)表示,用SPSS 20.0 統計軟件進行分析。多組間均數比較采用單因素方差分析(one-way ANOVA),組間兩兩比較采用Bonferroni 校正的t檢驗。以P<0.05 為差異有統計學意義。
結 果
1 PBC患者外周血hsa_circ_087631水平顯著升高
本研究選取30 例PBC 患者和30 例健康對照者血漿標本進行檢測,結果顯示,PBC 患者的hsa_circ_087631 水平與健康對照者相比顯著升高(P<0.05),見圖1。
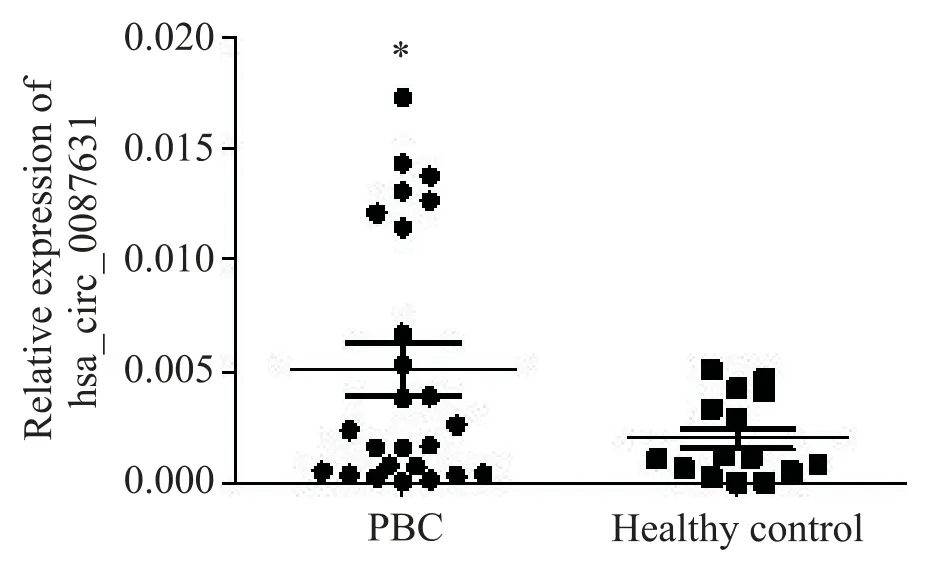
Figure 1.The expression of hsa_circ_0087631 in the peripheral blood from PBC patients and healthy controls was de?tected by RT-qPCR.Mean±SD. n=30.*P<0.05 vs healthy control group.圖1 PBC 患者和健康對照者外周血hsa_circ_0087631 表達水平的比較
2 hsa-miR-346 在Jurkat 細胞過表達后hsa_circ_0087631表達降低
通過芯片生物信息學分析,我們發現在PBC 外周血中高表達的hsa_circRNA_087631 與miR-346 存在結合位點。接下來我們驗證兩者是否存在調控作用。將目的基因hsa-miR-346 插入空載體pLenti-EF1a-EGFP-F2A-Puro-CMV-MCS(H145),成功構建hsa-miR-346 過表達的慢病毒pLenti-EF1a-EGFPF2A-Puro-CMV-MCS(H9113),見圖2A。該慢病毒感染293TN 細胞的效果如2B 所示,H9113 能夠穩定過表達hsa-miR-346,見圖2C。將該慢病毒以MOI=60感染人急性T細胞白血病Jurkat細胞后,與空載體對照Jurkat-H145 組相比,轉染H9113 的Jurkat-H9113 組中hsa-miR-346 的表達水平顯著增高(P<0.01),而hsa_circ_087631 的表達水平降低(P<0.01),見圖2D、E。
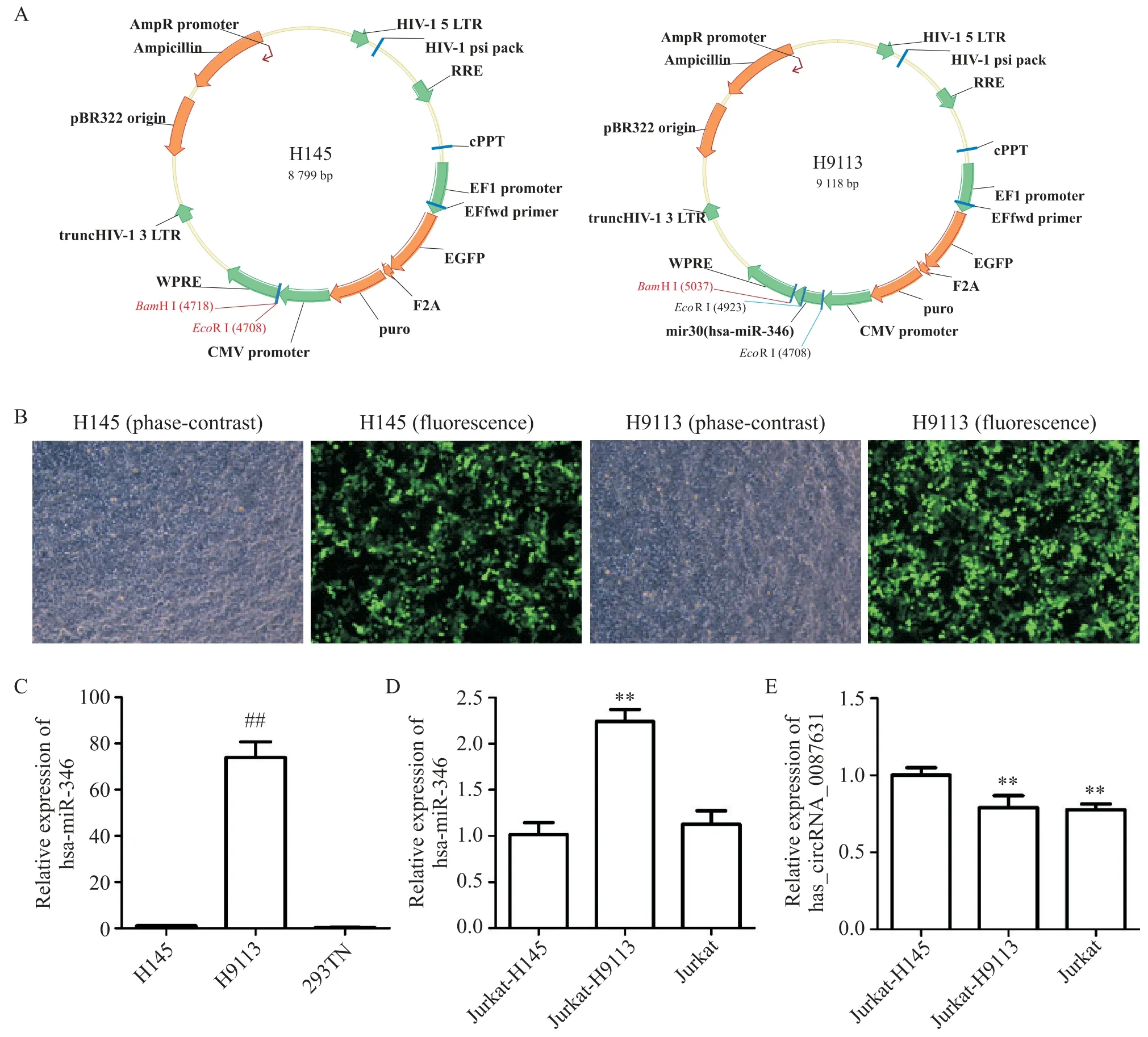
