GATA4基因慢病毒載體構建與鑒定
2020-12-09 01:07:44張帆厙廣巖鐘浩馬偉宗淵武建英
山東醫藥 2020年34期
張帆,厙廣巖,鐘浩,馬偉,宗淵,武建英
1 青海大學,西寧810016;2 青海大學附屬醫院
先天性心臟病(CHD)是指在胚胎發育時期心臟和大血管形成障礙或發育異常所致的先天性畸形。目前,CHD的成因尚不明確,遺傳因素可能是其形成的重要原因,尤其是基因單核苷酸多態性(SNP)位點突變[1-3]。鋅指轉錄因子4(GATA4)是人類CHD的致病基因之一。流行病學調查發現,GATA4外顯子基因突變可引起CHD,并且其發病率在不同民族間具有明顯統計學差異[4-5]。GATA4基因H436Y位點突變可明顯提高漢族人群房間隔缺損(ASD)、室間隔缺損(VSD)的發病率[6]。在VSD患者中能夠檢測到GATA4基因p.S335X位點突變[7]。以上研究提示,GATA4基因突變與CHD的發病密切相關,但目前對其機制尚不完全清楚。2019年3月—2020年4月,本研究構建了GATA4基因慢病毒載體并轉染293T細胞,觀察了GATA4在體外的轉錄水平,為進一步闡明GATA4基因突變導致CHD的機制提供理論依據。
1 材料與方法
1.1 材料 人源胚胎腎細胞293T(以下稱293T細胞),購自上海弘順生物科技有限公司;DH5α感受態細胞,購自日本TaKaRa公司。pMGic7.1載體,購自生工生物工程(上海)股份有限公司。VSVG-GFP質粒、pCMV-dR8.9包裝質粒,購自美國Addgene公司。引物序列設計合成和陽性克隆測序由生工生物工程(上海)股份有限公司完成。Annealing Buffer for DNA Oligos (5×),購自碧云天生物技術(上海)股份有限公司。Taq酶、dNTP,購自日本TaKaRa公司;T4 DNA ligase,購自加拿大Fermentas公司;AgeI-HF、EcoRI-HF,購自美國NEB公司。兔抗人GATA4單克隆抗體、β-actin一抗,購自美國Proteintech公司;鼠抗兔二抗,購自美國Santa Cruz公司。
1.2 RNAi慢病毒載體構建 將pMGic7.1載體置于38 ℃恒溫水箱,用AgeI-HF、EcoRI-HF酶切。酶切體系:pMGic7.1載體4 μL,AgeI-HF內切酶4 μL,EcoRI-HF 4 μL,Annealing Buffer for DNA Oligos (5×) 4 μL,ddH2O補足至20 μL。經瓊脂糖凝膠電泳回收目標條帶。
利用siRNA軟件(http://design.RNAi.jp/)設計2條目的基因靶序列,序列1:5′-CCGGGACTTCTCAGAAGGCAGAGAGCTCTCTGCCTTCTGAGAAGTC-
TTTTTTG-3′、序列2:5′-AATTCAAAAAAGACTTCTCAGAAGGCAGAGAGCTCTCTGCCTTCTGAGAAGTC-3′,得到siRNA重組載體,分別標記為PLVE2636-1、PLVE2637-1、PLVE2638-1和PLVE2636-2、PLVE2637-2、PLVE2638-2,作為實驗組,然后進行PCR擴增。PCR擴增體系共50 μL:Buffer 10 μL,dNTP 4 μL,上下游引物各1 μL,質粒模板1 μL,PrimeSTAR HS DNA Polymerase 1 μL,ddH2O補足至50 μL;反應條件:98 ℃ 5 min,98 ℃ 10 s、55 ℃ 10 s循環30次,72 ℃ 100 s,72 ℃ 8 min,4 ℃ 10 min。
