電場(chǎng)技術(shù)對(duì)于熱帶水果保鮮機(jī)理影響研究
2020-12-19 09:15:22左秋杰
日用電器 2020年11期
任 猛 左秋杰
(長(zhǎng)虹美菱股份有限公司 合肥 230001)
引言
前期研究發(fā)現(xiàn),電場(chǎng)條件下保鮮效果存在明顯差異,具體表現(xiàn)在:①不同電場(chǎng)條件下保鮮效果的差異;②不同代謝類型果蔬電場(chǎng)作用效果的差異,主要表現(xiàn)在呼吸躍變型和非呼吸躍變型水果。而根據(jù)文獻(xiàn)以及資料搜集,并無(wú)相關(guān)的研究分析。
果蔬作為活體,需要進(jìn)行一系列的代謝維持機(jī)體正常的生活活動(dòng),包括營(yíng)養(yǎng)物質(zhì)合成代謝和消耗代謝,而這些代謝活動(dòng)強(qiáng)弱均由相關(guān)的酶所催化。根據(jù)生物學(xué)遺傳知識(shí),酶由基因編碼控制,因此果蔬代謝的相關(guān)變化其實(shí)就是代謝途徑中相關(guān)酶活性及其含量發(fā)生變化的結(jié)果,所有的生命活動(dòng)均由基因所控制[1]。從基因?qū)用娼馕鲭妶?chǎng)處理的保鮮機(jī)理,根據(jù)電場(chǎng)處理對(duì)水果基因表達(dá)的影響進(jìn)行研究,找到電場(chǎng)處理對(duì)于熱帶水果代謝影響等更深層次的機(jī)理,能夠更好解釋電場(chǎng)對(duì)于熱帶水果作用的機(jī)理。本文通過(guò)實(shí)驗(yàn)分析不同電場(chǎng)強(qiáng)度下對(duì)于呼吸躍變型果蔬乙烯的釋放以及呼吸強(qiáng)度的影響,確定電場(chǎng)適宜的催熟處理?xiàng)l件。實(shí)現(xiàn)電場(chǎng)技術(shù)優(yōu)化的同時(shí),解決熱帶水果存儲(chǔ)難的問(wèn)題,為熱帶水果在冰箱中的保鮮提供技術(shù)以及理論支撐。
1 材料與方法
1.1 材料選擇及依據(jù)
選擇香蕉作為電場(chǎng)處理的研究材料,電場(chǎng)為高壓靜電場(chǎng),電場(chǎng)強(qiáng)度為30~200 V內(nèi)。由于香蕉作為熱帶水果,具有可靠的基因組信息資源可供利用,便于后續(xù)的基因信息挖掘。電場(chǎng)強(qiáng)度選擇根據(jù)前期保鮮效果試驗(yàn)結(jié)果確定。
