增溫和線蟲添加對西雙版納熱帶雨林土壤呼吸的短期效應
2020-11-23 06:05:38周清秋李靜雯王思敏吳彬陳輝鄧云吳建平
生態環境學報 2020年9期
周清秋 ,李靜雯,王思敏 ,吳彬 ,陳輝,鄧云,吳建平 *
1.云南大學/云南省植物繁殖適應與進化生態學重點實驗室,云南 昆明 650500;2.云南大學生態與環境學院,云南 昆明 650500;3.北京師范大學生命科學學院,北京 100875;4.中國科學院西雙版納熱帶雨林生態系統研究站,云南 西雙版納 666303
人類活動造成大氣CO2濃度持續增加,由此引發的氣候變化及溫室效應受到社會各界關注(Melillo et al.,2002;IPCC,2013)。土壤呼吸作為陸地生態系統碳循環的關鍵輸出途徑,主要以植物根系、土壤動物和土壤微生物的呼吸為主,釋放出的CO2影響全球碳平衡及氣候變化過程(Davidson et al.,2006;徐小鋒等,2007)。土壤動物可直接影響土壤碳排放,或通過間接影響土壤微生物和植物生理活動調控土壤呼吸,對有機碳礦化起重要作用(王學霞等,2018)。
土壤線蟲和微生物是陸地生態系統生物多樣性的重要組分,在生態系統過程及對全球變化的響應中扮演了重要角色(邵元虎等,2007;陸雅海等,2015)。土壤線蟲可調控土壤碳、氮等生物地球化學循環過程(Van den Hoogen et al.,2019)。食細菌線蟲和食真菌線蟲在捕食過程中會釋放出CO2、NH4+及其它含氮化合物,對土壤的C、N礦化造成直接影響(邵元虎等,2007)。通過室內微宇宙實驗發現,接種線蟲后土壤CO2排放量有顯著提高,且線蟲對微生物生物量有明顯的調控作用(Fu et al.,2005)。但是,增溫背景下土壤線蟲對微生物的調控及對土壤碳排放的影響研究還鮮有報道。
溫度是影響陸地生態系統物質循環的關鍵環境因素,溫度改變引起的諸多重要生理過程(如土壤生物生存動態及土壤養分循環)的變化,是全球氣候變化的主要表現形式(Melillo et al.,2002)。溫度升高加重土壤水分流失、改變土壤酶活性及熱應激效應,使土壤線蟲群落結構發生變化(Briones et al.,2009)。此外,增溫通過調控植物根系碳輸入和凋落物分解等途徑間接改變土壤生物多樣性,進而影響陸地生態系統功能和全球氣候(Cowles et al.,2016;Daniel et al.,2017)。
熱帶森林土壤生態系統的結構和功能對氣候變化較為敏感,研究全球變暖背景下熱帶地區土壤生物群落動態及其對土壤碳排放的影響具有重要意義(Malhi et al.,2004)。目前,增溫對土壤生態系統影響的研究多集中在溫帶和寒帶地區,在熱帶森林開展研究相對較少(Cavaleri et al.,2015)。因此,本實驗通過研究增溫和線蟲添加條件下熱帶森林土壤呼吸速率和土壤微生物碳利用效率動態,探討土壤呼吸對全球變暖和土壤生物互作的響應及其生物調控機制。
1 材料與方法
1.1 土壤樣品采集
土壤樣品采集于云南省西雙版納熱帶植物園(101°25′E,21°41′N)。該區海拔 570 m,年均溫21.5 ℃,年均降水量約 1400 mm,屬熱帶雨林氣候,土壤類型為磚紅壤。2018年8月,在20 m×20 m的研究樣地內,每隔5 m設置一個取樣點,每個取樣點用直徑5 cm土鉆隨機取5鉆土,取土深度為0—10 cm。將5鉆土混合成一個土壤樣品,共采集10個混合土壤樣品。土樣帶回實驗室后,去除可見根系和石礫,過2 mm篩并測定土壤理化性質(見表1)。有研究表明,熱帶雨林中土壤微生物活動的最佳含水量為飽和含水量的 70%(Clarisse et al.,2019)。因此,本實驗通過稱質量和添加去離子水來維持此適宜含水量。