轉化感受態大腸桿菌DH5α細胞,挑取菌落鑒定。PCR反應體系共20 μL:10×PCR Buffer(with Mg2+) 2 μL,dNTP 1.6 μL,模板1 μL,Taq酶0.1 μL,上游引物(5′-TACGATACAAGGCTGTTAGAGAG-3′)0.8 μL、下游引物(5′-CTATTAATAACTAATGCATGGC-3′)0.8 μL,ddH2O補足至20 μL;反應條件:94 ℃ 5 min,94 ℃ 30 s、55 ℃ 30 s、75 ℃ 50 s、70 ℃ 6 min共循環30次,最后10 ℃ 10 min。將反應產物送生工生物工程(上海)股份有限公司測序。
1.3 慢病毒包裝 將293T細胞接種于含10% FBS、1%青鏈霉素雙抗、1%谷氨酰胺的完全培養基,置于37 ℃、5% CO2的細胞培養箱培養。當細胞生長融合80%左右時,吸棄上清液,加入等體積完全培養基。將混合物移至10 mL離心管,依次加入GATA4載體質粒100 μg、VSVG-GFP質粒35 μg、pCMV-dR8.9包裝質粒65 μg,Trans-EZ溶液500 μL,用完全培養基補足至5 mL。室溫下孵育30 min,用25 mL移液器移至培養板中,8 h后移除上清液,加入等體積完全培養基,置于37 ℃、5% CO2的細胞培養箱內培養。持續培養48 h,收集上清液,高速離心后PVDF膜過濾上清液,濃縮病毒原液,-80 ℃保存。
1.4 病毒滴度測定 按10倍梯度稀釋病毒原液,取3個1.5 mL EP管,每管加入完全培養液90 μL,然后向第1個EP管中加入病毒原液10 μL,充分混勻;吸取第1個EP管中病毒稀釋液10 μL,加入第2個EP管中,充分混勻;吸取第2個EP管中病毒稀釋液10 μL,加入第3個EP管中,充分混勻。通過公式,計算病毒滴度。病毒滴度(IU/mL)=(C×N×D×1 000)/V。其中,C為平均每基因組整合的病毒拷貝數,N為感染時細胞的數目(約1×105個),D為病毒載體的稀釋倍數,V為加入的稀釋病毒體積。
1.5 慢病毒轉染293T細胞 體外常規培養293T細胞。取傳2代、對數生長期、生長狀態良好的293T細胞,消化后計數,制成密度為5×104個/mL細胞懸液。將細胞懸液接種于6孔板,每孔2 mL,隨機分為實驗組(PLVE2636、PLVE2637、PLVE2638)、空白對照組、陰性對照組,每組設3個復孔,常規培養24 h。293T細胞的最佳感染復數(MOI)為10。根據公式:病毒體積=(MOI×細胞數)/病毒滴度,計算需要加入的轉染病毒體積。轉染24 h,加入Polybrene,培養48 h后更換新的培養基,倒置熒光顯微鏡(100×)下觀察熒光標記細胞。
1.6 GATA4蛋白和mRNA表達檢測 ①GATA4蛋白表達檢測:采用Western blotting法。取傳2代、對數生長期、生長狀態良好的293T細胞,經細胞裂解液裂解,提取細胞總蛋白。蛋白定量合格后,加入上樣緩沖液,100 ℃水浴10 min,使蛋白充分變性。取變性蛋白20 μg,SDS-PAGE分離,將蛋白電泳產物轉印至PVDF膜,10%脫脂奶粉室溫封閉6 h,加入兔抗人GATA4單克隆抗體、β-actin一抗,4 ℃孵育過夜。次日加入鼠抗兔二抗,室溫孵育4 h。DAB顯影,Image J軟件分析各蛋白電泳條帶灰度值。以β-actin為內參,以目的蛋白電泳條帶灰度值與內參蛋白電泳條帶灰度值的比值作為目的蛋白相對表達量。②GATA4 mRNA表達檢測:采用RT-qPCR法。取傳2代、對數生長期、生長狀態良好的293T細胞,根據RNAiso Plus總RNA抽提試劑盒說明提取細胞總RNA,經紫外分光光度計鑒定,提取的總RNA濃度和純度合格,可用于后續實驗。按One ShineTMFirst Strand cDNA Synthesis Kit說明,將總RNA逆轉錄為cDNA。