Figure 2.The expression of hsa_circ_0087631 was decreased after the hsa-miR-346-overexpressing lentiviral vector was transfected into the Jurkat cells.A:the structural diagram of blank vector pLenti-EF1a-EGFP-F2A-Puro-CMV-MCS(H145)and hsamiR-346-overexpressing lentiviral vector pLenti-EF1a-EGFP-F2A-Puro-CMV-MCS(H9113);B:the cell infection rate of H145 and H9113(48 h after transfection,×200);C:the expression of hsa-miR-346 in 293TN cells after transfection;D:the expression of hsa-miR-346 after the hsa-miR-346-overexpressing lentiviral vector H9113 was transfected into the Jurkat cells;E:the expression of hsa_circ_087631 after the hsa-miR-346-overexpressing lentiviral vector H9113 was transfected into the Jurkat cells.Mean±SD. n=3.##P<0.01 vs H145 group;**P<0.01 vs Jurkat-H145 group.圖2 hsa-miR-346過表達慢病毒降低Jurkat細胞中hsa_circ_0087631表達
3 hsa-miR-346對Bcl-6和IL-21的調控
我們進一步驗證miR-346 是否對下游靶基因Bcl-6和IL-21存在調控。將hsa-miR-346 過表達慢病毒質粒H9113 以MOI=60 感染Jurkat 細胞后,Jurkat-H9113 組Bcl-6 和IL-21 的mRNA 表達均顯著降低(P<0.05),見圖3。這提示hsa-miR-346 對下游基因Bcl-6和IL-21存在調控作用。
4 驗證hsa_circ_0087631與hsa-miR-346的相互作用
