水稻矮桿小粒突變體dsg1 的表型鑒定及粒形基因精細定位
2020-12-09 07:00:32呂召坤玉一嵐李蘭英張德春
農業現代化研究 2020年6期
關鍵詞:水稻
呂召坤 ,玉一嵐 ,李蘭英,張德春
(1.中國科學院亞熱帶農業生態研究所亞熱帶農業生態過程重點實驗室,湖南 長沙 410125;2.中國科學院大學,北京 100049;3.三峽大學生物技術研究中心,湖北 宜昌 443002)
稻米的產量和品質對于確保國民糧食安全和促進農業可持續發展具有重要意義。水稻(Oryza sativa)是世界主要糧食作物之一,全世界有接近一半的人口以稻米為主食。雖然水稻半矮桿基因的發掘和應用以及雜種優勢的利用使得水稻產量取得了巨大的飛躍[1],然而,面對世界人口的持續增長(預計2050 年將達到90 億)[2],水稻育種家和植物遺傳學家需要進一步提高水稻的產量。
水稻產量是一種多性狀共同決定的復雜農藝性狀,主要受有效穗數、每穗粒數和千粒重三因素制約[3]。有效穗數在很大程度上取決于單株分枝數、分蘗角度和株高;每穗粒數主要取決于穗部結構,如穗長、每穗分枝數、籽粒密度和育性;而千粒重直接受到籽粒的形狀和大小的影響[4]。此外,粒形也是評價稻米品質的重要指標之一,印度、越南和美國偏愛細長型稻米,而中國東北、韓國和日本更喜歡短圓型稻米[5]。因此,剖析稻米粒形的遺傳基礎對水稻產量和品質具有重要意義。
粒形可以劃分為粒長、粒寬、粒厚三個不同的影響因子,且三者都是由多基因控制的數量性狀[6]。揭示水稻粒長、粒寬和粒厚的分子遺傳機理,對于全面解析控制粒形性狀的分子調控網絡和挖掘可利用的優異育種基因資源具有重要意義。因此,尋找并克隆控制相應性狀的數量性狀位點(Quantitative trait loci, QTL)及剖析相關信號轉導網絡是研究粒形的重要途徑。目前,在水稻12 條染色體上檢測到與粒形相關的QTL 已經超過600 個,其中93 個已被成功克隆并進行了功能分析[7-8]。它們通過不同的信號途徑調控水稻粒形的發育,主要包括泛素—蛋白酶體途徑、G 蛋白信號途徑、轉錄調控途徑、MAPK 信號途徑以及植物激素調控途徑。GW2 和GW5 通過泛素—蛋白酶體途徑調控水稻籽粒大小。GW2編碼一個E3 泛素連接酶,通過降解促進細胞分裂的蛋白酶負調控水稻穎殼大小,進而影響籽粒大小[9]。GW5編碼的蛋白質與多聚泛素有相互作用,通過泛素—蛋白酶體途徑調控籽粒穎殼的細胞分裂數目對籽粒大小進行調控[10-11]。GS3和DEP1通過G 蛋白信號途徑參與稻米粒形的調控。GS3編碼一個由5 個外顯子組成的Gγ 亞基,它的功能缺失會導致籽粒明顯變小;而DEP1是一種功能獲得性突變基因,功能獲得性突變dep1能夠促進細胞分裂,并使穗粒數增加,促進水稻增產[12-14]。GLW7、GW8、LGY3等在轉錄調控因子途徑中參與水稻粒形的調控,GW8和GLW7的編碼蛋白都是調控粒形的正向調控因子,其高表達水平能夠促進穎殼細胞分裂,導致籽粒變大,LGY3編碼的轉錄因子在第7 內含子和第8 外顯子的連接區發生突變,產生一個新的可變剪切位點,導致籽粒變長[15-17]。SMG1編碼一個絲裂原活化蛋白激酶MAPK4,通過MAPK 信號途徑調控籽粒形狀;而GS2/GL2參與了植物激素油菜素內酯調控途徑[18-21]。雖然這些基因部分解析了調控稻米粒形的分子機理,但這對于了解水稻粒形遺傳網絡還遠遠不夠。因此,發現新的調控粒形的QTL 位點及其調控機制更有利于全面闡述整個遺傳網絡,為今后水稻產量和稻米品質的提高提供遺傳基礎。