逆轉錄條件:37 ℃ 15 min,85 ℃ 5 s。按TaKaRa PCR試劑盒說明進行PCR擴增。引物序列:GATA4上游引物5′-CTCTACATGAAGCTCCAC-3′、下游引物5′-CTGCTGGTGTCTTAGATT-3′,β-actin上游引物5′-CATGTACGTTGCTATCCAGGC-3′、下游引物5′-CTCCTTAATGTCACGCAC-3′。PCR反應體系共10 μL:SYBR Premix Ex Taq Ⅱ 5 μL,上下游引物各0.5 μL,cDNA模板 2 μL,RNase-Free ddH2O 2 μL;反應條件:90 ℃ 40 s,95 ℃ 5 s、65 ℃ 40 s共40個循環,95 ℃ 10 s,65 ℃ 5 s。以β-actin為內參,采用2-ΔΔCT法計算GATA4 mRNA相對表達量。
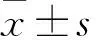
2 結果
2.1 RNAi慢病毒載體鑒定 pMGic7.1載體經AgeI-HF、EcoRI-HF酶切,回收載體大片段,片段長度約8.2 kb。將GATA4干擾片段連接入表達載體,經PCR鑒定,實驗組(PLVE2636、PLVE2637、PLVE2638)轉化單斑長度為260 bp,經生工生物工程(上海)股份有限公司測序,對比siRNA序列,證實pMGic7.1載體與序列1正確連接,其基因序列如下:CGAAACACCGGGGACATAATCACTGCGTAATCC-TCGAGGATTACGCAGTGATTATGTCCTTTTTTG-AATTCGGA(黑體部分為siRNA干擾序列),符合實驗要求。
2.2 慢病毒包裝與病毒滴度測定結果 實驗組(PLVE2636、PLVE2637、PLVE2638)持續培養24 h,顯微鏡下觀察細胞融合接近70%時,可見GFP標記;培養48 h,可見60%以上細胞貼壁生長,細胞內逐漸形成多核體。顯微鏡下觀察病毒原液,明視野下病毒生長良好,結構完整,GFP標記較多,亮度高,呈現“滿天星”現象。顯微鏡下觀察病毒10倍稀釋時,明視野清晰,無雜質,病毒生長良好,結構完整,病毒密度下降,GFP標記較病毒原液減少,亮度降低。顯微鏡下觀察病毒100倍稀釋時,明視野下病毒生長良好,與病毒原液相比,單個病毒形態變化不大,包膜完整,但每視野下GFP標記明顯減少,呈“星空”現象。經測定,病毒拷貝數隨稀釋倍數增加而降低,但病毒滴度變化不大,即病毒效價變化不大。結果見表1。

表1 實驗組病毒滴度測定結果
2.3 慢病毒轉染293T細胞鑒定 維持Polybrene濃度5 μg/mL轉染48 h,GFP標記基本穩定。轉染72 h,顯微鏡下觀察,293T細胞生長狀態良好,無核破裂、核溶解。空白對照組因未添加目的基因片段,無熒光標記;陰性對照組和實驗組熒光標記達峰,但陰性對照組滿視野可見熒光標記,而實驗組熒光標記明顯少于陰性對照組。根據病毒感染率=熒光細胞數/明視野細胞數×100%,實驗組病毒感染率為(81.345±1.173)%。
2.4 各組GATA4蛋白和mRNA表達比較 GATA4蛋白大小約45 kDa,空白對照組未添加目的基因,在45 kDa處無蛋白條帶。陰性對照組和實驗組(PLVE2636、PLVE2637、PLVE2638)均可見目的基因蛋白條帶。實驗組GATA4蛋白相對表達量為0.739±0.056,陰性對照組為0.997±0.001。實驗組GATA4蛋白相對表達量低于陰性對照組(t=6.600,P<0.05)。