我們通過雙螢光素酶報告基因系統檢測hsamiR-346和hsa_circ_087631是否存在相互作用,再結合定點突變技術確定miRNA 與靶基因的作用位點。構建hsa_circ_087631 野生型質粒(pMIR-REPORThsa_circ_087631 3′UTR-WT,H7168)和hsa_circ_087631 雙突變質粒(pMIR-REPORT-hsa_circ_087631 3′UTR-Mut,H7169),其中hsa_circ_087631雙突變位點針對生物信息學預測的hsa-miR-346 與hsa_circ_087631 結合的兩個靶位點進行突變,見圖4A。將H7168 及H7169 分別與hsa-miR-346 mimics共轉染至293T 細胞,hsa-miR-346 可顯著降低野生型hsa_circ_087631 的螢光素酶活性(P<0.01);而在結合位點突變后,這種調控關系仍然存在(P<0.01),見圖4B。由此推測該miRNA 可能不是通過我們預測的結合位點調控了螢光素酶的表達。上述結果證實hsa_circ_087631 與hsa-miR-346 的確存在相互作用,hsa-miR-346 抑制了hsa_circ_087631 的表達,但結合位點需要進一步通過實驗驗證。
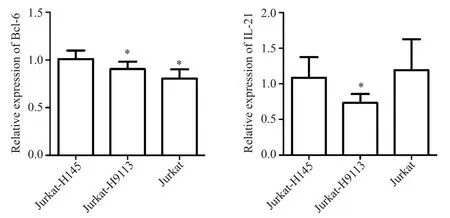
Figure 3.The mRNA expression of Bcl-6 and IL-21 after the hsa-miR-346-overexpressing lentiviral vector was transfected into the Jurkat cells.Mean±SD. n=3.*P<0.05 vs Jurcat-H145 group.圖3 hsa-miR-346過表達對Jurkat細胞中Bcl-6和IL-21表達的影響
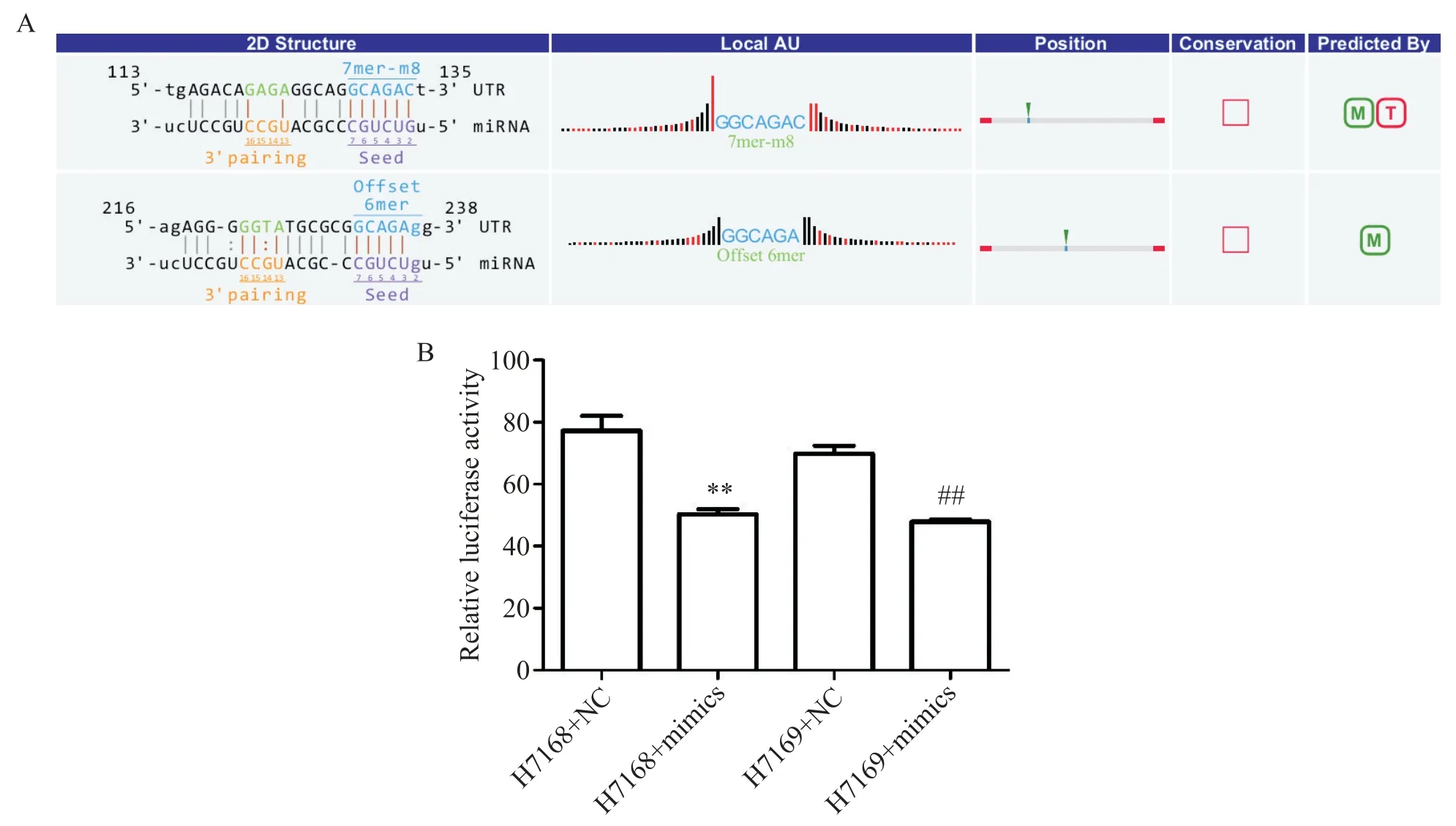
Figure 4.Dual-luciferase reporter assay was performed to identify the interactions between hsa_circ_087631 and hsa-miR-346.A:the schematic diagram of double mutant hsa_circ_087631 vector(H7169);B:relative luciferase activity in dual-luciferase reporter assay.Mean±SD. n=3.**P<0.01 vs H7168+NC group;##P<0.01 vs H7169+NC group.圖4 雙螢光素酶報告基因實驗驗證hsa_circ_0087631與hsa-miR-346的相互作用