在本研究中,矮桿小粒突變體dsg1(dwarf and short grain)是由TB309 經甲基磺酸乙酯(thymethylsulfone,EMS,體積比為0.8%)溶液誘變后篩選得到的。通過對dsg1進行表型和農藝性狀考察,發現粒寬、粒厚和千粒重顯著降低,并且穗長和地上部節間距也顯著縮短。對候選基因進行精細定位,將突變基因dsg1定位于水稻第4 號染色體,位于分子標記ID2798 與ID2803 之間的52.28 kb 區間內,并利用Gramene 數據庫在該定位區間找到11 個候選基因,為dsg1基因的克隆和功能研究奠定了基礎。
1 材料與方法
1.1 試驗材料
dsg1突變體由TB309 經EMS 誘變獲得,經連續多年自交已穩定遺傳。dsg1與9311 雜交后構建F2分離群體進行基因定位。
1.2 試驗方法
1.2.1 主要農藝性狀考察 將實驗材料TB309、突變體dsg1、9311 和F2群體種植于湖南寧鄉關山和海南三亞水稻田,單株種植,行距為26 cm,株距為16 cm,大田常規管理。在水稻成熟期進行株高測量,分別測100 株;并測定莖稈的節間長度,測10 個重復。種子成熟后,單株收取各家系植株,用恒溫干燥箱60 ℃烘烤直至籽粒重量恒重為止。利用游標卡尺對實驗材料TB309、突變體dsg1、9311 和F2群體的成熟種子進行粒長、粒寬和粒厚等指標的測定,每個性狀測10 個重復。將TB309 和dsg1植株的整個穗子取回,測量其穗長、千粒重、結實率等性狀,每個性狀測10 個重復[22]。
1.2.2 遺傳分析 以秈稻9311 和突變體dsg1為親本,經過雜交后自交得到由68 個單株組成的F2群體,對F2群體的粒形分離情況進行統計分析,并進行卡方測驗。
1.2.3 基因定位 根據實驗室已有的存在于水稻12條染色體上的Indel 引物,篩選在dsg1和9311 兩親本間存在多態性的引物。從dsg1/9311 的F2分離群體挑選10 株極端矮化小粒的單株和10 株極端長粒表型的單株,采用極端個體法進行連鎖分析。利用均勻分布于12 條染色體的171 對有多態性的分子標記擴增極端個體的DNA,用3%的瓊脂凝膠進行檢測,并做好數據記錄[23]。使用作圖軟件MAPMAKER(EXP3.0b)構建突變基因所在范圍的分子標記連鎖圖譜,將突變基因定位在一條染色體上的兩個標記之間,完成初定位。初定位后,擴大F2群體,同時使用16 個具有多態性的標記進行候選基因的精細定位,并繪制突變基因所在位置的遺傳圖譜。精細定位所用引物序列見表1。
1.2.4 BR 激素敏感性實驗 將去殼的飽滿水稻種子經75%乙醇和5%次氯酸鈉消毒處理后,在無菌條件下分別播種至含有表油菜素內酯(EpiBL)和不含EpiBL 的1/2 MS 固體培養基,生長8 d 后,觀察根長生長情況,記錄數據[24]。
1.2.5 數據分析 所有數據的平均值計算、顯著性差異分析、卡平方測驗均用Microsoft Excel 軟件進行。根據x2的數值和自由度得到概率值,以P=0.05 為適合性測驗符合的下限。
2 結果與分析
2.1 突變體dsg1 的表型分析