1.2 試驗(yàn)設(shè)備
美菱BCD-481WQ3M冰箱2臺(tái),高壓靜電場(chǎng)模塊、氣相色譜儀等。
1.3 分析方法
測(cè)定電場(chǎng)條件下香蕉代謝基因表達(dá)差異性,結(jié)合主要代謝指標(biāo)(酶活、丙二醛、乙烯)數(shù)據(jù),分析電場(chǎng)熱帶水果保鮮的機(jī)理。
2 試驗(yàn)方法
2.1 基因分析方法
選擇香蕉作為試驗(yàn)材料,通過(guò)電場(chǎng)處理,進(jìn)行轉(zhuǎn)錄組分析。測(cè)試樣品分別存儲(chǔ)在適宜溫度(12 ℃)條件下設(shè)有電場(chǎng)模塊的冰箱以及對(duì)照冰箱。每?jī)商鞙y(cè)試乙烯釋放量,在最佳效果的時(shí)間點(diǎn)取樣(根據(jù)對(duì)照與試驗(yàn)組數(shù)據(jù)出現(xiàn)明顯差異判斷),提取樣品總RNA進(jìn)行轉(zhuǎn)錄組測(cè)序和分析,統(tǒng)計(jì)差異基因,結(jié)合代謝通路進(jìn)行分析。
2.2 酶活性測(cè)試
2.2.1 酶液的提取
稱取樣品10 g預(yù)冷研磨,加入20 mL 經(jīng)預(yù)冷處理的95 %乙醇,低溫靜置10 min后,低溫4 ℃下12 000 r/min離心20 min,去除上清液。重復(fù)上述操作后,加入5 mL預(yù)冷的1.8 mol/L NaCl提取緩沖溶液(50 mmol/L乙酸-乙酸鈉緩沖液,pH值為5.5),4 ℃條件下放置20 min,離心后收集上清液,于 4 ℃ 保存。
2.2.2 多聚半乳糖醛酸酶[2]活性測(cè)定
取2支試管,均加入1 mL 50 mmol/L,pH 5.5乙酸-乙酸鈉緩沖溶液和0.5 mL 10g/L多聚半乳糖醛酸溶液(1.0 g多聚半乳糖醛酸溶于100 mL 50 mmol/L pH 5.5乙酸-乙酸鈉緩沖溶液),1支試管加入0.5 mL酶提取液,另1支加入0.5 mL經(jīng)煮沸5 min的酶提取液作為對(duì)照并標(biāo)記,混勻后在37 ℃保溫1 h,保溫后迅速加入1.5 mL 3,5-二硝基水楊酸 (DNS),沸水浴 5 min,取出后經(jīng)流水冷卻,蒸餾水稀釋至25 mL,在540 nm處測(cè)定吸光度。計(jì)算酶活,公式如下:
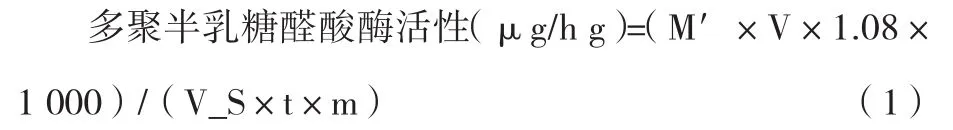
式中:
M′—從標(biāo)準(zhǔn)曲線查的葡萄糖質(zhì)量,mg;
V—樣品提取液總體積,mL;
VS—測(cè)定時(shí)索取樣品提取液體積,mL;
T—酶促反應(yīng)時(shí)間,h;
M—樣品質(zhì)量,g。
2.2.3 羧甲基纖維素酶活性測(cè)定
取2支試管,每支加入1.5 mL底物(底物配制:稱取1.0 g羧甲基纖維素鈉溶于 pH值為5.0的 50 mmol/L檸檬酸-檸檬酸鈉緩沖液中,定容到100 mL),1支試管加入0.5 mL酶提取液,另1支加入0.5 mL經(jīng)煮沸5 min的酶提取液作為對(duì)照,混勻后在37 ℃保溫1 h,保溫后迅速加入1.5 mL 3,5-二硝基水楊酸(DNS),沸水浴 5 min,取出后經(jīng)流水冷卻,蒸餾水稀釋至25 mL,在540 nm處測(cè)定吸光度。
計(jì)算公式:
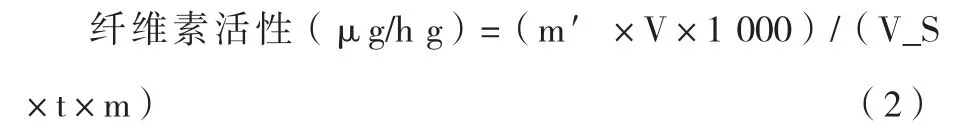
式中:
m′—從標(biāo)準(zhǔn)曲線查的葡萄糖質(zhì)量,mg;
V—樣品提取液總體積,mL;
VS—測(cè)定時(shí)索取樣品提取液體積,mL;
T—酶促反應(yīng)時(shí)間,h;
M—樣品質(zhì)量,g。
3 結(jié)果與分析
3.1 測(cè)序樣品乙烯釋放量的測(cè)定
從表1中可以看出,香蕉從第4 天開始乙烯釋放量開始出現(xiàn)差異,第6天的差異比較顯著,顯示了通過(guò)抑制乙烯達(dá)到保鮮的效果,由此選擇最佳電場(chǎng)強(qiáng)度處理6 d的香蕉作為測(cè)序的樣品。
3.2 香蕉Unigene功能注釋及相關(guān)性分析
根據(jù)測(cè)試分析,71 139個(gè)unigenes被成功注釋。Unigenes有48.72 %與Nr數(shù)據(jù)庫(kù)一致,有30.76 %的unigenes得到GO注解,分別有20.77 %,11.88 %和31.91 %的 unigenes得到KEGG,COG,Pfam數(shù)據(jù)庫(kù)的注釋。香蕉unigenes功能注釋圖見圖1。