表1 供試土壤理化性狀Table 1 Physico-chemical characteristics of Xishuangbanna tropical rainforest soil
1.2 實驗設計
將過篩后的10個樣品均勻混合,稱取1000 g混合土置于容器中,攤成2—3 cm薄層,放入-26 ℃冰箱冰凍3 d,再轉入生化培養箱于22 ℃下培養7 d,按此操作反復凍融5—7次。期間用淺盤法進行分離檢驗,直至去除土壤中所有活體線蟲為止(毛小芳等,2004;周際海等,2017)。
本實驗中添加的線蟲為秀麗隱桿線蟲(Caenorhabditiselegans),由云南省生物資源開發與利用國家重點實驗室提供。此類線蟲取食細菌,常被作為模式生物開展研究(周際海等,2017)。實驗開始前,用0.1%硫酸鏈霉素和0.002%放線菌酮的混合消毒液對線蟲進行殺菌,防止其表面微生物對實驗產生干擾(李輝信等,2002)。準備30個500 mL的無菌玻璃瓶,每瓶稱取30 g凍融土。根據該地區土壤線蟲豐度,往線蟲添加處理的培養瓶中加入20條殺菌后的線蟲(Xiao et al.,2014)。在所有培養瓶蓋子上開一個小口,接上 PVC管且連接三通氣閥,用熱熔膠密封以保證培養過程的密閉性,采氣時打開三通氣閥。
依據西雙版納年均溫和IPCC報告預測的增溫范圍(1.1—6.4 ℃)(IPCC,2013),實驗設置3個增溫梯度,共計 30個獨立培養瓶。線蟲添加完畢后,關閉所有三通氣閥,并將培養瓶分別放入21.5、24.5、27.5 ℃的恒溫培養箱中。參照土壤生物通用培養研究方法(Treat et al.,2014),本實驗將培養時間設置為30 d。具體實驗處理為:
(1)處理一T21.5:21.5 ℃培養(CK)
(2)處理二T24.5:24.5 ℃培養
(3)處理三T27.5:27.5 ℃培養
(4)處理四 NeT21.5:21.5 ℃培養+線蟲添加(20 條)
(5)處理五 NeT24.5:24.5 ℃培養+線蟲添加(20 條)
(6)處理六 NeT27.5:27.5 ℃培養+線蟲添加(20 條)
1.3 土壤呼吸和土壤微生物生物量碳測定
基于頂空CO2濃度隨時間的變化計算土壤呼吸速率,培養和測量方法參照Chen et al.(2016)。初次測量之前,打開玻璃瓶瓶蓋并放入通風櫥對流 1 h。待瓶子中的氣體濃度分布均勻后,用注射器抽取5 mL頂空氣體在氣相色譜儀上測定CO2含量,此為CO2初始濃度。為避免負壓造成的潛在影響,抽取頂空氣體后立即將5 mL無CO2空氣注入瓶中。然后將所有玻璃瓶置于各自的設定溫度下,48 h后,再次抽取5 mL頂空氣體,以測算期間的CO2濃度變化值,此過程共重復3次。從實驗的第7天起(即第3次抽氣結束后),每隔5—8天測定1次CO2含量,即在實驗開始的當天以及培養的第 2、4、6、11、16、23、30天分別對每個玻璃瓶進行抽氣,依次記錄為 D0、D2、D4、D6、D11、D16、D23、D30。最后1次抽氣完成后,用氯仿熏蒸法測定土壤微生物生物量碳(Yao et al.,2000)。
1.4 統計分析