在自然生長條件下,對dsg1與TB309 進行表型考察。利用t測驗對表型數據進行分析,分析結果顯示:與野生型相比,突變體dsg1表現出明顯的矮桿表型(表2,圖1a),其株高約為TB309 的79.00%,倒一節、倒二節、倒三節和穗長的長度顯著低于TB309(表2,圖1b、圖1d)。與TB309 相比,dsg1的籽粒出現明顯的小粒表型,粒長、粒寬、粒厚分別減小15.53%、7.26%和10.45%,千粒重降低了34.14%(表2,圖1c)。另外,與野生型相比,dsg1分蘗數也顯著減少,單株總產量顯著降低。

表1 本研究使用的引物Table 1 The primers used for this study

表2 野生型TB309 與突變體dsg1 的表型鑒定Table 2 Comparison of agronomic trait of the wild type TB309 and the dsg1 plants
2.2 油菜素內酯激素敏感性分析
由于突變體不僅在粒形性狀上發生改變,在植株高度、分蘗數目、節間長度和稻穗長度等方面也表現出顯著性變化,這與先前報道的油菜素內酯的合成或代謝缺陷表型相似[25-26],因此,我們猜測dsg1突變體可能與BR 激素有關。為了驗證這個猜測,我們對野生型TB309 和突變體dsg1進行了BR激素響應的敏感性實驗。我們通過外施EpiBL 的方式檢測DSG1是否參與BR 響應。在施加EpiBL 培養后,野生型TB309 的主根長度明顯縮短,說明外源施加EpiBL 抑制了根部的伸長;然而突變體dsg1的主根長度并沒有顯著性變化(圖2a, 圖2b)。由此表明,dsg1并不是一個BR 合成缺陷突變體,而有可能是BR 信號轉導過程中出現缺陷導致了突變。

圖1 野生型TB309 與突變體dsg1 的表型鑒定Fig. 1 Phenotypic identification of the wild type TB309 and the dsg1 mutant
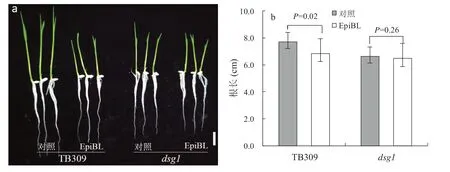
圖2 突變體dsg1 對BR 響應敏感性Fig. 2 Mutant dsg1 as a BR-insensitive mutant
2.3 遺傳分析
為了克隆該突變體的基因,把粒長作為目標性狀對DSG1 進行遺傳效應分析和精細定位研究。我們將dsg1與9311 進行雜交,得到F1代植株,其表型與9311 表型相似。將F1代自交獲得F2群體,在F2群體中出現9311 粒長表型和突變體小粒表型分離,說明該突變表型是由隱性基因控制的。統計F2群體中粒長表型的單株數量,發現粒長表型頻率呈現雙峰分布(圖3),表明存在主效QTL。在F2群體中,長粒和短粒表型數目分別為42 和26,。對粒長表型進行x2測驗,兩種表型在F2群體中的分離比符合3 ∶1(x2=0.16,x20.05=3.84,df=1),符合孟德爾遺傳規律,表明該dsg1的粒長性狀受單基因控制。
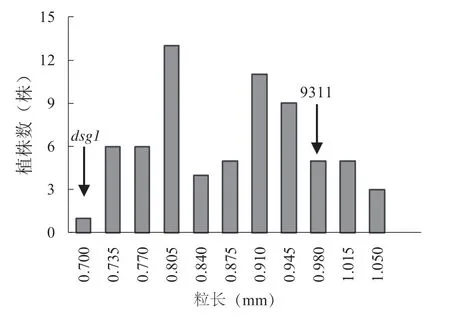
圖3 F2 群體粒長性狀頻率分布Fig. 3 Frequency distribution of grain length in a F2 population
2.4 基因定位
將dsg1與9311 進行雜交,在F2分離群體中分別選取10 株極端小粒表型單株和極端長粒表型的單株,利用均勻分布于12 條染色體的Indel 分子標記進行連鎖分析,將DSG1基因初定位于第4 染色體標記ID2610 和ID3004 之間。為了進一步對候選QTL 進行精細定位,在初定位區間內設計14 對新的多態性分子標記,利用交換單株縮小定位區間,最終將突變基因定位在分子標記ID2798 與ID2803的區間內,物理距離約為52.28 kb,其間包含11 個候選基因(圖4)。