通過(guò)樣品之間相關(guān)性的分析(如圖2),可觀察組內(nèi)樣品之間的生物學(xué)重復(fù)性。同時(shí)組內(nèi)樣品相對(duì)組間樣品的相關(guān)系數(shù)越高,獲得的差異代謝物越可靠,且驗(yàn)證了試驗(yàn)設(shè)計(jì)的合理性。如圖3~5所示,可以看到相關(guān)系數(shù)均在0.92以上,說(shuō)明測(cè)定的樣本穩(wěn)定性較好,獲得的數(shù)據(jù)質(zhì)量高,可以進(jìn)行下一步的轉(zhuǎn)錄組測(cè)序分析。
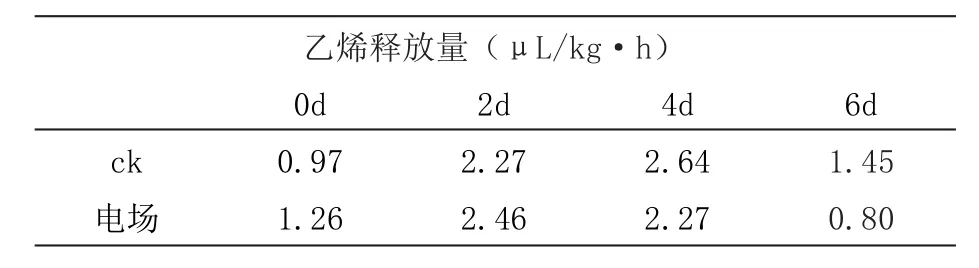
表1 香蕉在不同電場(chǎng)強(qiáng)度的乙烯釋放量
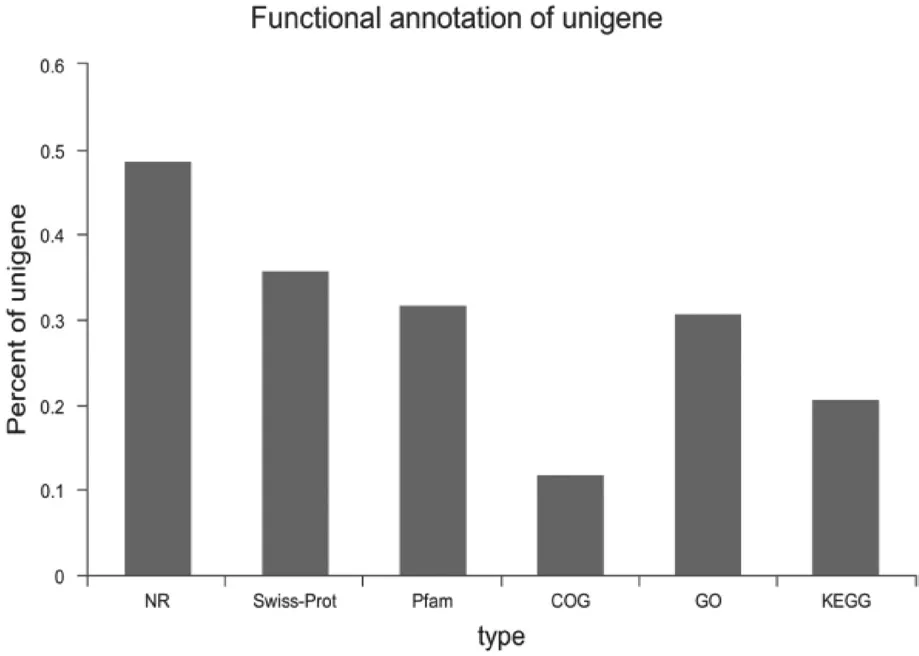
圖1 香蕉unigenes功能注釋圖
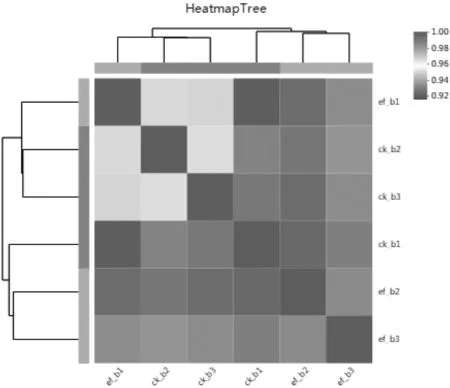
圖2 香蕉相關(guān)性分析
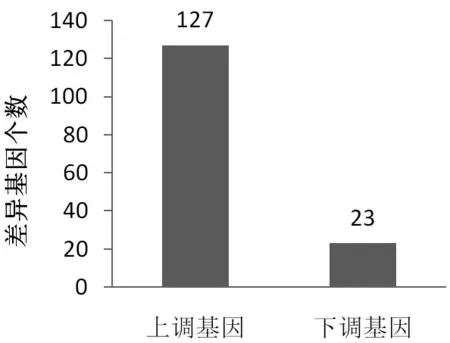
圖3 香蕉差異基因統(tǒng)計(jì)圖
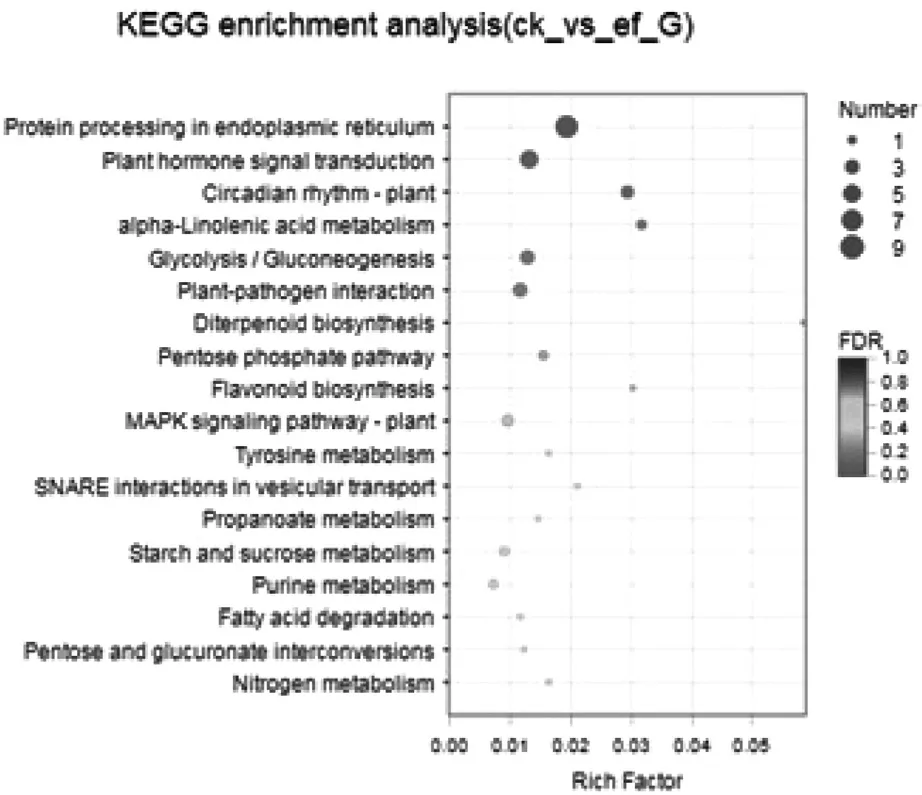
圖4 差異基因富集分析
3.3 差異基因的篩選
電場(chǎng)處理果實(shí)中有150個(gè)基因的表達(dá)水平受到顯著影響,進(jìn)一步研究發(fā)現(xiàn)這150個(gè)基因中有127個(gè)基因的表達(dá)量增加了,而剩下的23個(gè)基因表達(dá)量下降了,如圖3~5所示;進(jìn)一步分析發(fā)現(xiàn),這有150受到電場(chǎng)處理影響的基因里面有3個(gè)與乙烯有關(guān),7個(gè)與細(xì)胞壁代謝有關(guān)以及32個(gè)與細(xì)胞膜結(jié)構(gòu)和代謝有關(guān)(見表2),這些基因的表達(dá)變化會(huì)影響到電場(chǎng)處理后的果實(shí)乙烯生成情況,果實(shí)軟化和細(xì)胞膜的透性或者衰老特性。除此之外,其他受到電場(chǎng)處理的差異表達(dá)基因大多是和蛋白質(zhì)的折疊和加工相關(guān),這些蛋白與果實(shí)的保鮮關(guān)系不大。
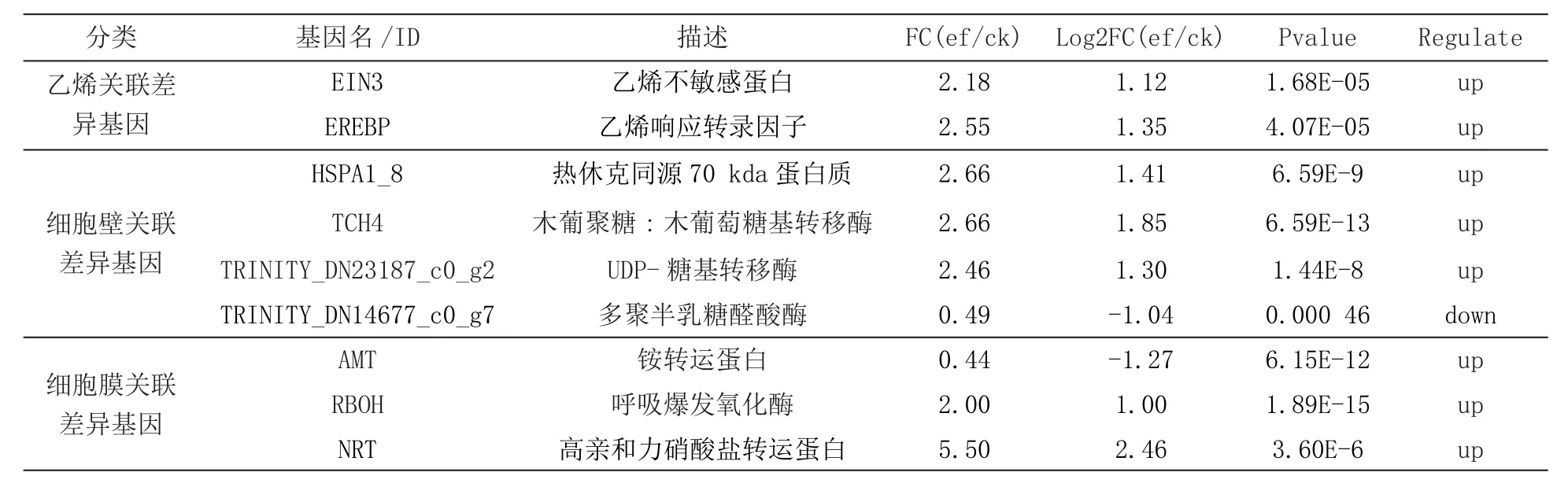