參照Wu et al.(2014)計算土壤CO2呼吸速率(Rt,μg·g-1·d-1),公式如下:
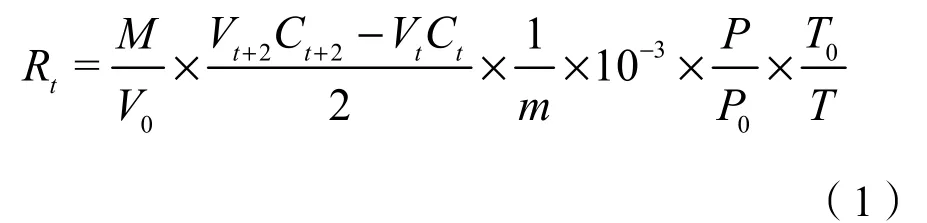
式中:M:CO2摩爾質量,44 g·mol-1;V0:氣體體積常數,22.4 L·mol-1;Vt+2:測量當天培養瓶內氣體體積(mL);Ct+2:測量當天培養瓶內CO2濃度(μg·g-1);Vt:培養瓶內氣體初始體積(mL);Ct:培養瓶內 CO2初始濃度(μg·g-1);m:干土質量(g);P0:標準大氣壓,1013.25 hPa;P:測量時培養瓶內實際氣壓(hPa);T0:標準狀態熱力學溫度,273.15 K;T:測量時實際培養溫度(K)。
土壤呼吸累積量(Rc)的計算公式參照(王春新等,2017):

式中:Rc為測定時間內的土壤呼吸累積量(μg·g-1);Rt為測量時間內的土壤呼吸速率(μg·g-1·d-1);t為測量時間(d)。
土壤呼吸速率與溫度間呈指數函數關系,通常用土壤呼吸的溫度敏感性(Q10)來表示溫度每升高10 ℃土壤呼吸速率增加的倍數。Q10的計算公式(毛小芳等,2004)為:

式中:Rt為土壤呼吸速率;θ為土壤溫度;a、b為實際參數。
土壤微生物生物量碳(MBC)的計算公式為:

式中:ω(C)為微生物生物量碳(mg·kg-1);EC 為已熏蒸土樣的可溶性有機碳含量與未熏蒸土樣的可溶性有機碳含量差值(mg·kg-1);KEC為微生物體中碳被浸提出來的比例,一般定為0.38(陳國潮,2002)。
微生物碳利用效率(CUE)是衡量微生物群落新陳代謝能力的一個重要參數,其數值大小反映了微生物釋放CO2的強弱程度(Spohn et al.,2016)。CUE的計算公式參考(Maynard et al.,2017):

式中:B為土壤呼吸測定結束后的微生物生物量碳含量(μg);R為土壤呼吸測定結束當天的測得的呼吸碳含量(μg)。
用單因素方差分析(One-way ANOVA)比較處理間差異性,處理間的主效應及交互效應用雙因素方差分析(Two-way ANOVA)檢驗,用一元回歸分析(Simple regression analysis)檢驗土壤溫度、MBC和 CUE對土壤呼吸速率的影響,所有分析顯著水平均設為P<0.05。統計分析和數據整合用Microsoft Excel 2016和SPSS 19.0完成,用Sigma Plot 12.5和Excel軟件繪圖。
2 結果與分析
2.1 增溫和線蟲添加對土壤呼吸的影響
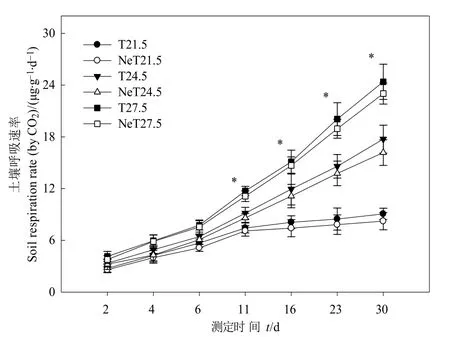
圖1 增溫和線蟲添加對土壤呼吸速率的影響Fig.1 Effects of warming and nematodes addition on soil respiration rate
增溫和線蟲添加均顯著影響土壤呼吸速率(P<0.05,圖1)。在培養期內,6組處理的土壤呼吸速率均隨培養時間呈明顯上升趨勢。從第 11天開始,T21.5、NeT21.5的土壤呼吸速率逐漸趨于平穩,其余處理仍呈明顯上升趨勢;相同溫度下未添加線蟲與添加線蟲處理的土壤呼吸速率差異顯著(P<0.05)。第11天時,T24.5、T27.5分別是T21.5的1.23倍和1.58倍;同一溫度下添加線蟲后土壤呼吸速率比未添加降低4.64%—5.96%。第30天時,T24.5、T27.5分別是T21.5的1.96倍和2.69倍;同一溫度下添加線蟲的土壤呼吸速率比未添加降低 5.59%—9.25%。