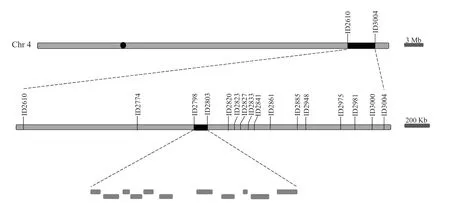
圖4 水稻突變體dsg1 的基因定位Fig. 4 Fine mapping of rice mutant dsg1 gene
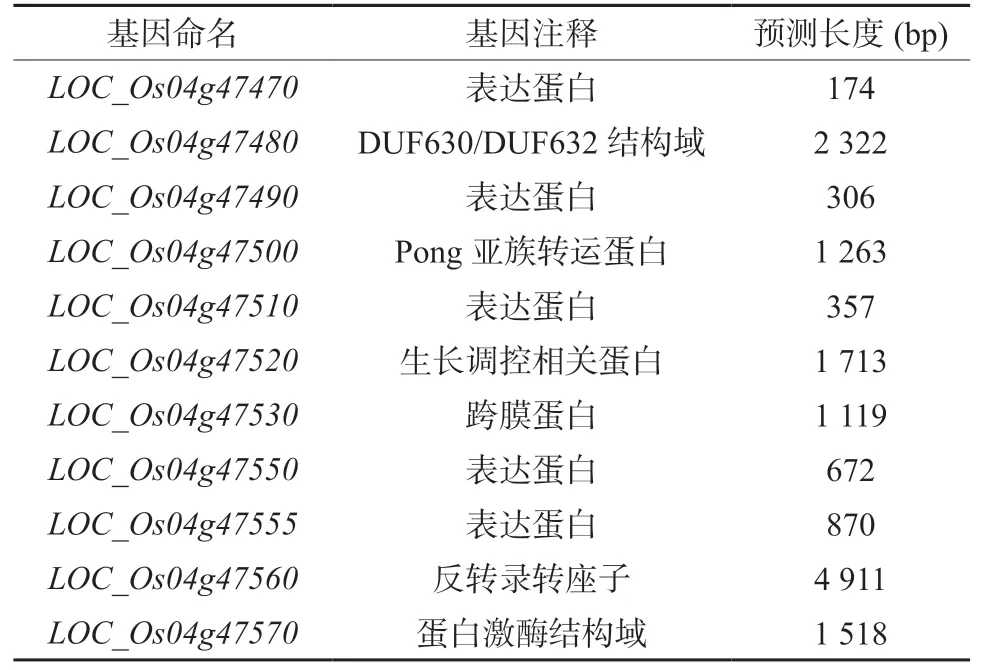
表3 定位區間內的候選基因注釋Table 3 Annotation of DSG1 candidate genes in the location interval
2.5 候選基因的預測及分析
根據水稻基因組數據庫(http://rice.plantbiology.msu.edu)提供的基因注釋信息,在52.28 kb 候選區間內存在11 個注釋基因(表3)。其中3 個編碼表達蛋白,2 個功能未知蛋白,其余分別編碼轉座蛋白、逆轉錄轉座子蛋白、跨膜蛋白、含有DUF630/DUF632 域的蛋白、含有激酶結構域的蛋白和生長調控相關蛋白。通過Gramene 網站進行生物信息學分析后發現,該區間內存在3 個可能與水稻株高、穗型和粒性發育相關的基因。LOC_Os04g47480預測結果顯示,其編碼蛋白可能含有DUF630/DUF632 結構域。未知功能結構域蛋白家族(domains of unknown function protein families, DUFs)是一類進化保守且有功能的蛋白家族,其不同成員參與不同的生長發育過程及脅迫響應[27]。含有DUF630/DUF632 結構域的蛋白參與了葉片形態建成,進而導致了稻米籽粒大小的變化。LOC_Os04g47570的表達蛋白含有激酶結構域,其同源基因LOC_Os03g40400 的表達產物參與了GS3 和qGL3 調控稻米籽粒大小的信號轉導過程,其表達水平上調了1.5 倍,該基因的表達可能會影響稻米籽粒的大小[28]。LOC_Os04g47510 在水稻中有一個同源基因LOC_Os07g27670,該基因的編碼蛋白是WRKY 家族中的一員。WRKY 家族蛋白是一種轉錄因子,廣泛參與并調控植物生長發育、形態建成和各種生物或非生物脅迫反應[29]。WRKY 蛋白能夠與生長素(IAA)、赤霉素(GA)、脫落酸(ABA)以及油菜素內酯(BR)等多種植物激素相互作用調控植物生長發育[30-31]。在擬南芥中,WRKY46、WRKY54 和WRKY70 能促進與BR 相關的生長基因表達并抑制抗旱基因的表達,調節擬南芥生長[32]。在蘋果中,轉錄因子MdWRKY9 直接抑制油菜素類固醇限制合成酶MdDWF4 的轉錄,減少BR 產生,調控植株高度[33]。在水稻中,OsWRKY53 能夠正向調控BR信號進而調控水稻葉片角度及籽粒大小,其過表達植株的葉角增加,稻米籽粒變大;而OsWRKY78的RNAi 植株可能由于細胞長度的縮減而表現出半矮化和小籽粒的表型[34-35]。因此,這3 個基因都有可能是DSG1的候選基因。
3 討論