討 論
PBC 是一種慢性膽汁淤積性自身免疫性疾病,可進展為終末期肝病,其發病機制復雜,目前臨床尚缺乏PBC 早期診斷理想的分子標志物以及監測PBC疾病進展和治療效果的有效指標[15]。
circRNA 是一類具有調控基因表達作用的特殊的內源性非編碼RNA,在進化上高度保守[5],與線性的mRNA 相比有更好的穩定性。circRNA 在外周血中穩定存在,參與免疫細胞發育和免疫調節的多個階段,不僅可以作為診斷人類自身免疫性疾病的生物標志物,還可以代表疾病的活動性或嚴重程度。miRNA 是一種內源性的小分子非編碼RNA,廣泛參與生物體的生長、凋亡等的調控,與多種自身免疫性疾病的發展密切相關,circRNA 可通過作為miRNA的分子海綿調控許多生物學過程,包括DNA 甲基化、免疫反應和炎癥反應,與多種自身免疫性疾病的相關[16-18]。
在前期工作中,我們通過比較PBC 患者及性別年齡匹配的健康對照者外周血血漿中circRNA 表達譜芯片分析以及聚類分析,發現了22 條差異表達的circRNA,其中hsa_circ_087631 是外周血中高表達的circRNA 之一。本研究中我們再次通過RT-qPCR 對芯片結果進行驗證,結果證實hsa_circ_087631 在PBC 患者外周血中相對于健康對照者表達升高。將hsa-miR-346過表達慢病毒感染人急性T細胞白血病Jurkat 細胞后,hsa_circ_087631 表達降低,提示hsa_circ_0087631 可能通過調控hsa-miR-346 發揮作用。
Tfh 細胞作為新近發現的效應T 細胞亞群,輔助生發中心B 細胞產生抗體,其調節抗體生成的促生發中心反應可被Tfr 細胞抑制。多種自身免疫病的發生與Tfr 細胞分化不足、Tfh 數量增多及Tfr/Tfh 比值失衡有關[19-20]。本課題前期研究結果發現PBC 患者相對于健康者Tfh 細胞比例顯著升高,Tfr 細胞比例下降,Tfr/Tfh比值顯著降低[11]。