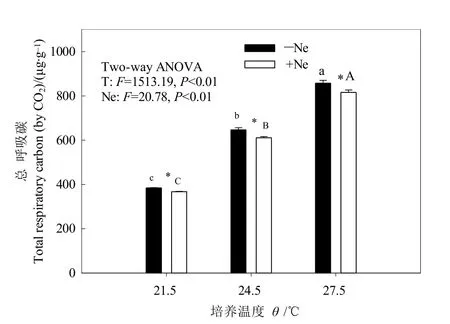
圖2 增溫和線蟲對土壤總呼吸碳的影響Fig.2 Effects of warming and nematode on total soil respiratory carbon
增溫和線蟲添加對土壤總呼吸碳有顯著影響(P<0.05),但二者交互作用不明顯(圖2)。增溫使土壤總呼吸碳顯著升高(P<0.05),T24.5、T27.5分別是T21.5的1.68倍和2.23倍。相同溫度下,添加線蟲后的土壤總呼吸碳比未添加線蟲降低4.44%—5.43%。
2.2 增溫和線蟲添加條件下土壤呼吸的溫度敏感性
用指數函數模型計算不同處理下Q10的變化。結果表明,土壤呼吸速率與土壤溫度顯著正相關(圖3)。線蟲添加條件下Q10顯著提高(P<0.05),較未添加線蟲處理升高5.98%。
2.3 增溫和線蟲添加對MBC和CUE的影響
增溫、線蟲添加、增溫+線蟲添加均對MBC有顯著影響(P<0.05,圖4a)。與T21.5相比,MBC在T24.5下提高8.50%,在T27.5下降低13.88%,線蟲添加后顯著降低15.57%—20.18%。相同溫度下,MBC在線蟲添加后顯著降低7.31%—23.32%。
增溫和線蟲添加對 CUE均有顯著影響(P<0.05,圖4b),但增溫與線蟲添加處理間交互作用不明顯。隨著培養溫度的升高,相同線蟲處理的CUE均顯著下降,未添加線蟲降低 30.49%—55.90%,添加線蟲降低36.16%—52.45%。相同溫度下,添加線蟲處理 CUE比未添加線蟲處理降低2.37%—16.85%。
2.4 MBC和CUE對土壤呼吸的影響
將MBC和CUE與第30天的土壤呼吸速率進行回歸分析(圖5),結果顯示CUE能顯著影響土壤呼吸速率(P<0.05,圖5a),且模型擬合度較高;而MBC與土壤呼吸速率相關性不顯著(圖5b)。
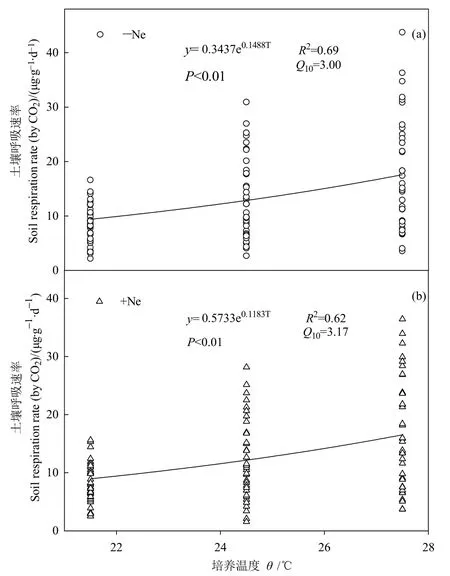
圖3 線蟲添加條件下溫度對土壤呼吸速率的影響Fig.3 Effects of temperature on soil respiration rate under nematodes addition
3 討論
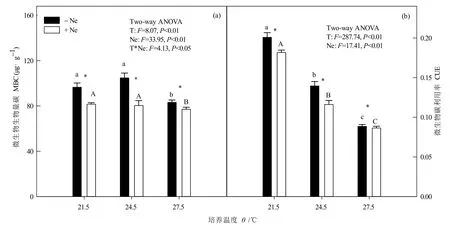
圖4 增溫及線蟲添加對MBC和CUE的影響Fig.4 Effects of warming and nematodes addition on soil MBC and CUE
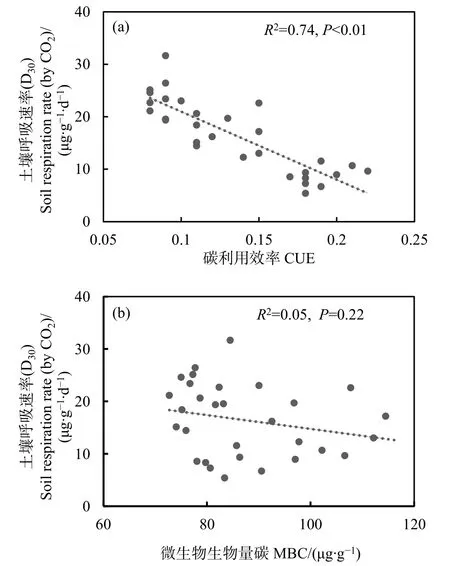
圖5 MBC和CUE對第30天土壤呼吸速率的影響Fig.5 Effects of MBC and CUE on soil respiration rate of the 30th day
本研究中,短期增溫顯著提高土壤呼吸速率,這與增溫可短暫增加土壤呼吸量的研究結果一致(Melillo et al.,2017;康靜等,2019)。溫度作為土壤呼吸速率的主要調控因子,增溫可能直接加速微生物分解作用,促進CO2向大氣中排放(Melillo et al.,2017)。相反,短期增溫降低了CUE。較高的CUE表明土壤中有充足的有效底物供微生物進行自身固碳(Manzoni et al.,2012),而CUE下降則表明微生物同化的碳此時用于呼吸的比例增加,用于自身合成生物量的比例減少,從而促進土壤碳排放。