水稻粒形是重要的農藝性狀之一,對水稻的產量和品質起到重要作用,是多基因、多途徑調控的復雜遺傳性狀,也是分子設計育種的主要性狀之一[36]。分子設計育種是在解析作物重要農藝性狀遺傳機理的基礎上,通過品種設計對復雜性狀進行定向改良,達到聚合優異性狀的目標,實現作物復雜性狀的精準改良[37]。實現精準設計育種的難點在于調控目標性狀的遺傳網絡非常復雜,所以,分子設計育種的前提和要求是對目標性狀的分子機理和遺傳基礎有較為深刻的理解。雖然在水稻中已克隆93個與粒型相關的QTL,然而,可應用于分子設計育種的控制稻米籽粒的基因非常有限。利用Ghd7不同等位基因之間地理分布不同的特點,Ghd7在不同生態區的水稻生育期改良中得到了高效利用,獲得了生育期改良的新品系[38]。Wang 等將GW8和GW7導入我國高產水稻品種,在不減產的基礎上提升稻米品質[39]。還有研究表明,將GS3、DPE1、qSW5、SD1、Ghd7、Ghd8和Wx等基因聚合獲得的改良品種與超級雜交稻“兩優培九”相比,該新品種表現出更高的產量潛力和更好的籽粒品質[40]。所以,隨著分子設計育種構想的發展,需要挖掘更多的粒形相關基因。在本研究中,我們利用EMS 誘變粳稻品種TB309,篩選得到遺傳性狀穩定的矮桿小粒突變體dsg1,并將其定位于第4 染色體分子標記ID2798 與ID2803 之間52.28 kb 的區間內,以期克隆到相關基因,以便應用于分子設計育種。
此外,我們發現dsg1突變體不僅粒形性狀發生顯著性變化,而且株高、穗長、分蘗數目和節間長度等其他農藝性狀也呈現顯著性減小,說明該基因能夠控制多種不同的表型,這可能與“一因多效”有關。一因多效是自然界中廣泛存在的現象,其概念最早是由孟德爾所提出的,用來描述一個單獨的基因或位點影響兩個或兩個以上不同性狀的現象[41]。例如,GS3編碼的Gγ 亞基,可通過G 蛋白信號途徑調控稻米粒長和粒重,同時對粒寬和粒厚也具有微效作用[15];DEP1不僅能夠調控粒長和千粒重,還對穗長、每穗實粒數、耐旱性等其他性狀有影響[42];WTG1同時控制每穗粒數和分蘗數目2 種不同的性狀[43-44];Ghd7同時調控水稻抽穗期、每穗粒數和株高3 種性狀,猜測可能是抽穗期延遲增加了穗部和秸稈等器官的發育進程從而使產量增加和植株變高[39]。同樣,DSG1可能能夠影響不同的生化代謝途徑,形成復雜的調控網絡,進而影響多種農藝性狀。本研究完成了對該基因的精細定位,并通過基因注釋信息分析和激素敏感性實驗得到3個可能是DSG1的候選基因,為進一步克隆該基因并剖析其分子機制奠定了基礎,同時對于利用分子設計育種的方法改善稻米產量和品質也具有重要意義。
4 結論
1)本研究利用EMS 技術誘變粳稻品種TB309獲得了一個矮桿小粒的突變體dwarf and short grain(dsg1)。與野生型相比,突變體dsg1的株高、粒長、粒寬、粒厚和千粒重顯著降低,并且突變體的穗長和節間距也顯著縮短。
2)通過植物激素敏感性實驗證實dsg1有可能是BR 信號轉導過程中出現缺陷導致了突變,而不是BR 合成缺陷突變體。在施加EpiBL 培養后,野生型TB309 的主根長度明顯縮短,但是突變體dsg1的主根長度并沒有顯著性變化。
3)對突變體dsg1與9311 雜交獲得的F2群體進行表型考察和遺傳分析,F2代出現表型分離,且符合孟德爾遺傳規律,說明該粒長突變基因是由隱性單基因控制。
4)通過精細定位,將突變基因定位于水稻第4染色體分子標記ID2798 與ID2803 之間52.28 kb 的區間內,并利用Gramene 數據庫對定位區間進行基因預測,發現該區間存在11 個基因,這為該突變基因的克隆和功能分析奠定了基礎。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