Chen(陳娟)等[13,21]報道了miR-346 對Bcl-6 有調控作用,而黃勇武等[22]的研究表明IL-21 可能通過調節Th17/Treg 平衡而參與了PBC 的發病機制。IL-21 可介導多種生物學效應的細胞因子,主要由活化的CD4+T 輔助細胞等合成和分泌,同時也可促進Tfh17 和Tfh2 分化、B 細胞轉化為漿細胞以及免疫球蛋白的分泌,與多種自身免疫性疾病發病有關[22-23]。Bcl-6 主要表達于生發中心的B 細胞和CD4+T 細胞,參與調節T 細胞依賴的抗原反應。有研究顯示miR-346 在PBC 患者外周血T 細胞中顯著低于健康者,并可能參與了PBC 的發病過程[24],其機制可能是通過在轉錄水平和翻譯水平抑制Bcl-6,從而調控自身免疫病中循環CD4+CXCR5+Tfh 細胞的表達情況[13]。本研究中,將hsa-miR-346 過表達慢病毒感染Jurkat細胞后,Bcl-6和IL-21的mRNA表達均顯著降低。這提示在人T 細胞中可能存在hsa_circ_0087631/hsamiR-346/Bcl-6 或hsa_circ_0087631/hsa-miR-346/IL-21 功能關聯的調控。hsa_circ_0087631 可能通過下調hsa-miR-346,調控下游靶基因Bcl-6和IL-21,上調Tfh細胞比例,從而在PBC的發病過程中發揮作用。
接下來,我們通過雙螢光素酶報告基因系統研究hsa-miR-346 與hsa_circ_087631 的相互作用,并結合定點突變技術確定miRNA 與靶基因的作用位點。將野生型及雙突變的hsa_circ_087631 質粒分別與hsa-miR-346 mimics 共轉染至293T 細胞后,hsa-miR-346 可顯著降低野生型hsa_circ_087631 的螢光素酶活性,而在結合位點突變后,這種調控關系仍然存在。上述實驗結果證實了hsa-miR-346 與hsa_circ_087631 之間存在特異結合作用,但可能不是通過我們預測的結合位點調控螢光素酶的表達。在下一步的實驗中,我們將應用多數據庫,交叉預測hsa-miR-346 與hsa_circ_087631 之間的作用位點,繼續探明hsa_circ_087631在PBC中作用的確切分子機制。
綜上所述,本研究探索了參與自身免疫病理進程重要的Tfh 細胞相關的circRNA 在PBC 中的表達及其與上下游調控分子相互作用的機制。此類研究在國內外尚無報道,在相關研究領域屬于首創,具有一定的創新性和科學價值,為探索PBC 新的靶向治療方法提供理論依據。本研究也存在一定局限性,circRNA 可能影響的潛在下游靶基因對細胞功能的作用以及circRNA 與miR-346 的正確結合位點還有待進一步深入研究。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