表2 與乙烯,細(xì)胞壁以及細(xì)胞膜相關(guān)的差異基因統(tǒng)計(jì)表

圖5 植物信號(hào)轉(zhuǎn)導(dǎo)代謝通路圖
考慮到上述差異基因大部分集中于生物體的蛋白質(zhì)加工和代謝過(guò)程,這些與保鮮關(guān)系不大。參考差異基因以及代謝通路,結(jié)合乙烯釋放量的差異,電場(chǎng)處理導(dǎo)致了香蕉中的乙烯信號(hào)傳導(dǎo)途徑關(guān)鍵酶編碼基因EIN3受到影響,抑制乙烯的信號(hào)轉(zhuǎn)導(dǎo)過(guò)程,阻斷乙烯在細(xì)胞間的傳遞,使乙烯不能和乙烯受體結(jié)合,也就是細(xì)胞不能感受到乙烯信號(hào),最終延緩水果的成熟衰老過(guò)程;另外,在植物激素信號(hào)轉(zhuǎn)導(dǎo)通路中發(fā)現(xiàn),細(xì)胞在油菜素類固醇正常生成后,油菜素類固醇作為一種信號(hào)調(diào)控關(guān)鍵酶基因TCH4(木葡聚糖:木葡萄糖基轉(zhuǎn)移酶)被電場(chǎng)處理后顯著上調(diào)表達(dá),也就意味著木葡聚糖:木葡萄糖基轉(zhuǎn)移酶的活性得到增強(qiáng),促進(jìn)細(xì)胞壁半纖維素類多糖—木葡聚糖的生成,促進(jìn)細(xì)胞壁的生成和活性,從而控制細(xì)胞增大,保持細(xì)胞硬度,維持水果新鮮度。電場(chǎng)施加之后,作為一種信號(hào)進(jìn)入植物MAPK信號(hào)通路中,調(diào)控信號(hào)通路中的關(guān)鍵酶的基因RBOH(呼吸爆發(fā)氧化酶)上調(diào)表達(dá),RBOH作為一種呼吸作用氧化酶,以氧氣和過(guò)氧化氫為受體,可以消耗細(xì)胞內(nèi)的氧化物質(zhì),維持細(xì)胞內(nèi)的活性氧的穩(wěn)態(tài),防止細(xì)胞氧化包括細(xì)胞膜的過(guò)氧化,維持細(xì)胞內(nèi)正常氧化還原狀態(tài),避免細(xì)胞膜的破壞,維持細(xì)胞活性,延緩了植物衰老,維持良好品質(zhì)。
3.4 生理指標(biāo)的測(cè)試