實驗組GATA4 mRNA相對表達量為0.362±0.024,陰性對照組為0.998±0.001,空白對照組為0.812±0.001。實驗組GATA4 mRNA相對表達量明顯低于陰性對照組和空白對照組(t分別為36.653、27.401,P均<0.05)。
3 討論
CHD是小兒最常見的出生缺陷性疾病。流行病學調查發現,目前我國新生兒CHD的發病率為2.9‰,較10年前明顯提高,男嬰高于女嬰,主要以VSD為主。CHD的發病機制至今仍不完全清楚,但遺傳因素和環境因素已被證實是其主要病因。
有研究證實,GATA4是人類CHD的致病基因之一。GATA4屬于GATA家族,其基因定位于人染色體8p22-23,cDNA全長3 371 bp,C-末端和N-末端的鋅指結構能夠與DNA穩定轉錄因子連接[8],從而在心肌發育和成熟過程中發揮重要作用[9]。GATA4通過Wnt信號通路控制心臟結構形成,正向增強心肌蛋白活性,協同HDAC2參與心肌細胞增殖,同時亦參與心肌細胞凋亡[10-11]。有研究對比41例動脈導管未閉(PDA)患者與342例健康志愿者DNA序列,發現PDA患者存在GATA4基因rs372504711位點突變,經關聯分析得出GATA4基因TT、CT基因型者PDA患病風險明顯高于CC基因型者[12]。有研究發現,GATA4基因rs4841587位點突變與CHD有一定相關性,rs4841587位點T/G多態性與VSD的易感性和病情程度密切相關[13]。有研究報道,在GATA4序列中家族性ASD患者存在非同義位點變異,如c.839 C>T(T280M),該位點雖然在CHD患者普遍存在,但在健康兒童中不存在此位點突變[14]。GATA4可在體內持續表達,但在不同階段對心肌的作用具有特異性。在大鼠心肌梗死模型中,提高GATA4表達可有效降低心肌纖維化,減少瘢痕組織代償性。在心肌炎癥病變期,炎癥因子可誘導GATA4高表達,從而促進心肌重塑、心肌纖維化,增加氧化應激和心臟重構易感性并參與房顫發生[15-16]。目前關于GATA4與CHD的關系僅涉及流行病學、SNP位點等方面,對其具體機制尚不明確。
本研究通過RNAi技術構建GATA4慢病毒載體并轉染293T細胞,多途徑觀察了GATA4在體外的表達水平。RNAi通過同源雙鏈RNA誘導沉默特定序列,從而影響其功能性表達,因其具有快速、高效、精準等特點,已被廣泛用于分子基礎研究。運用siRNA在線軟件設計2條目的基因靶序列,連接載體后送生工生物工程(上海)股份有限公司測序,證實pMGic7.1載體與第1條siRNA序列正確連接,其原因可能在于設計siRNA時避開了胞嘧啶和鳥嘌呤富集區,避免了重復序列,防止甲基化對堿基互補配對的干擾[17]。慢病毒載體被認為是進行RNAi的最佳搭檔,與一般載體相比,目的基因在慢病毒中穩定存活且慢病毒可轉染處于各生長周期的載體細胞,而被轉染的載體細胞不會因慢病毒的存在而影響傳代和增殖,在反轉錄體系中較為穩定[18]。慢病毒轉染293T細胞后,熒光顯微鏡下持續觀察發現,維持Polybrene濃度5 μg/mL轉染48 h,陰性對照組和實驗組GFP標記逐漸增多;轉染72 h,慢病毒載體中熒光標記細胞比例達到高峰,在此過程中空白對照組持續無GFP標記,而陰性對照組GFP標記最多。GATA4蛋白大小約45 kDa。本研究空白對照組在45 kDa處無蛋白條帶,陰性對照組因未添加siRNA片段,GATA4蛋白未受抑制,蛋白條帶明顯,而實驗組蛋白條帶亮度明顯低于陰性對照組。進一步分析蛋白條帶灰度值發現,實驗組GATA4蛋白相對表達量低于陰性對照組;通過RT-qPCR技術分析發現,實驗組GATA4 mRNA相對表達量明顯低于陰性對照組和空白對照組。結果證實,目的基因已被抑制。
綜上所述,本研究成功構建了靶向GATA4基因的慢病毒載體,并進一步證實CHD的致病基因GATA4在體外可被有效抑制。