添加線蟲后,土壤呼吸速率明顯下降,CUE下降趨勢則更強,這可能由于微生物受線蟲捕食導致數量減少(圖 4)。土壤微生物以異養呼吸方式釋放出的CO2量,通常能占土壤呼吸總量的一半以上(Bond-Lamberty et al.,2004)。線蟲添加后可能大量取食微生物,導致微生物數量減少,土壤呼吸速率減慢。雖然線蟲對土壤呼吸也具有一定貢獻(Ingham et al.,1985),但不足以彌補因微生物減少而造成的土壤碳排放損失,說明線蟲可能通過調控土壤微生物數量而促進土壤碳固定。
隨著培養溫度的升高,土壤微生物生物量碳呈現先上升后下降的趨勢。當培養溫度從 21.5 ℃升高至24.5 ℃時,微生物生長繁殖受到適度促進,總數量增加,微生物生物量碳含量增加;當溫度繼續升高至 27.5 ℃時,微生物數量卻大幅減少。一方面,微生物對環境溫度變化較為敏感,受增溫刺激后生長勢態和呼吸作用均會顯著增強;另一方面,如果微生物處在未馴化階段,加速變暖則可能使土壤中的有效底物耗盡,繼而對微生物活動產生負反饋效應,微生物生物量則隨之降低。Allison et al.(2010)發現微生物特性可間接影響土壤碳排放,通過降低增溫樣地中土壤微生物生物量來減弱土壤呼吸作用,有效彌補了因增溫引起的土壤碳損失。線蟲添加后,微生物量碳明顯下降,除上述的受到線蟲取食外,微生物生存可能還受到生存空間和資源的限制。線蟲加入加快了有限資源的消耗,為適應環境,微生物群落組成或將變為生長和周轉迅速的r-對策型微生物,而這類微生物生物量較低(周正虎等,2016)。
此外,隨著溫度升高,土壤呼吸作用增強,Q10也呈現出增大的趨勢,這與以往研究結果一致(張彥軍等,2019)。添加線蟲后,土壤呼吸速率有所減緩,但Q10反而增大,造成這一結果的原因可能與線蟲大量繁殖有關,土壤線蟲豐度與溫度有顯著正相關關系(Ruess et al.,1999)。本研究選取的秀麗隱桿線蟲是c-p值為1的食細菌線蟲,此類線蟲世代時間短,產卵量大,在食物富集條件下能快速繁殖(張曉珂等,2013)。線蟲剛添加時,充足的土壤養分使其快速繁殖。此時微生物受線蟲取食而數量減少,MBC含量明顯下降,土壤呼吸速率也隨之下降。由于線蟲本身呼吸作用,所以線蟲添加也貢獻了一部分呼吸碳,因此Q10比無線蟲處理略高。但由于本實驗處理周期較短,受實驗限制未能實時監測培養過程中線蟲數量變化,且秀麗隱桿線蟲的取食偏好在本實驗中也未能明確。因此,未能定量分析線蟲對土壤呼吸的貢獻率,后續研究將從培養過程中線蟲的數量動態和微生物群落結構進行補充驗證。
4 結論
通過室內的短期增溫和線蟲添加實驗,表明增溫顯著增強熱帶雨林土壤的呼吸作用。線蟲添加后土壤呼吸速率減小,土壤呼吸的溫度敏感性發生變化,且土壤微生物生物量碳隨之減少,從而導致微生物碳利用效率下降,影響土壤碳吸存效應,最終可能調控生態系統對全球變暖的響應。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
當代陜西(2021年2期)2021-03-29 07:41:24
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
媽媽寶寶(2017年3期)2017-02-21 01:22:28
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