上述試驗(yàn)從基因水平解析了電場(chǎng)處理對(duì)果實(shí)保鮮相關(guān)代謝途徑中關(guān)鍵基因的影響,包括乙烯生物合成和信號(hào)傳導(dǎo),細(xì)胞壁的代謝等。生理指標(biāo)測(cè)試從細(xì)胞和蛋白水平了解電場(chǎng)處理對(duì)細(xì)胞壁代謝和細(xì)胞衰老的影響。其中細(xì)胞壁代謝影響果實(shí)的軟化,細(xì)胞電導(dǎo)率和氧化能力反應(yīng)細(xì)胞的衰老水平,也間接反應(yīng)果實(shí)的保鮮效果。其中細(xì)胞壁代謝相關(guān)酶選擇了多聚半乳糖醛酸酶和纖維素酶;細(xì)胞衰老相關(guān)指標(biāo)選擇丙二醛含量指標(biāo)測(cè)定。
由表3可以看出,香蕉在電場(chǎng)中其多聚半乳糖醛酸酶,纖維素酶活性均低于對(duì)照組香蕉相應(yīng)酶的活性,說(shuō)明30~200 V電場(chǎng)可以有效抑制這兩種細(xì)胞壁酶的活性,抑制了細(xì)胞壁的降解,延緩果實(shí)軟化;香蕉在電場(chǎng)強(qiáng)度中丙二醛含量和電導(dǎo)率也低于對(duì)照組,說(shuō)明電場(chǎng)可以有效抑制細(xì)胞膜過(guò)氧化,降低膜的通透性,保持細(xì)胞活性,延緩衰老過(guò)程,保持水果新鮮度,與轉(zhuǎn)錄組的數(shù)據(jù)分析相吻合,香蕉生理指標(biāo)測(cè)定結(jié)果均說(shuō)明電場(chǎng)對(duì)水果的細(xì)胞壁和細(xì)胞膜不產(chǎn)生傷害,維持細(xì)胞壁和細(xì)胞膜的完整性和功能,延緩果實(shí)軟化和衰老過(guò)程,保持果實(shí)新鮮。

表3 香蕉電場(chǎng)處理第6天生理指標(biāo)測(cè)定表
4 結(jié)論
綜合得知,電場(chǎng)保鮮對(duì)于水果的保鮮機(jī)理是通過(guò)抑制乙烯信號(hào)傳導(dǎo)過(guò)程關(guān)鍵基因,抑制乙烯在細(xì)胞內(nèi)的傳遞,使乙烯不能與受體結(jié)合,實(shí)現(xiàn)果蔬催熟過(guò)程抑制的同時(shí),調(diào)控細(xì)胞壁多糖代謝過(guò)程,調(diào)控與細(xì)胞壁相關(guān)酶基因,實(shí)現(xiàn)酶活性的調(diào)節(jié),具體表現(xiàn)在促進(jìn)細(xì)胞壁的生成,維持細(xì)胞活性。
